
Diversity Patterns and Assemblage Structure of Non-Biting Midges (Diptera: Chironomidae) in Urban Waterbodies

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
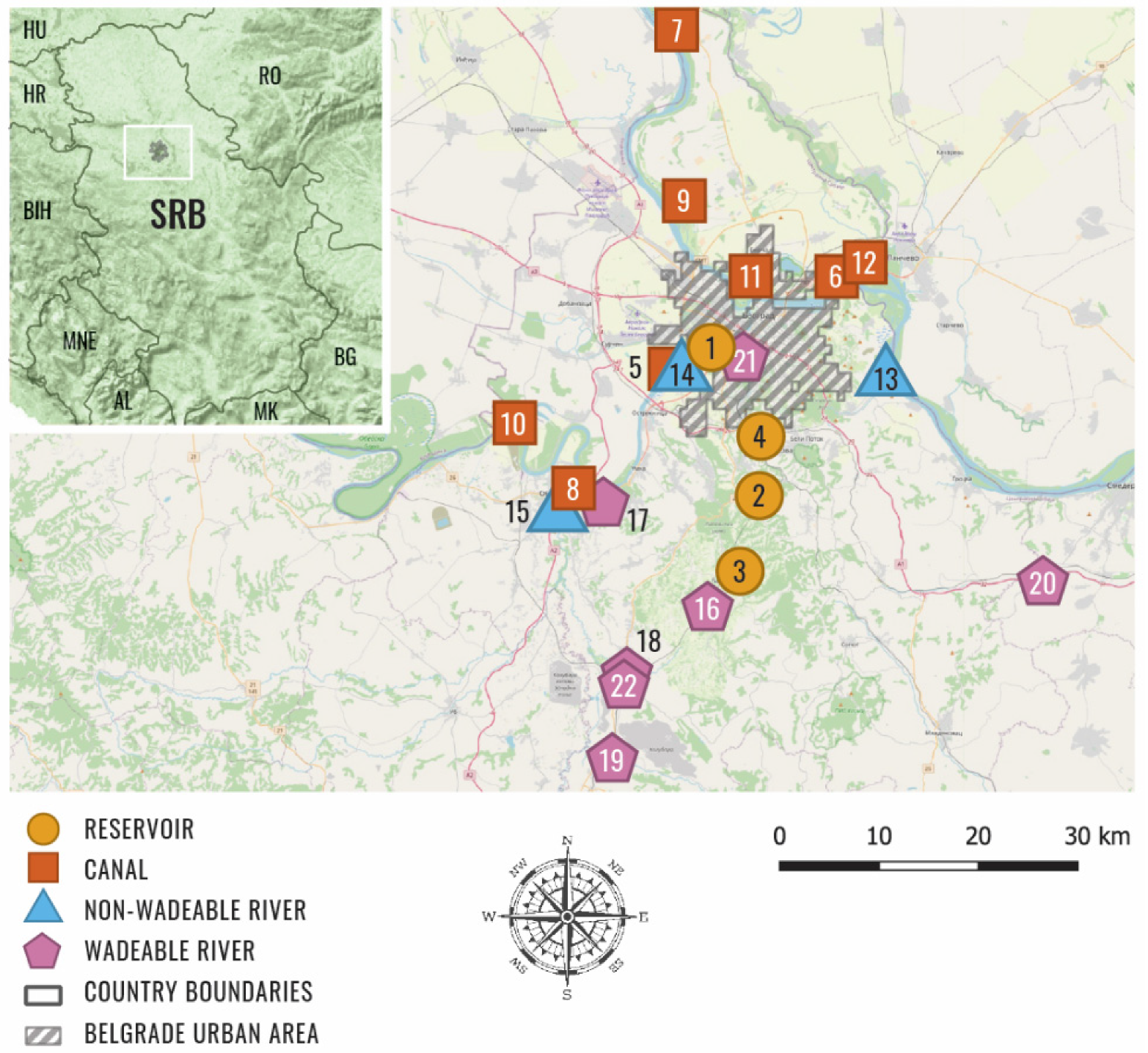
2.1. Study Area
2.2. Chironomidae
2.3. Environmental Variables
2.4. Data Analysis
3. Results
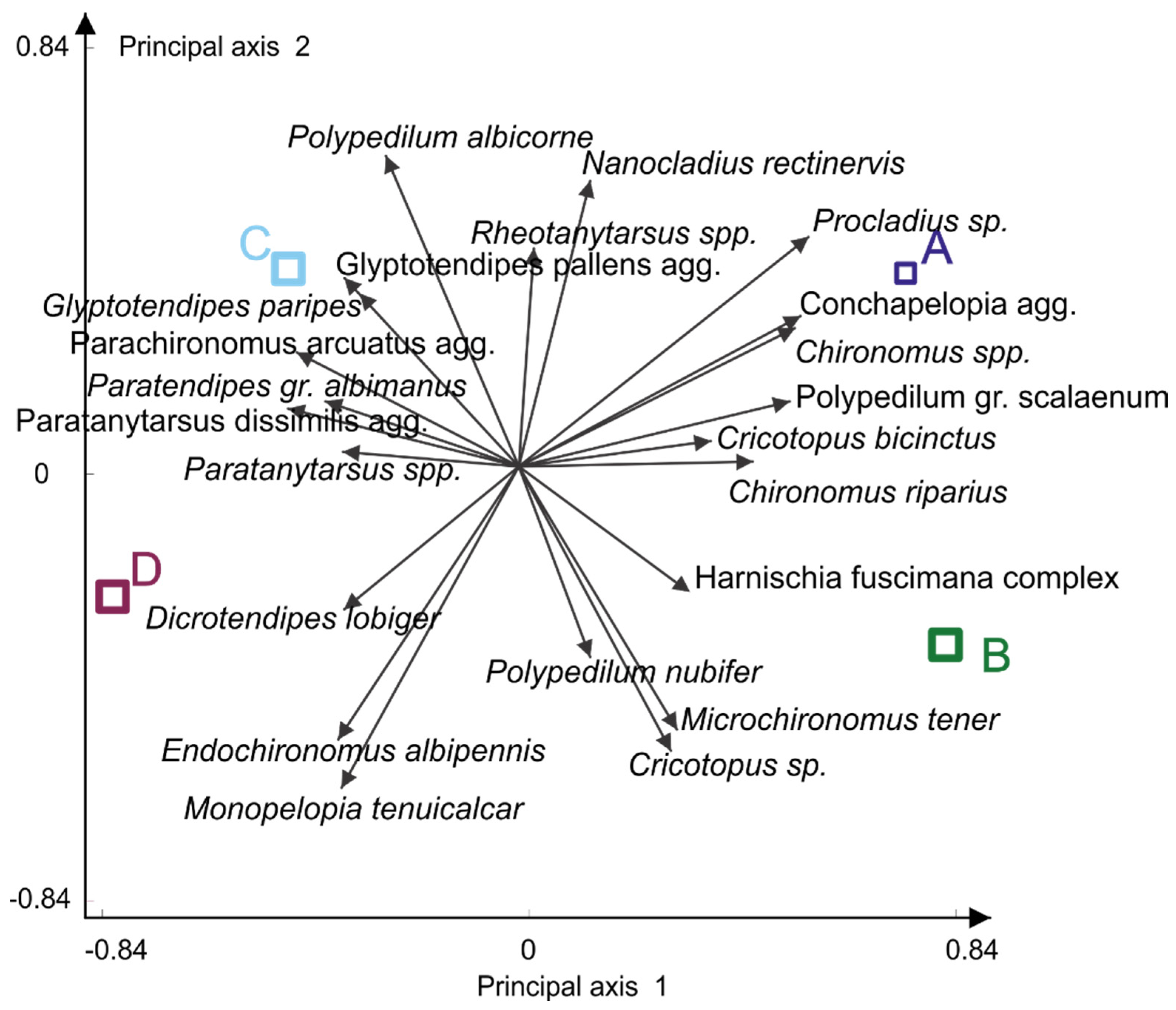
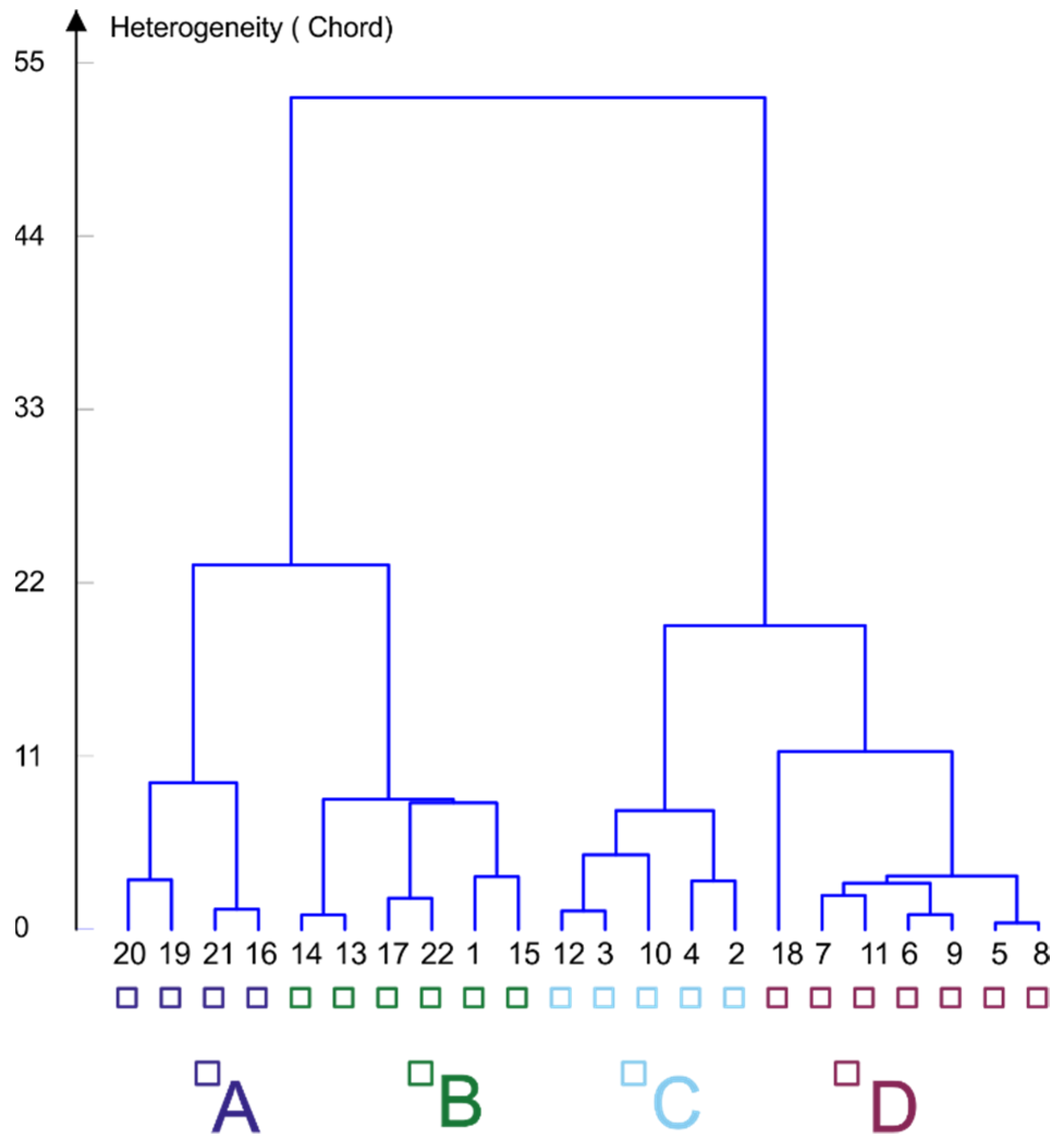
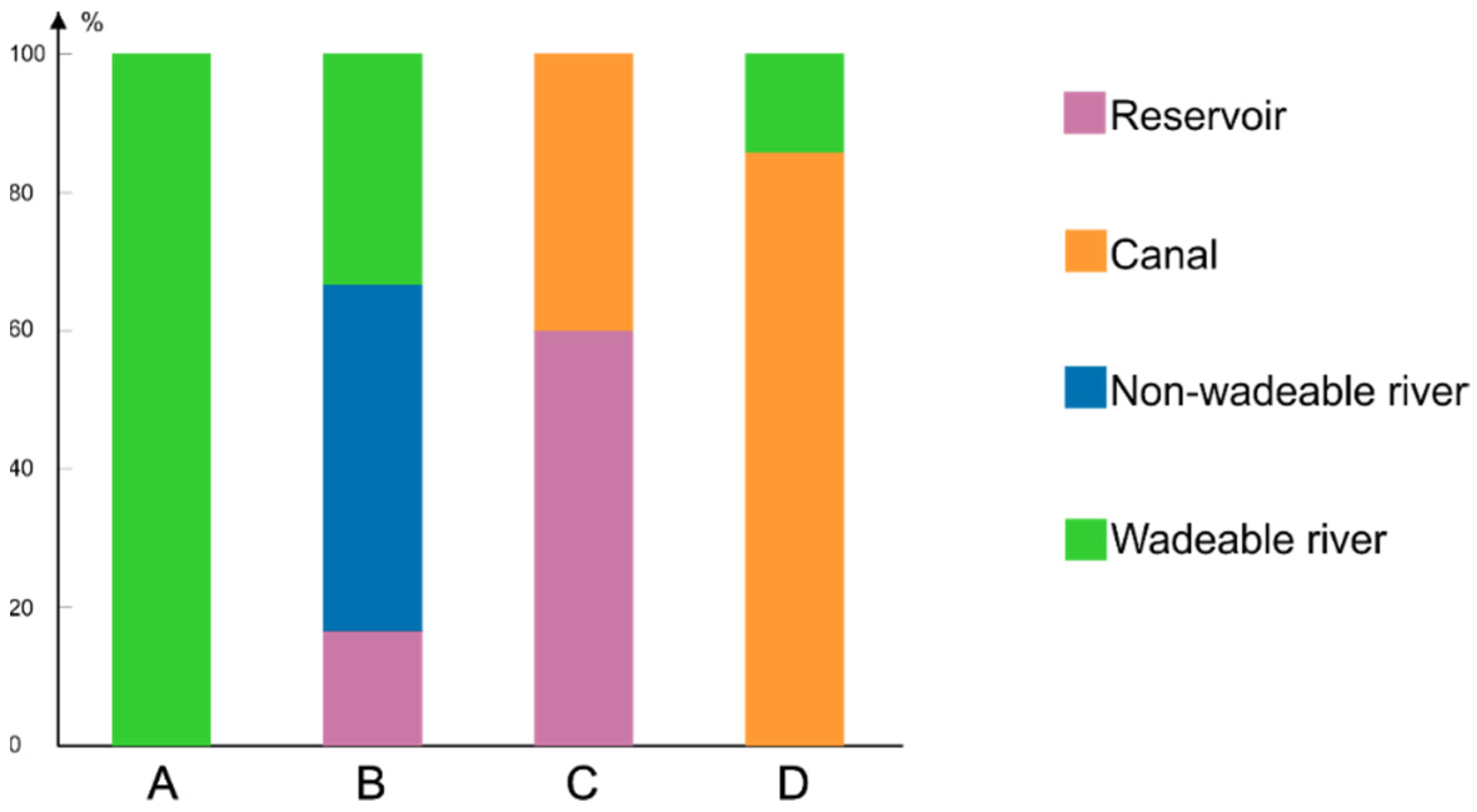
3.1. Chironomid Assemblage Classification
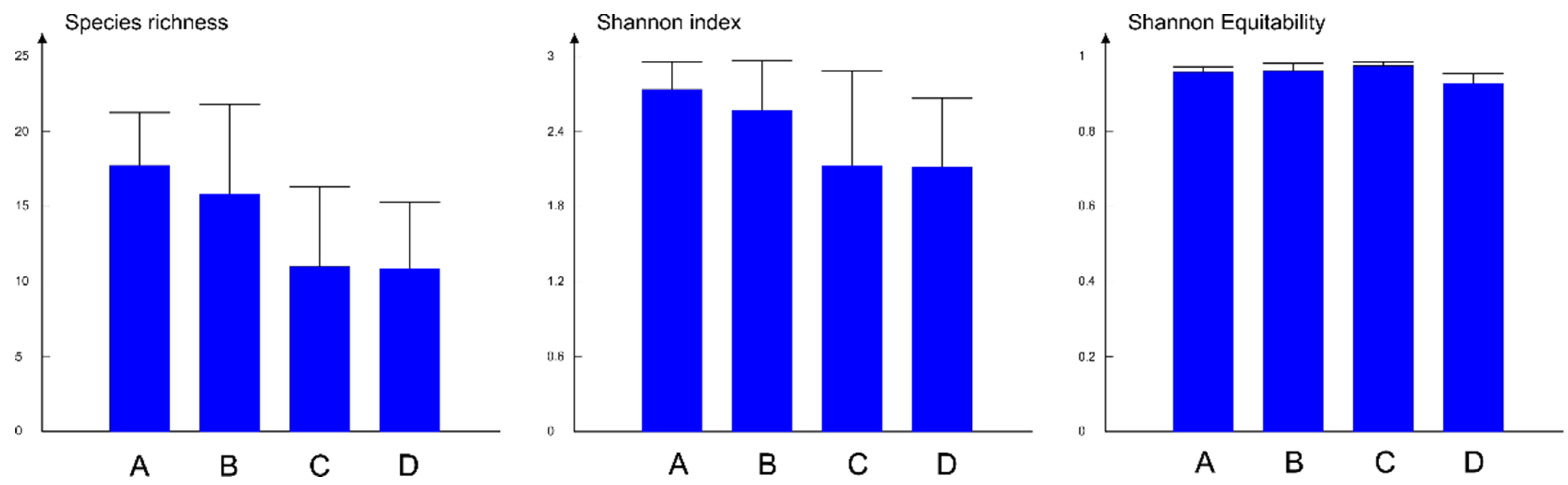
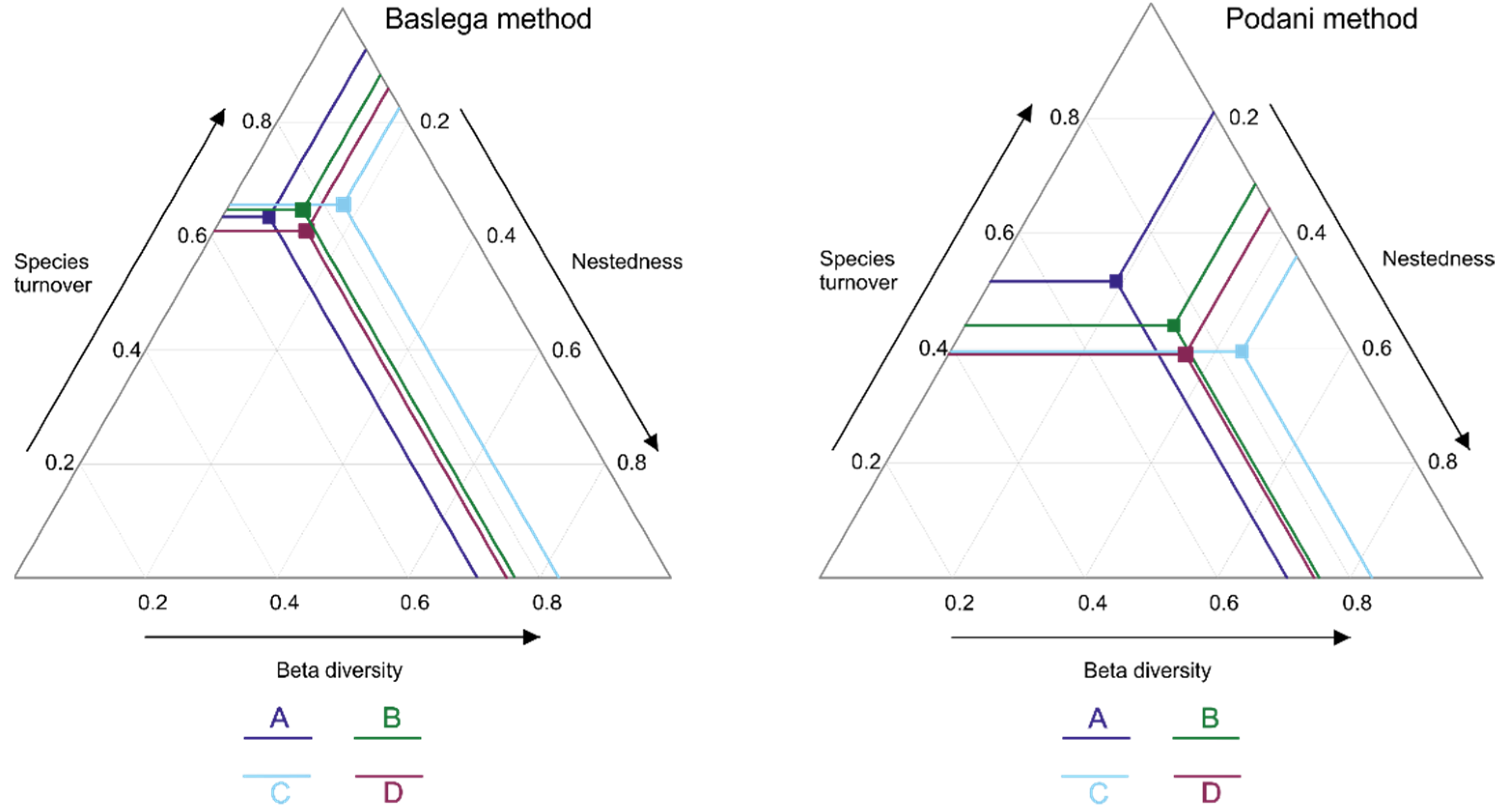
3.2. Diversity Components
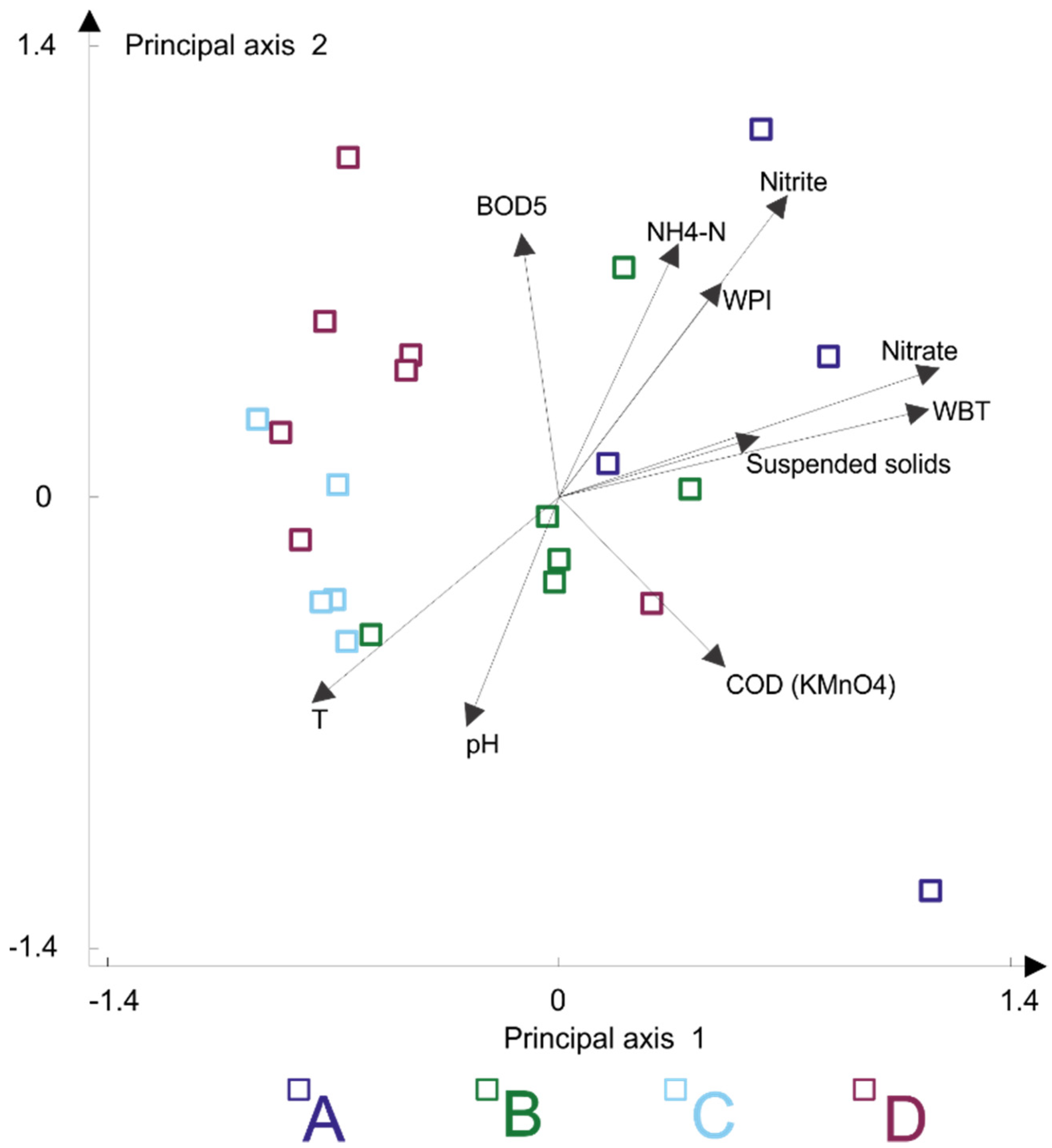
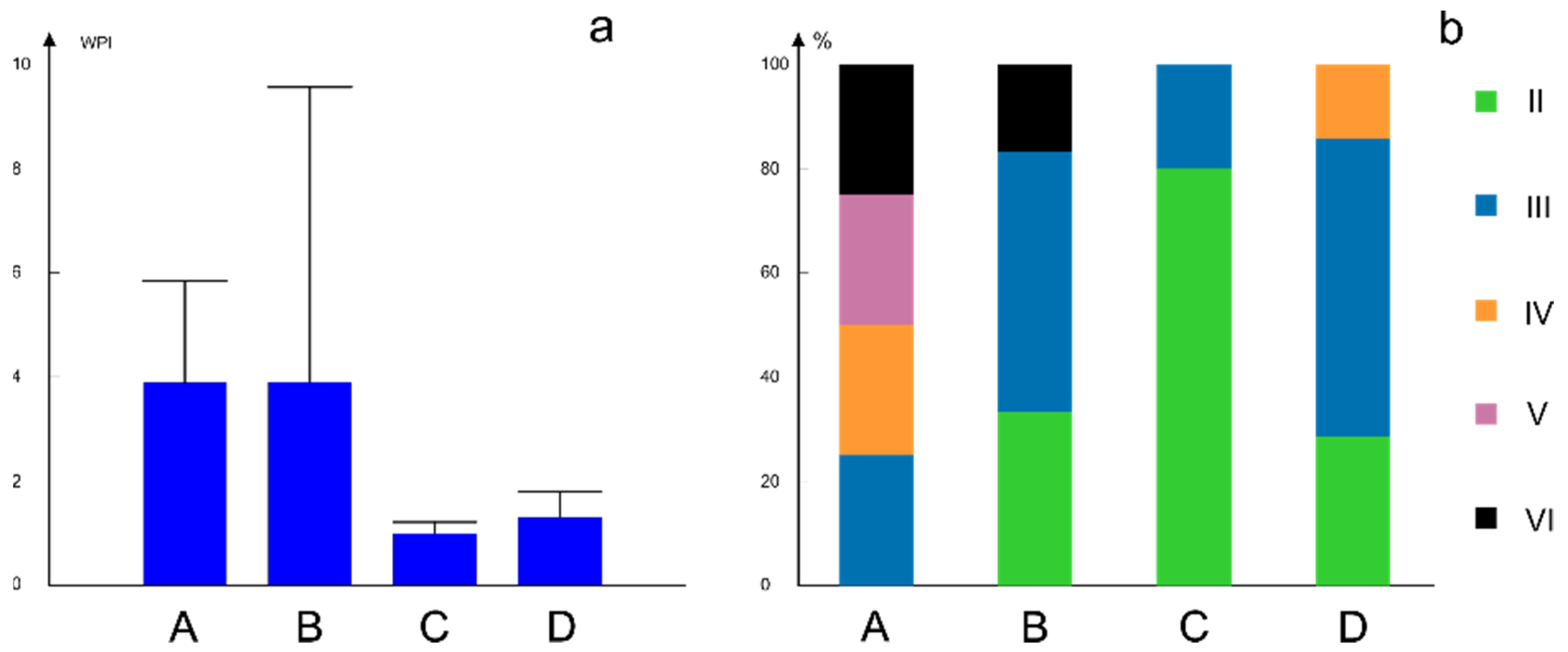
3.3. Patterns in the Chironomidae-Environment Relationship
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyke, A.P.; Hershey, A.E. Effects of fish predation on larval chironomid (Diptera: Chironomidae) communities in an arctic ecosystem. Hydrobiologia 1992, 240, 203–211. [Google Scholar] [CrossRef]
- Armitage, P.D.; Cranston, P.S.; Pinder, L.C.V. The Chironomidae: Biology and Ecology of Non-Biting Midges; Chapman & Hall: London, UK, 1995; ISBN 10041245260X. [Google Scholar]
- Martínez-Sánchez, A.; Allende, A.; Bennett, R.N.; Ferreres, F.; Gil, M.I. Microbial, nutritional and sensory quality of rocket leaves as affected by different sanitizers. Postharvest Biol. Technol. 2006, 42, 86–97. [Google Scholar] [CrossRef]
- Benke, A.C. Production dynamics of riverine chironomids: Extremely high biomass turnover rates of primary consumers. Ecology 1998, 79, 899–910. [Google Scholar] [CrossRef]
- Bogut, I.; Čerba, D.; Vidaković, J.; Gvozdić, V. Interactions of weed-bed invertebrates and Ceratophyllum demersum stands in a floodplain lake. Biologia 2010, 65, 113–121. [Google Scholar] [CrossRef]
- Čerba, D.; Mihaljević, Z.; Vidaković, J. Colonisation of temporary macrophyte substratum by midges (Chironomidae: Diptera). Ann. Limnol. 2010, 46, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Leszczyńska, J.; Grzybkowska, M.; Głowacki, Ł.; Dukowska, M. Environmental Variables Influencing Chironomid Assemblages (Diptera: Chironomidae) in Lowland Rivers of Central Poland. Environ. Entomol. 2019, 48, 988–997. [Google Scholar] [CrossRef]
- Milošević, D.; Čerba, D.; Szekeres, J.; Csányi, B.; Tubić, B.; Simić, V.; Paunović, M. Artificial neural networks as an indicator search engine: The visualization of natural and man-caused taxa variability. Ecol. Indic. 2016, 61, 777–789. [Google Scholar] [CrossRef]
- Milošević, D.; Mančev, D.; Čerba, D.; Piperac, M.S.; Popović, N.; Atanacković, A.; Đuknić, J.; Simić, V.; Paunović, M. The potential of chironomid larvae-based metrics in the bioassessment of non-wadeable rivers. Sci. Total Environ. 2018, 616, 472–479. [Google Scholar] [CrossRef]
- Milošević, D.; Simić, V.; Stojković, M.; Čerba, D.; Mančev, D.; Petrović, A.; Paunović, M. Spatio-temporal pattern of the Chironomidae community: Toward the use of non-biting midges in bioassessment programs. Aquat. Ecol. 2013, 47, 37–55. [Google Scholar] [CrossRef]
- Savić-Zdravković, D.; Jovanović, B.; Đurđević, A.; Stojković-Piperac, M.; Savić, A.; Vidmar, J.; Milošević, D. An environmentally relevant concentration of titanium dioxide (TiO2) nanoparticles induces morphological changes in the mouthparts of Chironomus tentans. Chemosphere 2018, 211, 489–499. [Google Scholar] [CrossRef]
- Stanković, J.; Milošević, D.; Savić-Zdraković, D.; Yalçın, G.; Yildiz, D.; Beklioğlu, M.; Jovanović, B. Exposure to a microplastic mixture is altering the life traits and is causing deformities in the non-biting midge Chironomus riparius Meigen (1804). Environ. Pollut. 2020, 262, 114248. [Google Scholar] [CrossRef] [PubMed]
- Fusari, L.M.; Fonseca-Gessner, A.A. Environmental assessment of two small reservoirs in southeastern Brazil, using macroinvertebrate community metrics. Acta Limnol. Bras. 2006, 18, 89–99. [Google Scholar]
- Martins, R.T.; Stephan, N.N.C.; Alves, R.G. Tubificidae (Annelida: Oligochaeta) as an indicator of water quality in an urban stream in southeast Brazil. Acta Limnol. Bras. 2008, 20, 221–226. [Google Scholar]
- Rosa, B.J.F.V.; Rodrigues, L.F.T.; de Oliveira, G.S.; da Gama Alves, R. Chironomidae and Oligochaeta for water quality evaluation in an urban river in southeastern Brazil. Environ. Monit. Assess. 2014, 186, 7771–7779. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, M.J.; Zhu, W. Aquatic insect diversity in streams across a rural–urban land-use discontinuum. Hydrobiologia 2019, 837, 15–30. [Google Scholar] [CrossRef]
- Qiu, Z.; Kennen, J.G.; Giri, S.; Walter, T.; Kang, Y.; Zhang, Z. Reassessing the relationship between landscape alteration and aquatic ecosystem degradation from a hydrologically sensitive area perspective. Sci. Total Environ. 2019, 650, 2850–2862. [Google Scholar] [CrossRef]
- Wenger, S.J.; Roy, A.H.; Jackson, C.R.; Bernhardt, E.S.; Carter, T.L.; Filoso, S.; Gibson, C.A.; Hession, W.C.; Kaushal, S.S.; Martí, E. Twenty-six key research questions in urban stream ecology: An assessment of the state of the science. J. N. Am. Benthol. Soc. 2009, 28, 1080–1098. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.H.; Rhea, L.K.; Mayer, A.L.; Shuster, W.D.; Beaulieu, J.J.; Hopton, M.E.; Morrison, M.A.; Amand, A.S. How much is enough? Minimal responses of water quality and stream biota to partial retrofit stormwater management in a suburban neighborhood. PLoS ONE 2014, 9, e85011. [Google Scholar] [CrossRef]
- Živadinović, I.; Ilijević, K.; Gržetić, I.; Popović, A. Long-term changes in the eco-chemical status of the Danube River in the region of Serbia. J. Serb. Chem. Soc. 2010, 75, 1125–1148. [Google Scholar] [CrossRef]
- Kolarević, S.; Knežević-Vukčević, J.; Paunović, M.; Tomović, J.; Gačić, Z.; Vuković-Gačić, B. The anthropogenic impact on water quality of the river Danube in Serbia: Microbiological analysis and genotoxicity monitoring. Arch. Biol. Sci. 2011, 63, 1209–1217. [Google Scholar] [CrossRef]
- Jovanović Marić, J.M.; Kračun-Kolarević, M.J.; Kolarević, S.M.; Đorđević, J.Z.; Paunović, M.M.; Kostić-Vuković, J.M.; Sunjog, K.Z.; Smiljanić, P.B.; Gačić, Z.M.; Vuković-Gačić, B.S. Sensitivity of Bleak (Alburnus alburnus) in Detection of the Wastewater Related Pressure in Large Lowland Rivers. Bull. Environ. Contam. Toxicol. 2020, 105, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Official Gazette of the Republic of Serbia. The Parameters of Ecological and Chemical Status of Surface Waters and Parameters of the Chemical and Quantitative Status of Groundwater; Off. Gaz. RS No. 74/2011; Official Gazette of the Republic of Serbia: Belgrade, Serbia, 2011. [Google Scholar]
- AQEM Consortium. Manual for the Application of the AQEM System; Version 1.0; Contract No: EVK1-CT1999-00027; AQEM: Duisburg, Germany, 2002. [Google Scholar]
- Schmid, P.-E. A Comprehensive Method to Assess European Streams Using Benthic Macroinvertebrates, Developed for the Purpose of the Water Framework Directive. In A Key to the Larval Chironomidae and Their Instars from Austrian Danube Region Streams and Rivers with Particular Reference to a Numerical Taxonomic Approach; Federal Institute for Water Quality of the Ministry of Agriculture and Forestry: Vienna, Austria, 1993; ISBN 390067292X. [Google Scholar]
- Vallenduuk, H.J.; Moller Pillot, H.K.M. Chironomidae Larvae, Vol. 1: Tanypodinae: General Ecology and Tanypodinae; Brill: Leiden, The Netherlands, 2007; ISBN 9004278036. [Google Scholar]
- Maschwitz, D.E.; Cook, E.F. Revision of the Nearctic Species of the Genus Polypedilum Kieffer (Diptera: Chironomidae) in the Subgenera P. (Polypedilum) Kieffer and P. (Uresipedilum) Oyewo and Sæether; Ohio Biological Survey, College of Biological Sciences, The Ohio State: Columbus, OH, USA, 2000; ISBN 0867271302. [Google Scholar]
- Epler, J.H. Identification Guide to the Larvae of the Tribe Tanytarsini (Diptera: Chironomidae) in Florida; Florida Department of Environmental Protection: Tallahassee, FL, USA, 2014. [Google Scholar]
- Moller Pillot, H.K.M. Chironomidae Larvae, Vol. 2: Chironomini: Biology and Ecology of the Chironomini; Brill: Leiden, The Netherlands, 2009; ISBN 9004278044. [Google Scholar]
- Orendt, C.; Spies, M. Chironomini:(Diptera: Chironomodae: Chironominae); Keys to Central European Larvae Using Mainly Macroscopic Characters; Orendt Hydrobiologie: Leipzig, Germany, 2012; ISBN 3000388427. [Google Scholar]
- Andersen, T.; Sæther, O.A.; Cranston, P.S.; Epler, J.H. The larvae of Orthocladiinae (Diptera: Chironomidae) of the Holarctic region-Keys and diagnoses. Insect Syst. Evol. 2013, 66, 189–385. [Google Scholar]
- Bitušík, P.; Hamerlík, L. Príručka na Určovanie Lariev Pakomárov (Diptera: Chironomidae) Slovenska; Čast’2. Tanypodinae; Belianum Vydav. Univerzity Matej Bela v Banskej Bystrici: Banska Bystrica, Slovakia, 2014. [Google Scholar]
- Vallenduuk, H.J. Chironomini Larvae of Western European Lowlands (Diptera: Chironomidae): Keys with Notes to the Species: With a Redescription of Glyptotendipes (Caulochironomus) Nagorskayae and a First Description of Glyptotendipes (Caulochironomus) Kaluginae New Species; Erik Mauch: Dinkelscherben, Germany, 2017. [Google Scholar]
- Popovic, N.Z.; Duknic, J.A.; Atlagic, J.Z.; Rakovic, M.J.; Marinkovic, N.S.; Tubic, B.P.; Paunovic, M.M. Application of the water pollution index in the assessment of the ecological status of rivers: A case study of the Sava River, Serbia. Acta Zool. Bulg. 2016, 68, 97–102. [Google Scholar]
- Armitage, P.D.; Moss, D.; Wright, J.F.; Furse, M.T. The performance of a new biological water quality score system based on macroinvertebrates over a wide range of unpolluted running-water sites. Water Res. 1983, 17, 333–347. [Google Scholar] [CrossRef]
- Zelinka, M.; Marvan, P. Zur Prazisierung der biologischen Klassifikation des Reinheit fliessender Gewasser. Arch. Hydrobiol. 1961, 57, 389–407. [Google Scholar]
- Moog, O. Fauna Aquatica Austriaca Katalog zur autökologischen Einstufung aquatischer Organismen Österreichs; Bundesministerium für Nachhaltigkeit und Tourismus: Vienna, Austria, 2002; p. 670. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Champaign, IL, USA, 1949; pp. 1–117. [Google Scholar]
- MacQueen, J. Some methods for classification and analysis of multivariate observations. In Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability; University of California Press: Oakland, CA, USA, 1967; Volume 1, pp. 281–297. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: Berlin/Heidelberg, Germany, 2013; Volume 112. [Google Scholar]
- Hill, M.O.; Bunce, R.G.H.; Shaw, M.W. Indicator Species Analysis, A Divisive Polythetic Method of Classification, and its Application to a Survey of Native Pinewoods in Scotland. J. Ecol. 1975, 63, 597–613. [Google Scholar] [CrossRef]
- Marinković, N.; Karadžić, B.; Pešić, V.; Gligorović, B.; Grosser, C.; Paunović, M.; Nikolić, V.; Raković, M. Faunistic patterns and diversity components of leech assemblages in karst springs of Montenegro. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 26. [Google Scholar] [CrossRef]
- Bray, J.H.; Maxwell, S.E.; Maxwell, S.E. Multivariate Analysis of Variance; SAGE: Thousand Oaks, CA, USA, 1985; ISBN 0803923104. [Google Scholar]
- Ward, J.H., Jr.; Hook, M.E. Application of an hierarchical grouping procedure to a problem of grouping profiles. Educ. Psychol. Meas. 1963, 23, 69–81. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Podani, J.; Ricotta, C.; Schmera, D. A general framework for analyzing beta diversity, nestedness and related community-level phenomena based on abundance data. Ecol. Complex. 2013, 15, 52–61. [Google Scholar] [CrossRef] [Green Version]
- ter Braak, C.J.F. Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Karadžić, B. FLORA: A software package for statistical analysis of ecological data. Water Res. Manag. 2013, 3, 45–54. [Google Scholar]
- Sekulić, D.; Karadžić, B.; Kuzmanović, N.; Jarić, S.; Mitrović, M.; Pavlović, P. Diversity of ostrya carpinifolia forests in ravine habitats of serbia (S-e Europe). Diversity 2021, 13, 59. [Google Scholar] [CrossRef]
- Leszczyńska, J.; Głowacki; Grzybkowska, M.; Przybylski, M. Chironomid riverine assemblages at the regional temperate scale–compositional distance and species diversity. Eur. Zool. J. 2021, 88, 731–748. [Google Scholar] [CrossRef]
- Orendt, C. Results of 10 years sampling of Chironomidae from German lowland running waters differing in degradation. J. Limnol. 2018, 77 (Suppl. S1), 169–176. [Google Scholar] [CrossRef] [Green Version]
- Zilli, F.L.; Montalto, L.; Marchese, M.R. Benthic invertebrate assemblages and functional feeding groups in the Paraná River floodplain (Argentina). Limnologica 2008, 38, 159–171. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, E.M.; Uieda, V.S.; da Motta, R.L. Colonization of rocky and leaf pack substrates by benthic macroinvertebrates in a stream in Southeast Brazil. Títulos Não-Correntes 2008, 22, 37–44. [Google Scholar]
- Vermonden, K.; Brodersen, K.P.; Jacobsen, D.; Van Kleef, H.; Van der Velde, G.; Leuven, R. The influence of environmental factors and dredging on chironomid larval diversity in urban drainage systems in polders strongly influenced by seepage from large rivers. J. N. Am. Benthol. Soc. 2011, 30, 1074–1092. [Google Scholar] [CrossRef]
- Jiang, X.; Xiong, J.; Xie, Z. Longitudinal and seasonal patterns of macroinvertebrate communities in a large undammed river system in Southwest China. Quat. Int. 2017, 440, 1–12. [Google Scholar] [CrossRef]
- Moller Pillot, H.K.M. 2 General Aspects of the Systematics, Biology and Ecology of the Aquatic Orthocladiinae. In Chironomidae Larvae, Vol. 3: Orthocladiinae; KNNV: Zeist, The Netherlands, 2013; pp. 7–21. ISBN 9004278052. [Google Scholar]
- Hynes, H.B.N. The Biology of Polluted Waters, 1st ed.; Liverpool University Press: Liverpool, UK, 1960. [Google Scholar]
- Calle-Martínez, D.; Casas, J.J. Chironomid species, stream classification, and water-quality assessment: The case of 2 Iberian Mediterranean mountain regions. J. N. Am. Benthol. Soc. 2006, 25, 465–476. [Google Scholar] [CrossRef]
- De Haas, E.M.; Paumen, M.L.; Koelmans, A.A.; Kraak, M.H.S. Combined effects of copper and food on the midge Chironomus riparius in whole-sediment bioassays. Environ. Pollut. 2004, 127, 99–107. [Google Scholar] [CrossRef]
- Raunio, J.; Paavola, R.; Muotka, T. Effects of emergence phenology, taxa tolerances and taxonomic resolution on the use of the Chironomid Pupal Exuvial Technique in river biomonitoring. Freshw. Biol. 2007, 52, 165–176. [Google Scholar] [CrossRef]
- McCord, S.B.; Kuhl, B.A. Macroinvertebrate community structure and its seasonal variation in the Upper Mississippi River, USA: A case study. J. Freshw. Ecol. 2013, 28, 63–78. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Quality Class | WPI | |
|---|---|---|
| I | very pure | ≤0.3 |
| II | pure | 0.3–1.0 |
| III | moderately polluted | 1.0–2.0 |
| IV | polluted | 2.0–4.0 |
| V | impure | 4.0–6.0 |
| VI | heavily impure | >6.0 |
| K-Means Clustering | Bayesian Classification | |||||
|---|---|---|---|---|---|---|
| No. of clusters | WGV | BGV | / | WGV | BGV | / |
| 2 | 1.1361 | 0.0522 | 0.0459 | 1.1346 | 0.0591 | 0.0521 |
| 3 | 1.5603 | 0.1009 | 0.0647 | 1.5768 | 0.0980 | 0.0621 |
| 4 | 1.9335 | 0.1404 | 0.0726 | 1.9562 | 0.1341 | 0.0685 |
| 5 | 2.3112 | 0.1610 | 0.0696 | 2.2934 | 0.1599 | 0.0697 |
| Variable | Eigenvalue | F Statistic | Probability |
|---|---|---|---|
| nutrients | |||
| NO3 | 0.383 | 2.357 | 0.000 |
| NH4-N | 0.244 | 1.445 | 0.004 |
| NO2 | 0.274 | 1.639 | 0.009 |
| Total nitrogen | 0.109 | 0.622 | 0.648 |
| Cl | 0.133 | 0.765 | 0.688 |
| Total phosphate | 0.083 | 0.469 | 0.804 |
| oxygen status parameters | |||
| BOD5 | 0.279 | 1.667 | 0.001 |
| COD (KMnO4) | 0.294 | 1.768 | 0.005 |
| O2% | 0.199 | 1.166 | 0.052 |
| O2 | 0.202 | 1.180 | 0.100 |
| TOC | 0.226 | 1.334 | 0.296 |
| physical parameters | |||
| WBT | 0.380 | 2.341 | 0.000 |
| pH | 0.281 | 1.681 | 0.003 |
| WPI | 0.259 | 1.539 | 0.003 |
| Suspended solids | 0.234 | 1.381 | 0.010 |
| T | 0.175 | 1.013 | 0.034 |
| Electrical conductivity | 0.161 | 0.929 | 0.087 |
| Dry residue | 0.108 | 0.618 | 0.344 |
| WPI | WBT | |
|---|---|---|
| Suspended solids | 0.000 | 0.011 |
| Total nitrogen | 0.000 | 0.059 |
| NH4-N | 0.000 | 0.077 |
| BOD5 | 0.000 | 0.932 |
| NO2 | 0.013 | 0.017 |
| Total phosphate | 0.017 | 0.315 |
| NO3 | 0.018 | 0.000 |
| WBT | 0.024 | |
| WPI | 0.024 | |
| T | 0.074 | 0.000 |
| TOC | 0.218 | 0.481 |
| O2% | 0.222 | 0.648 |
| pH | 0.231 | 0.061 |
| O2 | 0.287 | 0.903 |
| COD (KMnO4) | 0.492 | 0.341 |
| Electrical conductivity | 0.601 | 0.470 |
| Cl | 0.741 | 0.687 |
| Dry residue | 0.742 | 0.417 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popović, N.; Marinković, N.; Čerba, D.; Raković, M.; Đuknić, J.; Paunović, M. Diversity Patterns and Assemblage Structure of Non-Biting Midges (Diptera: Chironomidae) in Urban Waterbodies. Diversity 2022, 14, 187. https://doi.org/10.3390/d14030187
Popović N, Marinković N, Čerba D, Raković M, Đuknić J, Paunović M. Diversity Patterns and Assemblage Structure of Non-Biting Midges (Diptera: Chironomidae) in Urban Waterbodies. Diversity. 2022; 14(3):187. https://doi.org/10.3390/d14030187
Chicago/Turabian StylePopović, Nataša, Nikola Marinković, Dubravka Čerba, Maja Raković, Jelena Đuknić, and Momir Paunović. 2022. "Diversity Patterns and Assemblage Structure of Non-Biting Midges (Diptera: Chironomidae) in Urban Waterbodies" Diversity 14, no. 3: 187. https://doi.org/10.3390/d14030187
APA StylePopović, N., Marinković, N., Čerba, D., Raković, M., Đuknić, J., & Paunović, M. (2022). Diversity Patterns and Assemblage Structure of Non-Biting Midges (Diptera: Chironomidae) in Urban Waterbodies. Diversity, 14(3), 187. https://doi.org/10.3390/d14030187