
Biodiversity and Litter Breakdown in a Karstic Freshwater System (Doberdò Lake, Northeast Italy) in Relation to Water Level Fluctuations and Environmental Features


 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
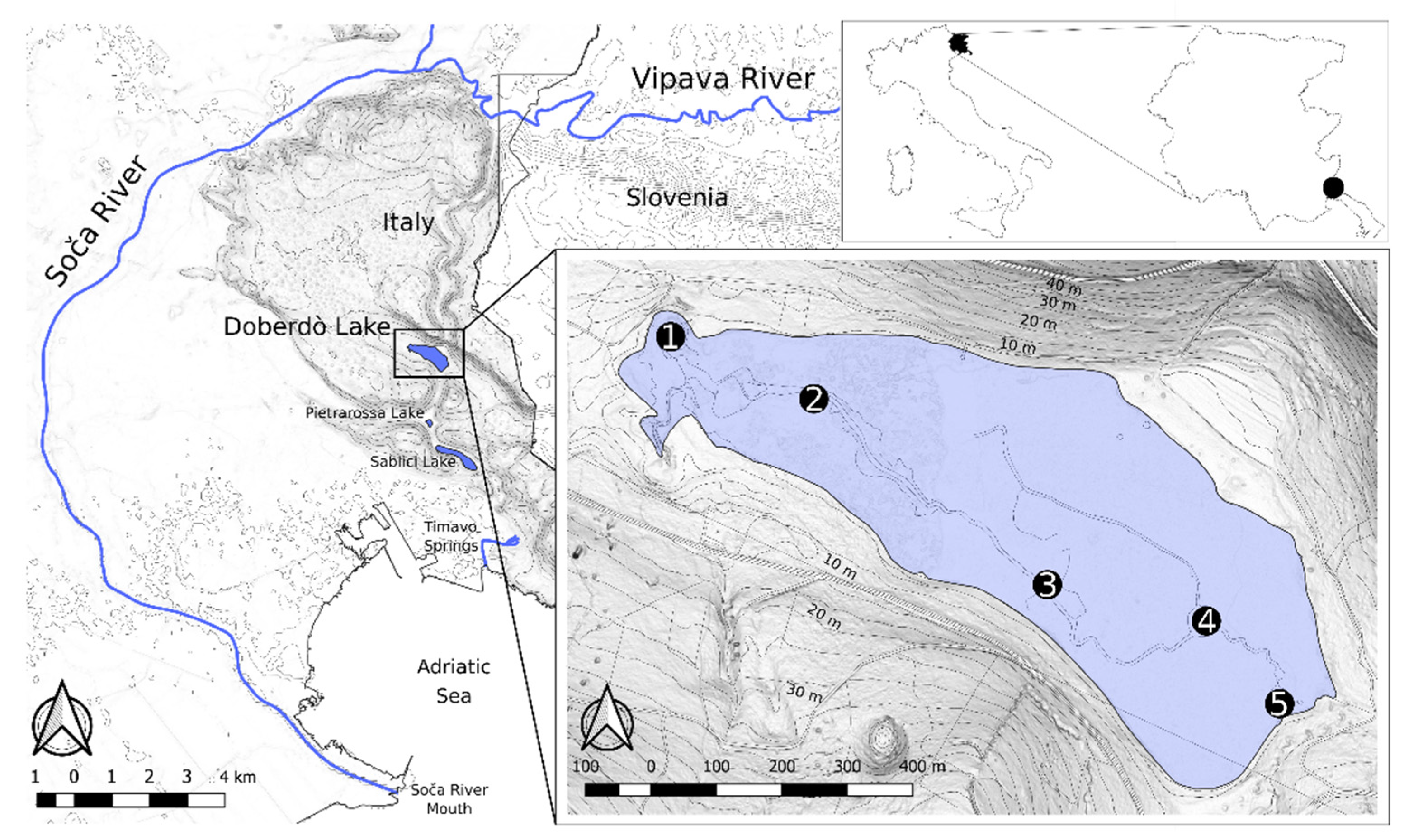
2.1. Study Area
2.2. Chemico-Physical Parameters
2.3. Macrobenthic Invertebrate Sampling through Leaf Bag Technique
2.4. Statistical Analysis
3. Results
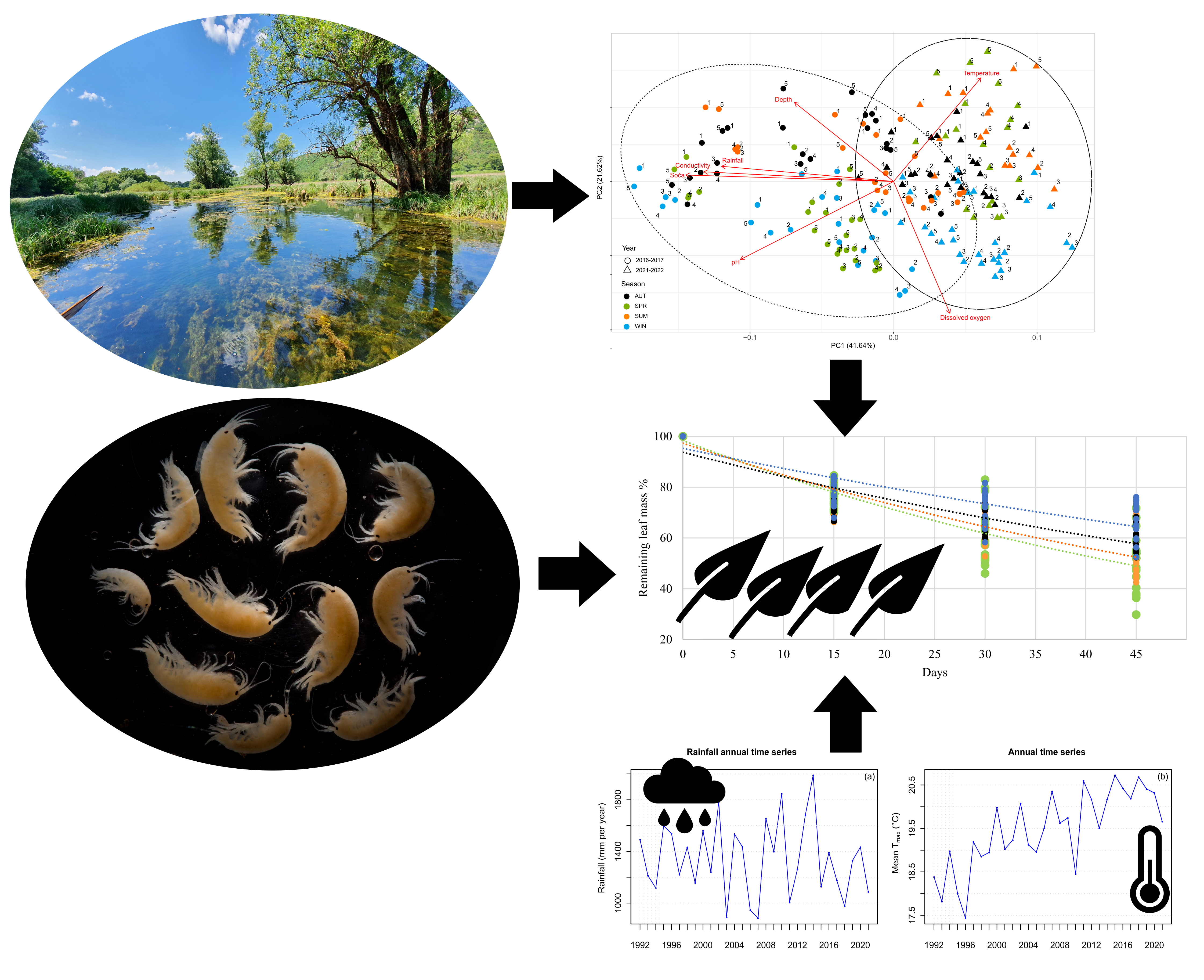
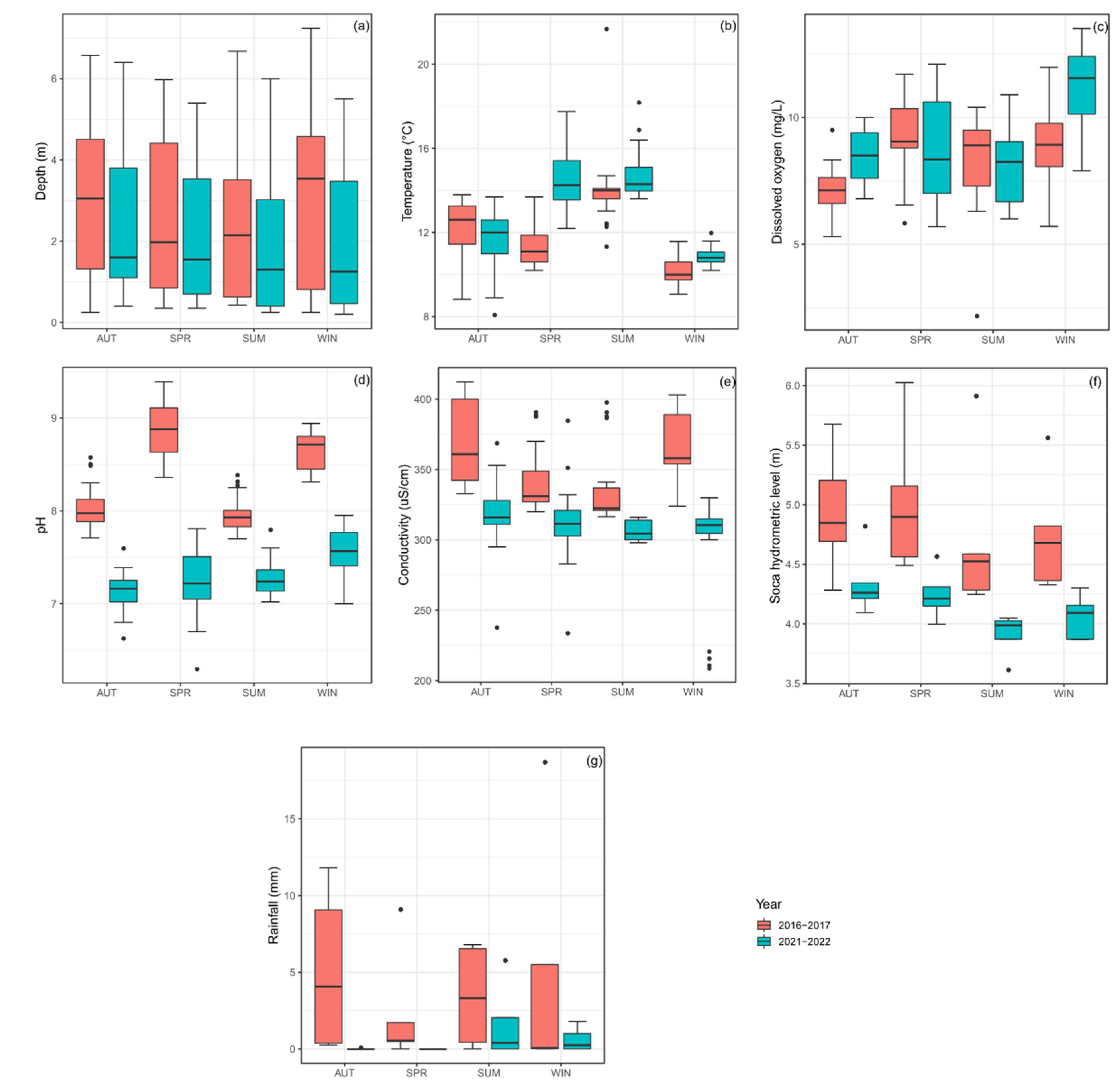
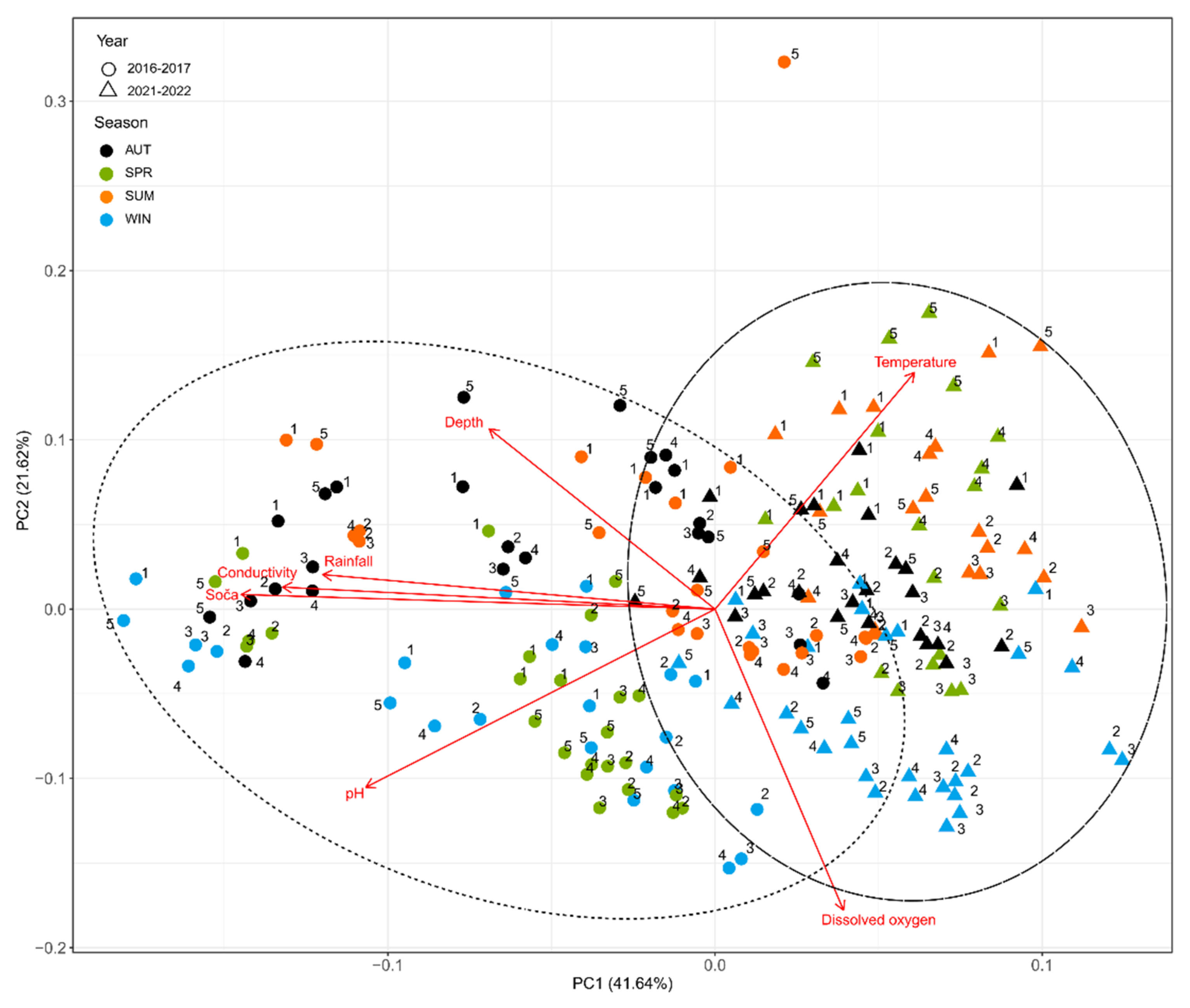
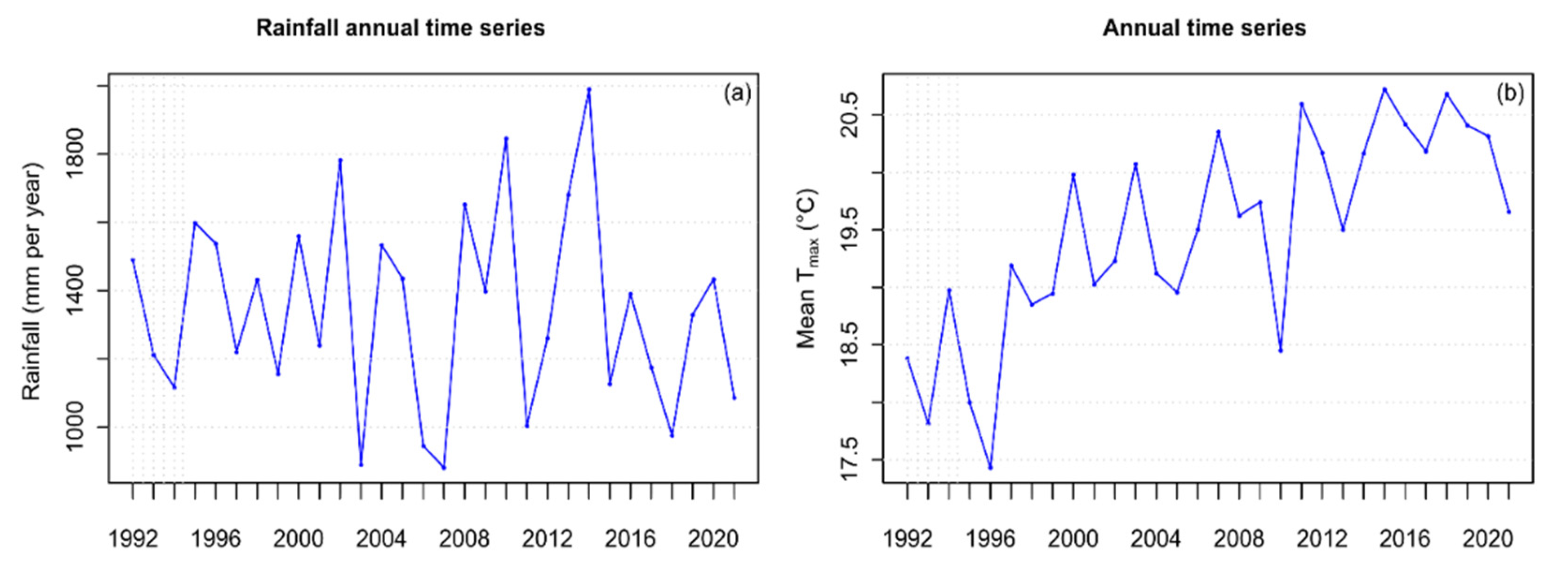
3.1. Physical and Chemical Parameters
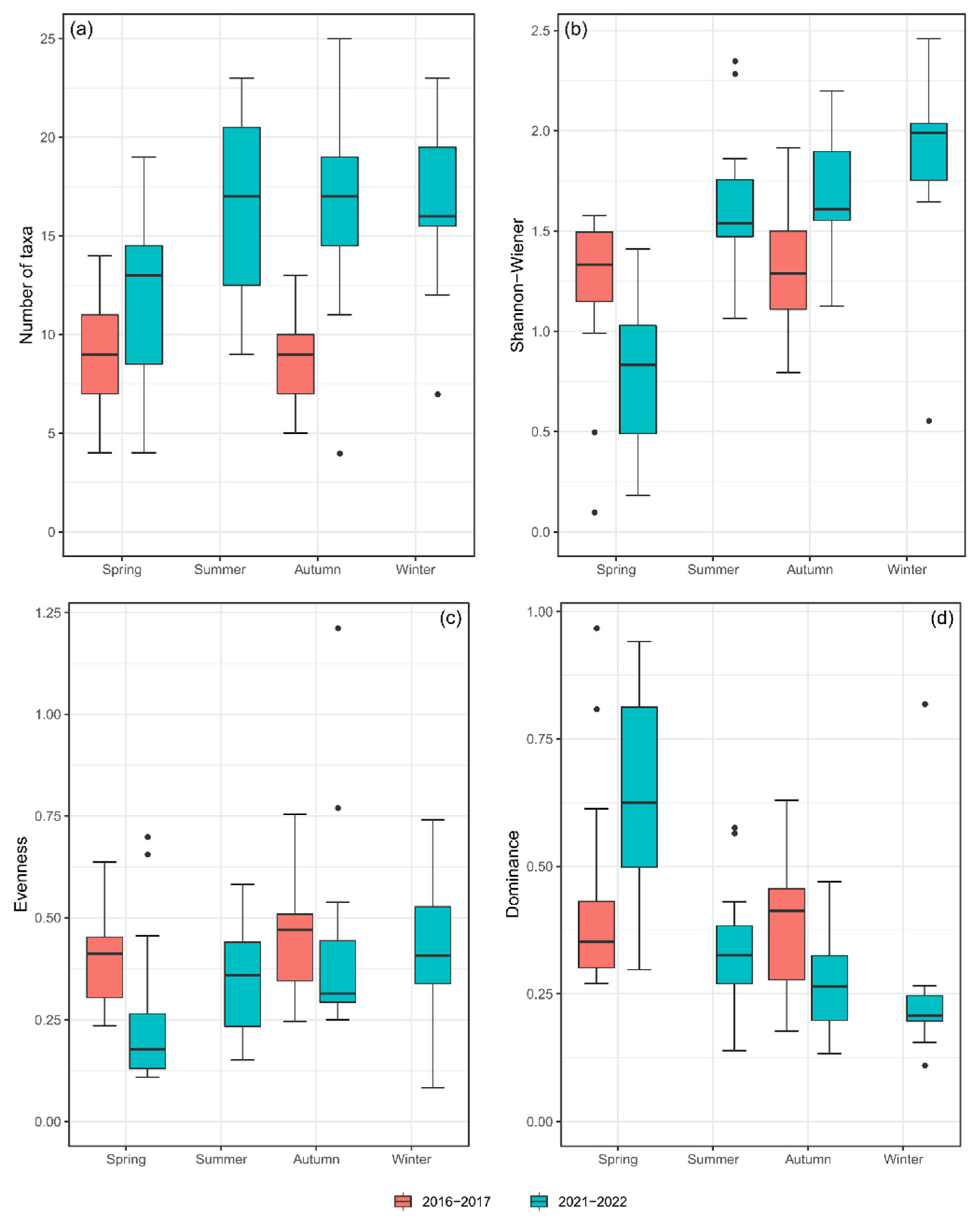
3.2. Macrobenthic Invertebrates
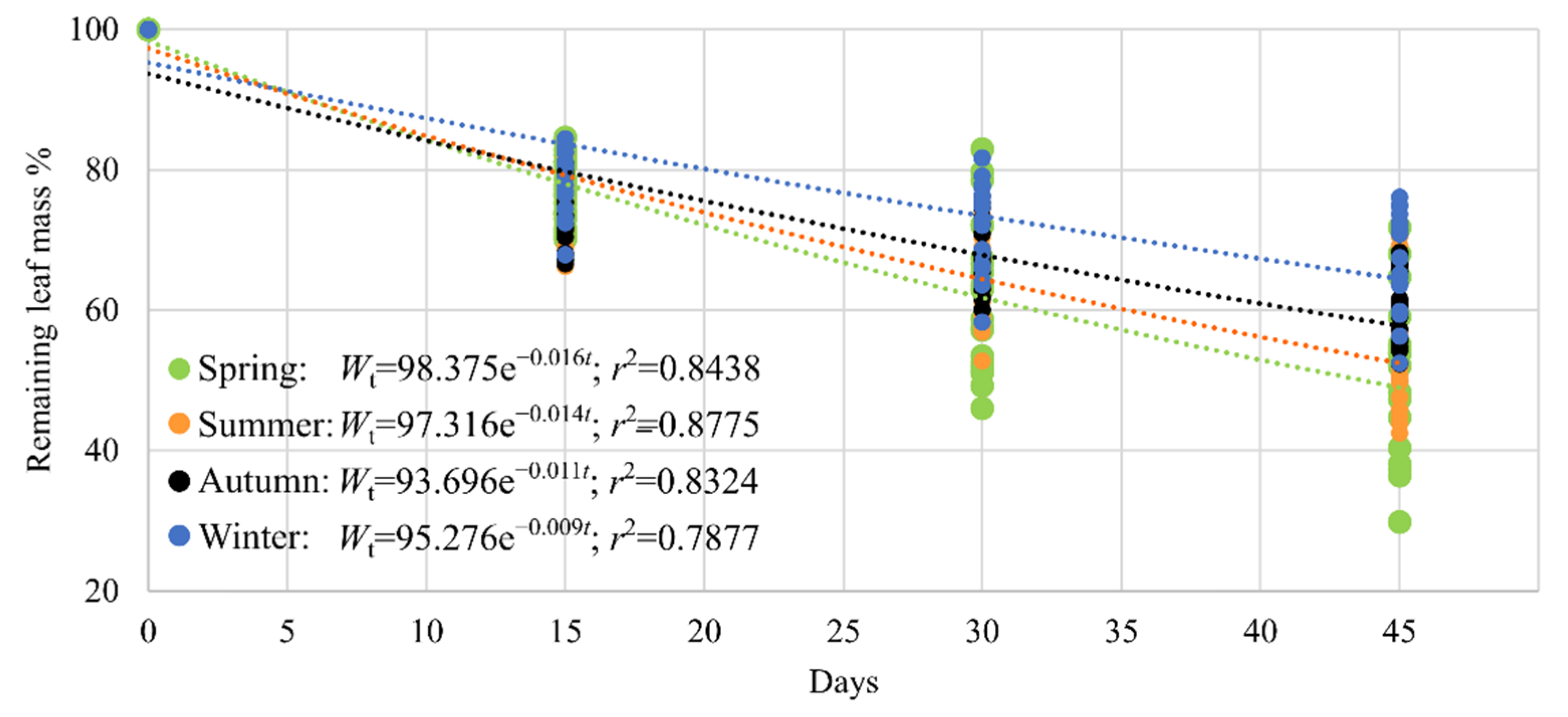
3.3. Decomposition Rates
4. Discussion
4.1. Chemico-Physical Parameters
4.2. Macrobenthic Invertebrates
4.3. Decomposition Rates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gombert, P. Role of karstic dissolution in global carbon cycle. Glob. Planet. Chang. 2002, 33, 177–184. [Google Scholar] [CrossRef]
- Sironić, A.; Baresič, J.; Horvatinčić, N.; Brozinčević, A.; Vurnek, M.; Kapelj, S. Changes in the geochemical parameters of karst lakes over the past three decades—The case of Plitvice Lakes, Croatia. Appl. Geochem. 2017, 78, 12–22. [Google Scholar] [CrossRef]
- Castello, M.; Poldini, L.; Altobelli, A. The aquatic and wetland vegetation of Lake Doberdò: An analysis for conservation value assessment of a disappearing lake of the Classical Karst (Northeast Italy). Plant Sociol. 2021, 58, 75–106. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C. Wetlands in cave and karst regions. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: San Diego, CA, USA, 2019; pp. 1156–1164. [Google Scholar]
- Smith, H.; Wood, P.J.; Gunn, J. The influence of habitat structure and flow permanence on invertebrate communities in karst spring systems. Hydrobiologia 2003, 510, 53–66. [Google Scholar] [CrossRef]
- Tanaka, M.O.; Ribas, A.C.A.; de Souza, A.L.T. Macroinvertebrate succession during leaf litter breakdown in a perennial karstic river in Western Brazil. Hydrobiologia 2006, 568, 493–498. [Google Scholar] [CrossRef]
- Boulton, A.J.; Brock, M.A. Australian Freshwater Ecology: Processes and Management; Gleneagles Publishing: Glen Osmond, Australia, 1999. [Google Scholar]
- Gabersčǐk, A.; Urban-Berčič, O.; Kržič, N.; Kosi, G.; Brancelj, A. The intermittent Lake Cerknica: Various face of the same ecosystem. Lakes Reserv. Res. Manag. 2003, 8, 159–168. [Google Scholar] [CrossRef]
- Dinka, M.; Ágoston-Szabó, E.; Urbanc-Berčič, O.; Germ, M.; Kržič, N.; Gabersčǐk, A. Reed stands conditions at selected wetlands in Slovenia and Hungary. In Wastewater Treatment, Plant Dynamics and Management in Constructed and Natural Wetlands; Vymazal, J., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–12. [Google Scholar]
- Dolinar, N.; Regvar, M.; Abram, D.; Gabersčǐk, A. Water-level fluctuations as a driver of Phragmites australis primary productivity, litter decomposition, and fungal root colonisation in an intermittent wetland. Hydrobiologia 2016, 774, 69–80. [Google Scholar] [CrossRef]
- Yi, Y.; Xie, H.; Yang, Y.; Zhou, Y.; Yang, Z. Suitable habitat mathematical model of common reed (Phragmites australis) in shallow lakes with coupling cellular automaton and modified logistic function. Ecol. Model. 2020, 419, 108938. [Google Scholar] [CrossRef]
- Hill, N.M.; Keddy, P.A.; Wisheu, I.C. A Hydrological model for predicting the effects of dams on the shoreline vegetation of lakes and reservoirs. Environ. Manag. 1998, 22, 723–736. [Google Scholar] [CrossRef]
- Gaberščik, A.; Grašič, M.; Abram, D.; Zelnik, I. Water Level Fluctuations and Air Temperatures Affect Common Reed Habitus and Productivity in an Intermittent Wetland Ecosystem. Water 2020, 12, 2806. [Google Scholar] [CrossRef]
- Dolinar, N.; Šraj, N.; Gabersčǐk, A. Water regime changes and the function of an intermittent wetland. In Water and Nutrient Management in Natural and Constructed Wetlands; Vymazal, J., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 251–262. [Google Scholar]
- Dolinar, N.; Rudolf, M.; Šraj, N.; Gabersčǐk, A. Environmental changes affect ecosystem services of the intermittent Lake Cerknica. Ecol. Complex. 2010, 7, 403–409. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and function of stream ecosystems. Bioscience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Metcalfe Smith, J.L. Biological water quality assessment of rivers: Use of macroinvertebrate communities. In The Rivers Handbook; Calow, P., Petts, G.E., Eds.; Blackwell Science: London, UK, 1994; Volume 2, pp. 144–170. [Google Scholar]
- Bertoli, M.; Piazza, G.; Pastorino, P.; Prearo, M.; Cozzoli, F.; Vignes, F.; Basset, A.; Pizzul, E. Macrobenthic invertebrate energy densities and ecological status in freshwater watercourses (Friuli Venezia-Giulia, Northeast Italy). Aquat. Ecol. 2021, 55, 501–518. [Google Scholar] [CrossRef]
- Hellawell, J.M. Biological Indicators of Freshwater Pollution and Environmental Management; Elsevier: New York, NY, USA, 1986. [Google Scholar]
- Bonada, N.; Prat, N.; Resh, V.; Statzner, B. Developments in aquatic insect biomonitoring: A comparative analysis of recent approaches. Annu. Rev. Entomol. 2006, 51, 495–524. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.L.; Resh, V.H.; Rosenberg, D.M.; Reynoldson, T.B. Biomonitoring in North American rivers: A comparison of methods used for benthic macroinvertebrates in Canada and the United States. In Biological Monitoring of Rivers: Applications and Perspectives; Ziglio, G., Siligardi, M., Flaim, G., Eds.; John Wiley & Sons: New York, NY, USA, 2006; pp. 203–228. [Google Scholar]
- Carter, J.L.; Resh, V.H.; Hannaford, M.J. Macroinvertebrates as biotic indicators of environmental quality. In Methods in Stream Ecology; Hauer, F.R., Lamberti, G.A., Eds.; Ecosystem Structure; Academic Press Elsevier: London, UK, 2017; Volume 1, pp. 293–318. [Google Scholar]
- Liccari, F.; Castello, M.; Poldini, L.; Altobelli, A.; Tordoni, E.; Sigura, M.; Bacaro, G. Do habitats Show a Different Invasibility Pattern by Alien Plant Species? A Test on a Wetland Protected Area. Diversity 2020, 12, 267. [Google Scholar] [CrossRef]
- Bertoli, M.; Franz, F.; Pastorino, P.; Prearo, M.; Pizzul, E. Seasonal patterns of Phragmites australis breakdown in a karstic freshwater system (Doberdò Lake, Northeast Italy) in relation to water level fluctuations, environmental features, and macrobenthic invertebrate communities. Hydrobiologia 2020, 847, 2123–2140. [Google Scholar] [CrossRef]
- Samez, D.; Casagrande, G.; Cucchi, F.; Zini, L. Idrodinamica dei laghi di Doberdò e Pietrarossa (Carso Classico, Italia). Relazioni con le piene dei fiumi Isonzo, Vipacco e Timavo. Atti e Memorie Della Commissione Grotte “E. Boegan”. Trieste 2005, 40, 133–152. [Google Scholar]
- Zini, L.; Calligaris, C.; Zavagno, E. Classical Karst hydrodynamics: A shared aquifer within Italy and Slovenia. In Proceedings of the Evolving Water Resources Systems: Understanding, Predicting and Managing Water–Society Interactions Proceedings of ICWRS2014, Bologna, Italy, 4–6 June 2014. [Google Scholar]
- Doctor, H.D. Hydrology connections and dynamics of water movement in the classical Karst (Kras) aquifer: Evidence from frequent chemical and stable isotope sampling. Acta Carsologica 2008, 37, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Kok, C.J.; Meesters, H.W.G.; Kempers, A.J. Decomposition rate, chemical composition and nutrient recycling of Nymphaea alba L. floating leaf blade detritus as influenced by pH, alkalinity and aluminum in laboratory experiments. Aquat. Bot. 1990, 37, 215–227. [Google Scholar] [CrossRef]
- Canhoto, C.; Graça, M.A.S. Decomposition of Eucalyptus globulus leaves and three native leaf species (Alnus glutinosa, Castanea sativa and Quercus faginea) in a Portuguese low order stream. Hydrobiologia 1996, 333, 79–85. [Google Scholar] [CrossRef]
- Reice, S.R.; Herbst, G. The role of salinity in decomposition of leaves of Phragmites australis in desert streams. J. Arid Environ. 1982, 5, 361–368. [Google Scholar] [CrossRef]
- Sangiorgio, F.; Basset, A.; Pinna, M.; Sabetta, L.; Abbiati, M.; Ponti, M.; Minocci, M.; Orfanidis, S.; Nicolaiou, A.; Moncheva, S.; et al. Environmental factors affecting Phragmites australis litter decomposition in Mediterranean and Black Sea transitional waters. Aquat. Conserv. Marine Freshw. Ecosyst. 2008, 18, 16–26. [Google Scholar] [CrossRef]
- Quintino, V.; Sangiorgio, F.; Ricardo, F.; Mamede, R.; Pires, A.; Freitas, R.; Rodrigues, A.M.; Basset, A. In situ experimental study of reed leaf decomposition along a full salinity gradient. Estuar. Coast. Shelf Sci. 2009, 85, 497–506. [Google Scholar] [CrossRef]
- Thompson, P.L.; Bärlocher, F. Effect of pH on leaf breakdown in streams and in the laboratory. J. N. Am. Benthol. Soc. 1989, 8, 203–210. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. System. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Sangiorgio, F.; Dragan, S.; Rosati, I.; Teodorof, L.; Staras, M.; Georgescu, L.; Basset, A. Decomposition of reed swamp detritus in the Danube Delta: A case study of four eutrophic systems. Trans. Waters Bull. 2008, 2, 26–37. [Google Scholar]
- Woodward, G.; Gessner, M.O.; Giller, P.S.; Gulis, V.; Hladyz, S.; Lecerf, A.; Malmqvist, B.; McKie, B.G.; Tiegs, S.D.; Cariss, H.; et al. Continental-scale effects of nutrient pollution on stream ecosystem functioning. Science 2012, 336, 1438–1440. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.; Castagneyrol, B.; Koricheva, J.; Gulis, V.; Chauvet, E.; Graça, M.A.S. A meta-analysis of the effects of nutrient enrichment on litter decomposition in streams. Biol. Rev. 2015, 90, 669–688. [Google Scholar] [CrossRef]
- Murphy, K.L.; Klopatek, J.M.; Klopatek, C.C. The effects of litter quality and climate on decomposition along an elevational gradient. Ecol. Appl. 1998, 8, 1061–1071. [Google Scholar] [CrossRef]
- Denward, C.M.T.; Tranvik, L.J. Effects of solar radiation on aquatic macrophyte litter decomposition. Oikos 1998, 82, 51–58. [Google Scholar] [CrossRef]
- Kuehn, K.A.; Lemke, M.J.; Suberkropp, K.; Wetzel, R.G. Microbial biomass and production associated with decaying leaf litter of the emergent macrophyte Juncus effusus. Limnol. Ocean. 2000, 45, 862–870. [Google Scholar] [CrossRef]
- Jackson, C.R.; Vallaire, S.C. Microbial activity and decomposition of fine particulate organic matter in a Louisiana cypress swamp. J. N. Am. Benthol Soc. 2007, 26, 743–753. [Google Scholar] [CrossRef]
- Gessner, M.O.; Chauvet, E. Importance of stream microfungi in controlling breakdown rates of leaf litter. Ecology 1994, 75, 1807–1817. [Google Scholar] [CrossRef]
- Findlay, S.; Dye, S.; Kuehn, K. Microbial growth and nitrogen retention in litter of Phragmites australis compared to Typha angustifolia. Wetlands 2002, 22, 616–625. [Google Scholar] [CrossRef]
- van Dokkum, H.P.; Slijkerman, D.M.E.; Rossi, L.; Costantini, M.L. Variation in the decomposition of Phragmites australis litter in a monomictic lake: The role of gammarids. Hydrobiologia 2002, 482, 69–77. [Google Scholar] [CrossRef]
- Bedford, A.P. Decomposition of Phragmites australis litter in seasonally flooded and exposed areas of a managed reedbed. Wetlands 2005, 25, 713–720. [Google Scholar] [CrossRef]
- Karádi-Kovács, K.; Selmeczy, G.B.; Padisák, J.; Schmera, D. Food, substrate or both? Decomposition of reed leaves (Phragmites australis) by aquatic macroinvertebrates in a large shallow lake (Lake Balaton, Hungary). Ann. Limnol. Int. J. Limnol. 2015, 51, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Hieber, M.; Gessner, M.O. Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates. Ecology 2002, 83, 1026–1038. [Google Scholar] [CrossRef]
- Cornut, J.; Elger, A.; Lambrigot, D.; Marmonier, P.; Chauvet, E. Early stages of leaf decomposition are mediated by aquatic fungi in the hyporheic zone of woodland streams. Freshw. Biol. 2010, 55, 2541–2556. [Google Scholar] [CrossRef]
- Raposeiro, P.M.; Ferreira, V.; Guri, R.; Gonçalves, V.; Martins, G.M. Leaf litter decomposition on insular lentic systems: Effects of macroinvertebrate presence, leaf species, and environmental conditions. Hydrobiologia 2016, 784, 69–79. [Google Scholar] [CrossRef]
- Brooks, R.T. Potential impacts of global climate change on the hydrology and ecology of ephemeral freshwater systems of the forests of the northeastern United States. Clim. Chang. 2009, 95, 469–483. [Google Scholar] [CrossRef]
- Gómez-Rodríguez, C.; Bustamante, J.; Díaz-Paniagua, C. Evidence of hydroperiod shortening in a preserved system of temporary ponds. Remote Sens. 2010, 2, 1439–1462. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.T.; Cartwright, J.M.; Collins, G.H.; Long, R.A.; Eitel, J.H. Legacy effects of hydrologic alteration in playa wetland responses to droughts. Wetlands 2020, 40, 2011–2024. [Google Scholar] [CrossRef]
- Cartwright, J.M.; Wolfe, W.J. Increasing Hydroperiod in a Karst-depression Wetland Based on 165 Years of Simulated Daily Water Levels. Wetlands 2021, 41, 75. [Google Scholar] [CrossRef]
- Cucchi, F.; Finocchiaro, F.; Zini, L. Karst Geosites in NE Italy. In Advances in Research in Karst Media. Environmental Earth Sciences; Andreo, B., Carrasco, F., Durán, J., LaMoreaux, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 393–398. [Google Scholar]
- Cucchi, F.; Finocchiaro, F.; Muscio, G. Geositi del Friuli Venezia Giulia; Dipartimento di Scienze Geologiche, Ambientali e Marine dell’Università degli Studi di Trieste—Regione Autonoma Friuli Venezia Giulia, Servizio Geologico: Udine, Italy, 2009; pp. 310–311. [Google Scholar]
- Cucchi, F.; Finocchiaro, F. Karst Landforms in Friuli Venezia Giulia: From Alpine to Coastal Karst. In Landscapes and Landforms of Italy; World Geomorphological Landscapes; Soldati, M., Marchetti, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 147–156. [Google Scholar]
- Calligaris, C.; Mezga, K.; Slejko, F.F.; Urbanc, J.; Zini, L. Groundwater characterization by means of conservative (d18O and d2H) and non-conservative (87Sr/86Sr) isotopic values: The classical Karst region aquifer case (Italy–Slovenia). Geoscience 2018, 8, 321. [Google Scholar] [CrossRef] [Green Version]
- Bertoli, M.; Pizzul, E.; Devescovi, V.; Franz, F.; Pastorino, P.; Giulianini, P.G.; Ferrari, C.; Nonnis Marzano, F. Biology and distribution of Danube barbel (Barbus balcanicus) (Osteichthyes: Cyprinidae) at the Northwestern limit of its range. Eur. Zool. J. 2019, 86, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Petersen, R.C.; Cummins, K.W. Leaf processing in a woodland stream. Freshw. Biol. 1974, 4, 343–368. [Google Scholar] [CrossRef]
- Basset, A.; Sangiorgio, F.; Sabetta, L. Handbook for the Application of Body Size Descriptors to Monitoring Safety of Transitional Ecosystems; TW ReferenceNet—EU INTERREG III B Project 3B073; Management and sustainable development of protected transitional waters; University of Lecce: Lecce, Italy, 2006. [Google Scholar]
- Bertoli, M.; Brichese, G.; Michielin, D.; Ruzič, M.; Vignes, F.; Basset, A.; Pizzul, E. Seasonal and multi-annual patterns of Phragmites australis decomposition in a wetland of the Adriatic area (Northeast Italy): A three-years analysis. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 14. [Google Scholar] [CrossRef] [Green Version]
- Bertoli, M.; Brichese, G.; Pastorino, P.; Prearo, M.; Vignes, F.; Basset, A.; Pizzul, E. Seasonal multi-annual trends in energy densities of the midges (genus Chironomus) in a Mediterranean temporary wetland (Natural Regional Reserve of the Isonzo River Mouth, Northeast Italy). Hydrobiologia 2018, 823, 153–167. [Google Scholar] [CrossRef]
- Bärlocher, F. Pitfalls of Traditional Techniques When Studying Decomposition of Vascular Plant Remains in Aquatic Habitats. Limnetica 1997, 13, 1–11. [Google Scholar] [CrossRef]
- Merritt, R.W.; Cummins, K.W. Trophic relationships of macroinvertebrates. In Methods in Stream Ecology; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: San Diego, CA, USA, 2006; pp. 585–610. [Google Scholar]
- Mauricio Zambrano-Bigiarini. hydroTSM: Time Series Management, Analysis and Interpolation for Hydrological Modelling. R Package Version 0.6-0. 2020. Available online: https://cran.r-project.org/web/packages/hydroTSM/hydroTSM.pdf (accessed on 25 May 2022).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd., PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall International Inc.: Hoboken, NJ, USA, 1984. [Google Scholar]
- Bärlocher, F. Leaf mass loss estimated by litter bag technique. In Methods to Study Litter Decomposition: A Practical Guide; Graça, M.A.S., Bärlocher, F., Gessner, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 37–42. [Google Scholar]
- Grömping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 April 2022).
- RStudio Team. RStudio: Integrated Development Environment for R; R Studio, PBC: Boston, MA, USA, 2021; Available online: http://www.rstudio.com/ (accessed on 20 April 2022).
- Friuli Venezia Giulia Regional Authority. Studio Conoscitivo dei Cambiamenti Climatici e di Alcuni loro Impatti in Friuli Venezia Giulia. Regione Autonoma Friuli Venezia Giulia—Assessorato All’ambiente ed Energia. 2018. Available online: https://www.meteo.fvg.it/clima/clima_fvg/03_cambiamenti_climatici/01_REPORT_cambiamenti_climatici_e_impatti_per_il_FVG/impattiCCinFVG_marzo2018.pdf (accessed on 24 April 2022).
- Boero, F.; Belmonte, G.; Bussotti, S.; Fanelli, G.; Fraschetti, S.; Giangrande, A.; Gravili, C.; Guidetti, P.; Pati, A.; Piraino, S.; et al. From biodiversity and ecosystem functioning to the roots of ecological complexity. Ecol. Complex. 2004, 1, 101–109. [Google Scholar]
- Dvořak, J.; Best, E.P.H. Macroinvertebrate communities associated with macrophytes of Lake Vechten: Structural and functional relationships. Hydrobiologia 1982, 95, 115–126. [Google Scholar] [CrossRef]
- Diehl, S.; Kornijòw, R. Influence of submerged macrophytes on trophic interactions among fish and macroinvertebrates. In The Structuring Role of Submerged Macrophytes in Lakes; Jeppesen, E., Søndergaard, M., Søndergaard, M., Christoffersen, K., Eds.; Springer: New York, NY, USA, 1998; pp. 24–46. [Google Scholar]
- Heino, J. Lentic macroinvertebrate assemblage structure along gradients in spatial heterogeneity, habitat size and water chemistry. Hydrobiologia 2000, 418, 229–242. [Google Scholar] [CrossRef]
- Zbikowski, J.; Kobak, J. Factors influencing taxonomic composition and abundance of macrozoobenthos in extralittoral zone of shallow eutrophic lakes. Hydrobiologia 2007, 584, 145–155. [Google Scholar] [CrossRef]
- Jayawardana, J.M.C.K.; Westbrooke, M.; Wilson, M.; Hurst, C. Macroinvertebrate communities in Phragmites australis (Cav.) Trin. ex Steud. reed beds and open bank habitats in central Victorian streams in Australia. Hydrobiologia 2006, 568, 169–185. [Google Scholar] [CrossRef]
- Sychra, J.; Adàmek, Z.; Petřivalskà, K. Distribution and diversity of littoral macroinvertebrates within extensive reed beds of a lowland pond. Int. J. Limnol. 2010, 46, 281–289. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; Al-Sodany, Y.M. Decomposition dynamics of Phragmites australis litter in Lake Burullus, Egypt. Plant Spec. Biol. 2014, 29, 47–56. [Google Scholar] [CrossRef]
- Economic Commission for Europe. Assessment of the water-food-energy-ecosystems nexus in the Isonzo/Soča River Basin. In Proceedings of the Meeting of the Parties to the Convention on the Protection and Use of Transboundary Watercourses and International Lakes, Budapest, Hungary, 17–19 November 2015; Economic and Social Council: Budapest, Hungary, 2015. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude | Longitude |
|---|---|---|
| 1 | 45°49′59.05″ N | 13°33′15.05″ E |
| 2 | 45°49′55.84″ N | 13°33′27.69″ E |
| 3 | 45°49′47.87″ N | 13°33′40.79″ E |
| 4 | 45°49′46.72″ N | 13°33′52.40″ E |
| 5 | 45°49′43.03″ N | 13°33′57.92″ E |
| Seasonal Comparison (Period 2021–2022) | |||||
| Source | Sum of squares | d.f. | Mean squares | F | p |
| Season | 1.518 | 3 | 0.506 | 3.290 | 0.001 |
| Residual | 8.612 | 56 | 0.153 | ||
| Total | 10.131 | 59 | |||
| Autumn 2016–2017 vs. Autumn 2021–2022 | |||||
| Source | Sum of squares | d.f. | Mean squares | F | p |
| Season | 0.745 | 1 | 0.745 | 4.915 | 0.001 |
| Residual | 4.245 | 28 | 0.151 | ||
| Total | 4.990 | 29 | |||
| Spring 2016–2017 vs. Spring 2021–2022 | |||||
| Source | Sum of squares | d.f. | Mean squares | F | p |
| Season | 0.484 | 1 | 0.484 | 3.701 | 0.001 |
| Residual | 3.532 | 28 | 0.131 | ||
| Total | 4.017 | 29 | |||
| Year 2021–2022, Seasonal Comparisons | 2016–2017 vs. 2021–2022—Autumn | 2016–2017 vs. 2021–2022—Spring | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxon | Av. Dissim | Contrib. % | Cumul. % | Taxon | Av. Dissim | Contrib. % | Cumul. % | Taxon | Av. Dissim | Contrib. % | Cumul. % |
| Asellus | 4.79 | 8.38 | 8.38 | Asellus | 5.475 | 9.108 | 9.108 | Asellus | 6.197 | 11.390 | 11.390 |
| Gammarus | 4.60 | 8.04 | 16.41 | Gammarus | 4.835 | 8.043 | 17.150 | Gammarus | 5.625 | 10.340 | 21.720 |
| Palpomyia | 4.12 | 7.19 | 23.61 | Palpomyia | 4.601 | 7.653 | 24.800 | Chironominae | 5.225 | 9.601 | 31.330 |
| Hydracarina | 3.41 | 5.96 | 29.56 | Erpobdella | 4.226 | 7.030 | 31.830 | Palpomyia | 3.852 | 7.078 | 38.400 |
| Chironominae | 3.17 | 5.53 | 35.10 | Tanypodinae | 3.109 | 5.173 | 37.010 | Tubificinae | 3.703 | 6.804 | 45.210 |
| Limnodrilus | 2.91 | 5.08 | 40.18 | Orthocladiinae | 2.773 | 4.613 | 41.620 | Erpobdella | 3.497 | 6.425 | 51.630 |
| Erpobdella | 2.50 | 4.36 | 44.54 | Tubificinae | 2.711 | 4.509 | 46.130 | Leptoceridae | 2.600 | 4.777 | 56.410 |
| Polycelis | 2.32 | 4.05 | 48.59 | Glossiphonia | 2.554 | 4.249 | 50.380 | Sphaerium | 2.271 | 4.174 | 60.580 |
| Dugesia | 2.06 | 3.60 | 52.19 | Dugesia | 2.417 | 4.022 | 54.400 | Glossiphonia | 2.180 | 4.005 | 64.590 |
| Dendrocoelum | 1.92 | 3.36 | 55.55 | Dendrocoelum | 2.239 | 3.725 | 58.130 | Orthocladiinae | 1.914 | 3.516 | 68.100 |
| Leptoceridae | 1.86 | 3.24 | 58.80 | Haliplus | 2.225 | 3.701 | 61.830 | Bithynia | 1.785 | 3.280 | 71.380 |
| Orthocladiinae | 1.84 | 3.21 | 62.00 | Polycelis | 1.816 | 3.022 | 64.850 | Polycelis | 1.729 | 3.177 | 74.560 |
| Tubifex | 1.78 | 3.10 | 65.11 | Dugesia | 1.683 | 3.093 | 77.650 | ||||
| Bithynia | 1.77 | 3.09 | 68.20 | ||||||||
| β | St. Error β | Estimate | St. Error | t Value | p-Level | Relative Importance (%) | |
|---|---|---|---|---|---|---|---|
| Intercept | 0.000 | 0.013 | −0.087 | 0.013 | −6.683 | 0.000 | |
| Temperature | 0.492 | 0.002 | 0.015 | 0.002 | 7.539 | 0.000 | 47.56 |
| Shredders | 0.320 | 0.000 | 0.001 | 0.000 | 5.846 | 0.000 | 22.56 |
| Soča | −0.348 | 0.005 | 0.007 | 0.005 | 6.097 | 0.000 | 22.42 |
| DO | 0.185 | 0.001 | 0.005 | 0.001 | 3.024 | 0.003 | 5.79 |
| Rainfall | −0.080 | 0.001 | 0.002 | 0.001 | 1.527 | 0.047 | 1.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertoli, M.; Lesa, D.; Merson, A.; Pastorino, P.; Prearo, M.; Pizzul, E. Biodiversity and Litter Breakdown in a Karstic Freshwater System (Doberdò Lake, Northeast Italy) in Relation to Water Level Fluctuations and Environmental Features. Diversity 2022, 14, 460. https://doi.org/10.3390/d14060460
Bertoli M, Lesa D, Merson A, Pastorino P, Prearo M, Pizzul E. Biodiversity and Litter Breakdown in a Karstic Freshwater System (Doberdò Lake, Northeast Italy) in Relation to Water Level Fluctuations and Environmental Features. Diversity. 2022; 14(6):460. https://doi.org/10.3390/d14060460
Chicago/Turabian StyleBertoli, Marco, Davide Lesa, Alessia Merson, Paolo Pastorino, Marino Prearo, and Elisabetta Pizzul. 2022. "Biodiversity and Litter Breakdown in a Karstic Freshwater System (Doberdò Lake, Northeast Italy) in Relation to Water Level Fluctuations and Environmental Features" Diversity 14, no. 6: 460. https://doi.org/10.3390/d14060460
APA StyleBertoli, M., Lesa, D., Merson, A., Pastorino, P., Prearo, M., & Pizzul, E. (2022). Biodiversity and Litter Breakdown in a Karstic Freshwater System (Doberdò Lake, Northeast Italy) in Relation to Water Level Fluctuations and Environmental Features. Diversity, 14(6), 460. https://doi.org/10.3390/d14060460