
The Impact of a Severe El Niño Event on Vascular Epiphytes in Lowland Panama



Abstract
:1. Introduction
2. Materials and Methods
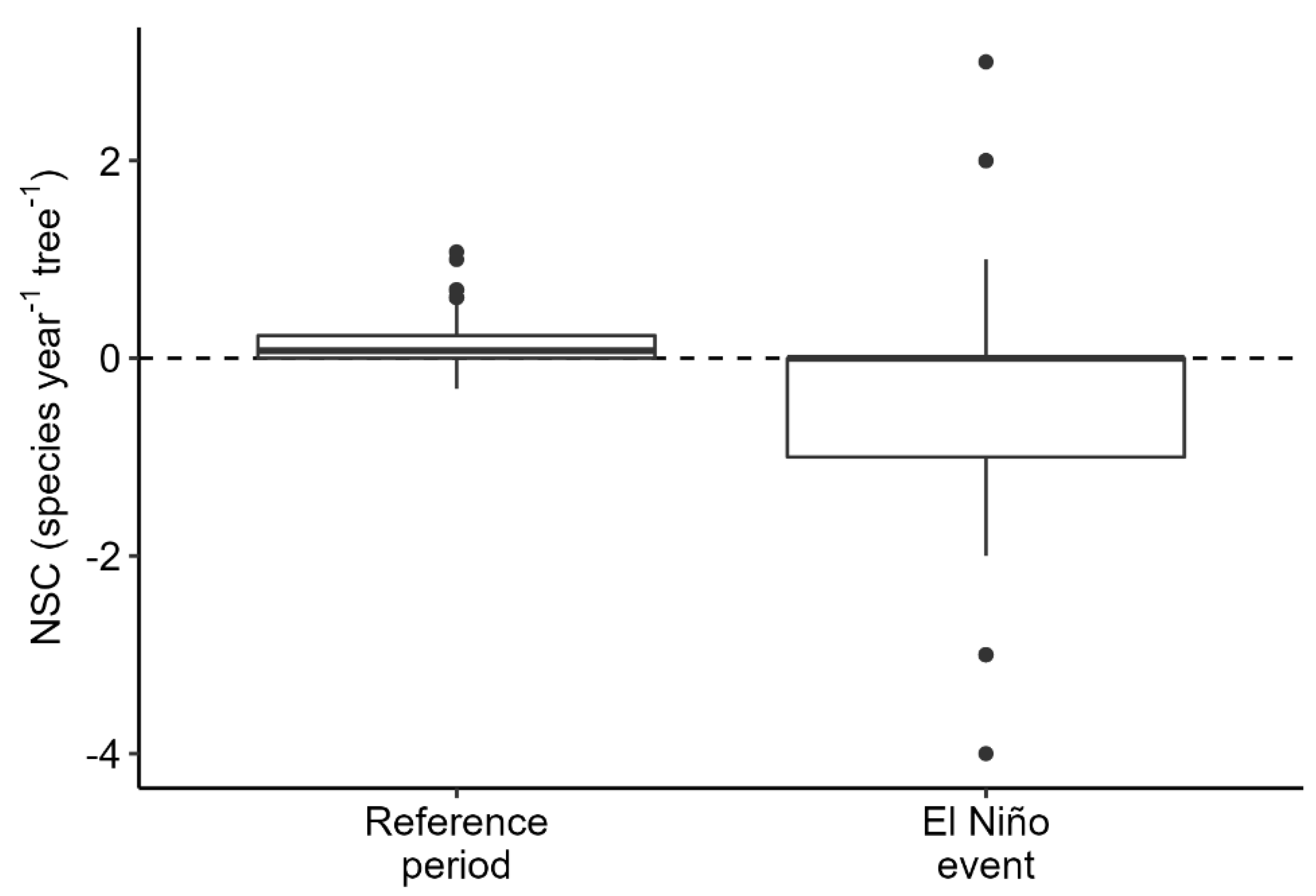
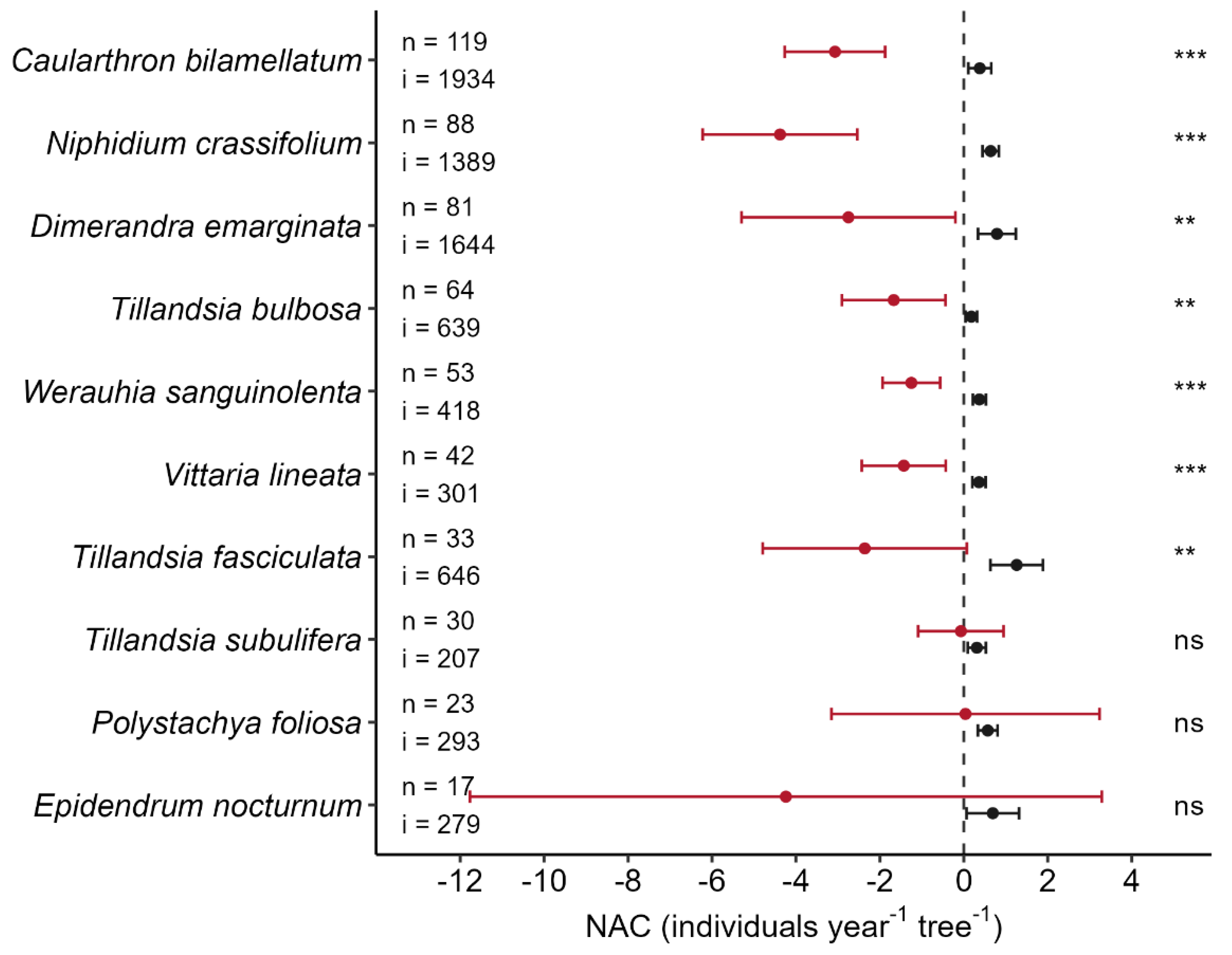
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, W.; Wang, G.; Dewitte, B.; Wu, L.; Santoso, A.; Takahashi, K.; Yang, Y.; Carréric, A.; McPhaden, M.J. Increased variability of eastern Pacific El Niño under greenhouse warming. Nature 2018, 564, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Leigh, E.G.; Windsor, D.M.; Rand, A.S.; Foster, R.B. The impact of the “El Niño” drought of 1982–83 on a Panamanian semideciduous forest. In Elsevier Oceanography Series; Glynn, P.W., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; Volume 52, pp. 473–486. [Google Scholar]
- Nakagawa, M.; Tanaka, K.; Nakashizuka, T.; Ohkubo, T.; Kato, T.; Maeda, T.; Sato, K.; Miguchi, H.; Nagamasu, H.; Ogino, K.; et al. Impact of severe drought associated with the 1997–1998 El Niño in a tropical forest in Sarawak. J. Trop. Ecol. 2000, 16, 355–367. [Google Scholar] [CrossRef]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragão Luiz, E.O.C.; Anderson Liana, O.; Myneni Ranga, B.; Nemani, R. Persistent effects of a severe drought on Amazonian forest canopy. Proc. Natl. Acad. Sci. USA 2013, 110, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, G.B.; Laurance, W.F.; Oliveira, A.A.; Delamônica, P.; Gascon, C.; Lovejoy, T.E.; Pohl, L. Amazonian tree mortality during the 1997 El Niño drought. Conserv. Biol. 2000, 14, 1538–1542. [Google Scholar] [CrossRef] [Green Version]
- Condit, R.; Aguilar, S.; Hernandez, A.; Perez, R.; Lao, S.; Angehr, G.; Hubbell, S.; Foster, R. Tropical forest dynamics across a rainfall gradient and the impact of an El Nino dry season. J. Trop. Ecol. 2004, 20, 51–72. [Google Scholar] [CrossRef] [Green Version]
- Nepstad, D.C.; Tohver, I.M.; Ray, D.; Moutinho, P.; Cardinot, G. Mortality of large trees and lianas following experimental drought in an Amazon forest. Ecology 2007, 88, 2259–2269. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Bongers, F. Increasing liana abundance and biomass in tropical forests: Emerging patterns and putative mechanisms. Ecol. Lett. 2011, 14, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A.; van der Heijden, G.M.F. Lianas have a seasonal growth advantage over co-occurring trees. Ecology 2019, 100, e02655. [Google Scholar] [CrossRef]
- Lee, T.; McPhaden, M.J. Increasing intensity of El Niño in the central-equatorial Pacific. Geophys. Res. Lett. 2010, 37, L14603. [Google Scholar] [CrossRef]
- Lugo, A.E.; Scatena, F.N. Epiphytes and climate change research in the Caribbean: A proposal. Selbyana 1992, 13, 123–130. [Google Scholar]
- Benzing, D.H. Vulnerabilities of tropical forests to climate change: The significance of resident epiphytes. Clim. Chang. 1998, 39, 519–540. [Google Scholar] [CrossRef]
- Zotz, G.; Bader, M.Y. Epiphytic plants in a changing world—Global change effects on vascular and non-vascular epiphytes. Prog. Bot. 2009, 70, 147–170. [Google Scholar] [CrossRef]
- Zotz, G.; Schmidt, G. Population decline in the epiphytic orchid Aspasia principissa. Biol. Conserv. 2006, 129, 82–90. [Google Scholar] [CrossRef]
- Zotz, G.; Laube, S.; Schmidt, G. Long-term population dynamics of the epiphytic bromeliad, Werauhia sanguinolenta. Ecography 2005, 28, 806–814. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Mortality rates of 205 neotropical tree and shrub species and the impact of severe drought. Ecol. Monogr. 1995, 65, 419–439. [Google Scholar] [CrossRef]
- Einzmann, H.J.R.; Weichgrebe, L.; Zotz, G. Long-term community dynamics in vascular epiphytes on Annona glabra along the shoreline of Barro Colorado Island, Panama. J. Ecol. 2021, 109, 1931–1946. [Google Scholar] [CrossRef]
- Gotsch, S.G.; Dawson, T.E.; Draguljić, D. Variation in the resilience of cloud forest vascular epiphytes to severe drought. New Phytol. 2018, 219, 900–913. [Google Scholar] [CrossRef] [Green Version]
- Laube, S.; Zotz, G. A metapopulation approach to the analysis of long-term changes in the epiphyte vegetation on the host tree Annona glabra. J. Veg. Sci. 2007, 18, 613–624. [Google Scholar] [CrossRef]
- Zotz, G.; Bermejo, P.; Dietz, H. The epiphyte vegetation of Annona glabra on Barro Colorado Island, Panama. J. Biogeogr. 1999, 26, 761–776. [Google Scholar] [CrossRef]
- Schmit-Neuerburg, V.M. Dynamics of Vascular Epiphyte Vegetation in the Venezuelan Lowland Rain Forest of the Surumoni Crane Project. Ph.D. Thesis, Rheinischen Friedrich-Wilhelms-Universität, Bonn, Germany, 2002. [Google Scholar]
- Laube, S.; Zotz, G. Neither host-specific nor random: Vascular epiphytes on three tree species in a Panamanian lowland forest. Ann. Bot. 2006, 97, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Einzmann, H.J.R.; Zotz, G. “No signs of saturation”: Long-term dynamics of vascular epiphyte communities in a human-modified landscape. Biodivers. Conserv. 2017, 26, 1393–1410. [Google Scholar] [CrossRef]
- Bretfeld, M.; Ewers, B.E.; Hall, J.S. Plant water use responses along secondary forest succession during the 2015–2016 El Niño drought in Panama. New Phytol. 2018, 219, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windsor, D.M. Climate and Moisture Variability in a Tropical Forest: Long-Term Records from Barro Colorado Island, Panamá; Smithsonian Institution Press: Washington, DC, USA, 1990; Volume 29, p. 145. [Google Scholar]
- The Plant List. Version 1.1Published on the Internet. 2013. Available online: http://www.theplantlist.org/ (accessed on 1 February 2021).
- Paton, S. 2017 Meteorological and Hydrological Summary for Barro Colorado Island; Smithsonian Tropical Research Institute: Amador Causeway, Panama, 2018; pp. 1–42. Available online: https://biogeodb.stri.si.edu/physical_monitoring/research/barrocolorado (accessed on 27 February 2019).
- R Core Team. R: A Language and Environment for Statistical Computing, R version 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 1st ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Alfaro-Sánchez, R.; Muller-Landau, H.C.; Wright, S.J.; Camarero, J.J. Growth and reproduction respond differently to climate in three Neotropical tree species. Oecologia 2017, 184, 531–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentry, A.H.; Dodson, C.H. Diversity and biogeography of neotropical vascular epiphytes. Ann. Mo. Bot. Gard. 1987, 74, 205–233. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, N.M.; Solano, R. Potential effects of climate change on canopy communities in a tropical cloud forest: An experimental approach. Oecologia 2002, 131, 580–586. [Google Scholar] [CrossRef]
- Bader, M.Y.; Menke, G.; Zotz, G. Pronounced drought tolerance characterizes the early life stages of the epiphytic bromeliad Tillandsia flexuosa. Funct. Ecol. 2009, 23, 472–479. [Google Scholar] [CrossRef]
- Zotz, G.; Tyree, M.T. Water stress in the epiphytic orchid, Dimerandra emarginata (G. Meyer) Hoehne. Oecologia 1996, 107, 151–159. [Google Scholar] [CrossRef]
- Zotz, G.; Andrade, J.L. Water relations of two co-occurring epiphytic bromeliads. J. Plant Physiol. 1998, 152, 545–554. [Google Scholar] [CrossRef]
- Zotz, G.; Winter, K. Annual carbon balance and nitrogen-use efficiency in tropical C3 and CAM epiphytes. New Phytol. 1994, 126, 481–492. [Google Scholar] [CrossRef]
- Andrade, J.L.; Nobel, P.S. Microhabitats and water relations of epiphytic cacti and ferns in a lowland neotropical forest. Biotropica 1997, 29, 261–270. [Google Scholar] [CrossRef]
- Mendieta-Leiva, G.; Buckley, H.L.; Zotz, G. Directional changes over time in the species composition of tropical vascular epiphyte assemblages. J. Ecol. 2022, 110, 553–568. [Google Scholar] [CrossRef]
- Zuleta, D.; Benavides, A.M.; López-Rios, V.; Duque, A. Local and regional determinants of vascular epiphyte mortality in the Andean mountains of Colombia. J. Ecol. 2016, 104, 841–849. [Google Scholar] [CrossRef] [Green Version]
- Hietz, P. Population dynamics of epiphytes in a Mexican humid montane forest. J. Ecol. 1997, 85, 767–777. [Google Scholar] [CrossRef]
- Zotz, G.; Hietz, P. The physiological ecology of vascular epiphytes: Current knowledge, open questions. J. Exp. Bot. 2001, 52, 2067–2078. [Google Scholar] [CrossRef]
- Schmidt, G.; Zotz, G. Inherently slow growth in two Caribbean epiphytic species: A demographic approach. J. Veg. Sci. 2002, 13, 527–534. [Google Scholar] [CrossRef]
- Zotz, G. Differences in vital demographic rates in three populations of the epiphytic bromeliad, Werauhia sanguinolenta. Acta Oecologica-Int. J. Ecol. 2005, 28, 306–312. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Individuals | % |
|---|---|---|
| Pre El Niño | 8074 | |
| Post El Niño (still present) | 6085 | 75 |
| Changes 2015 to 2016 | ||
| Dead | 841 | −10.4 |
| Missing | 1148 | −14.2 |
| Newly found | 630 | +7.8 |
| Net change | −16.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Einzmann, H.J.R.; Weichgrebe, L.; Zotz, G. The Impact of a Severe El Niño Event on Vascular Epiphytes in Lowland Panama. Diversity 2022, 14, 325. https://doi.org/10.3390/d14050325
Einzmann HJR, Weichgrebe L, Zotz G. The Impact of a Severe El Niño Event on Vascular Epiphytes in Lowland Panama. Diversity. 2022; 14(5):325. https://doi.org/10.3390/d14050325
Chicago/Turabian StyleEinzmann, Helena J. R., Letizia Weichgrebe, and Gerhard Zotz. 2022. "The Impact of a Severe El Niño Event on Vascular Epiphytes in Lowland Panama" Diversity 14, no. 5: 325. https://doi.org/10.3390/d14050325
APA StyleEinzmann, H. J. R., Weichgrebe, L., & Zotz, G. (2022). The Impact of a Severe El Niño Event on Vascular Epiphytes in Lowland Panama. Diversity, 14(5), 325. https://doi.org/10.3390/d14050325