
A Study of Phenolic Compounds and Their Chemophenetic Value in the Genus Thesium (Santalaceae)

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Used
2.2. Extraction
2.3. Standards
2.4. Liquid Chromatography–Mass Spectrometry (LCMS)
2.5. Compound Identification
2.6. Multivariate Analysis
2.7. Phylogenetic Comparison
3. Results
3.1. Overview of Phenolic Compounds
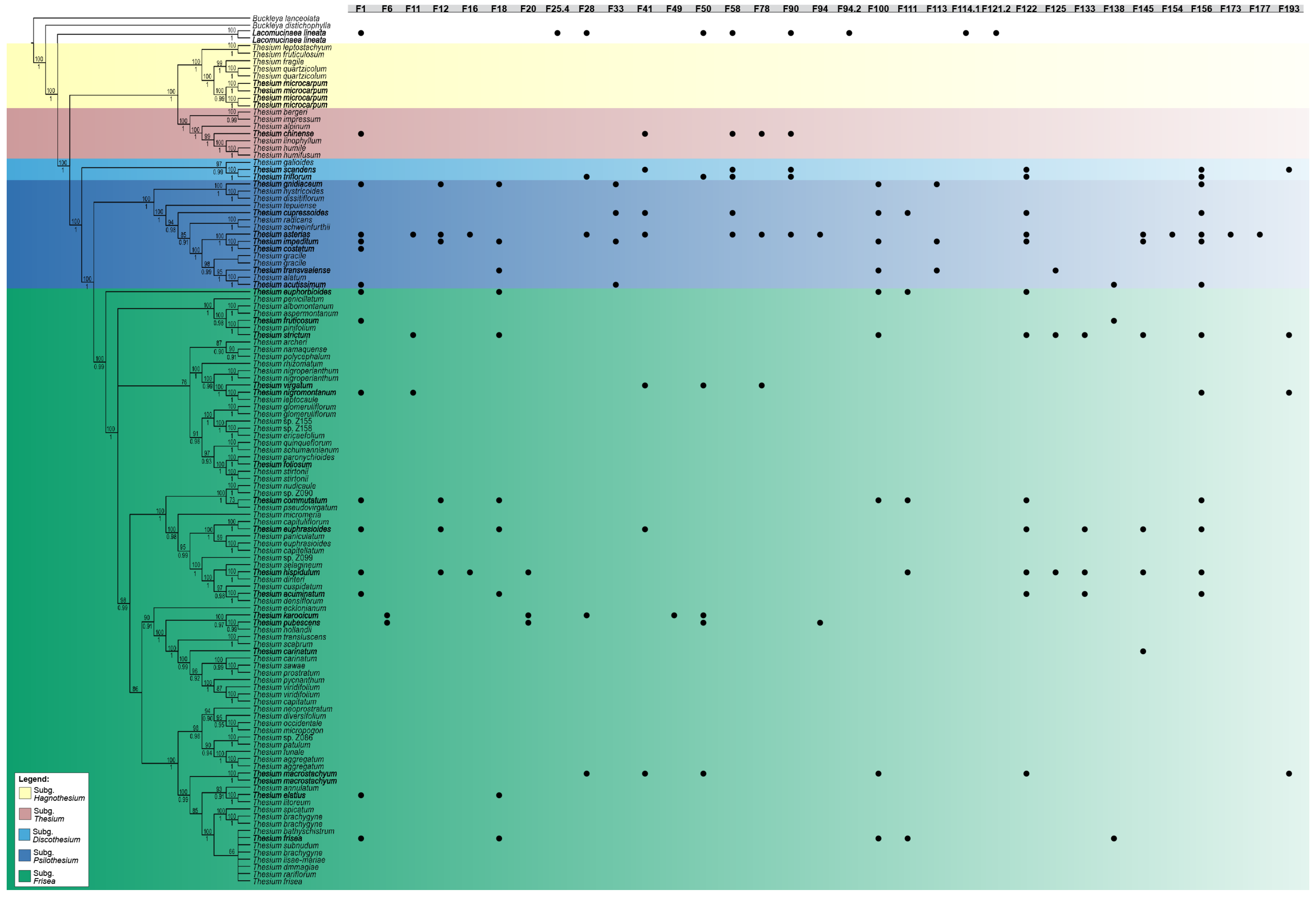
3.2. Chemophenetic Value at Generic Level
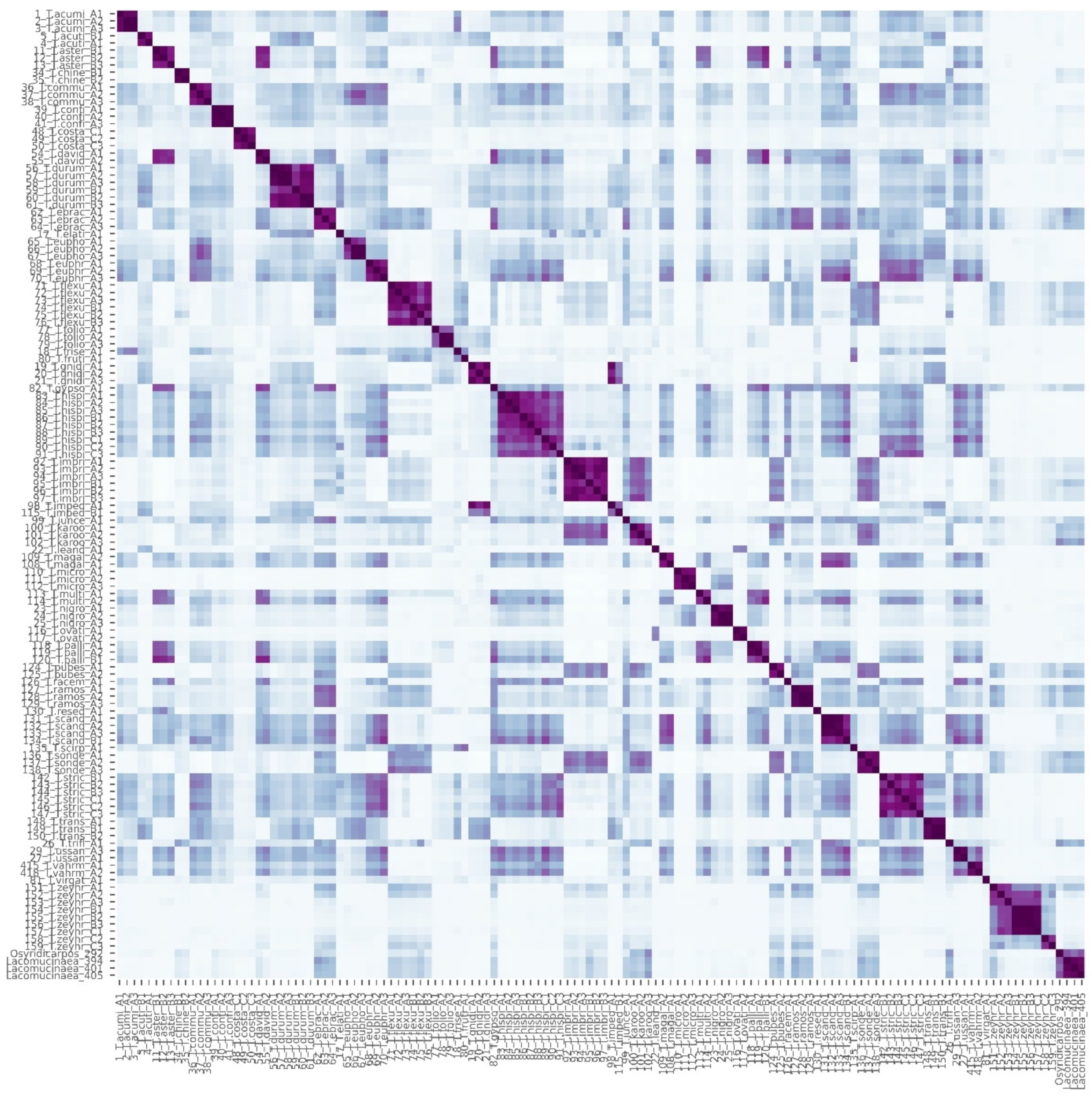
3.3. Chemophenetic Value at Infrageneric Level
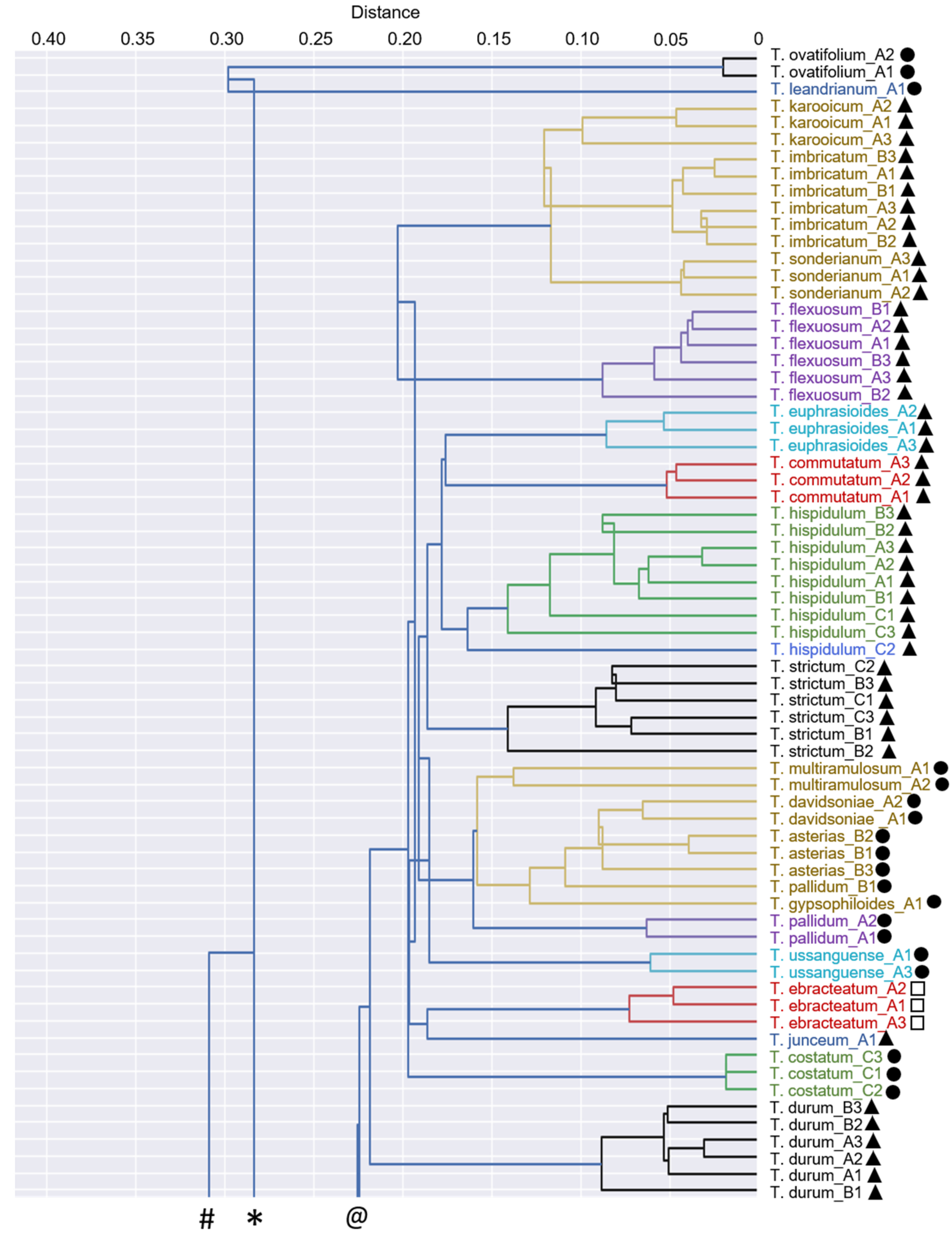
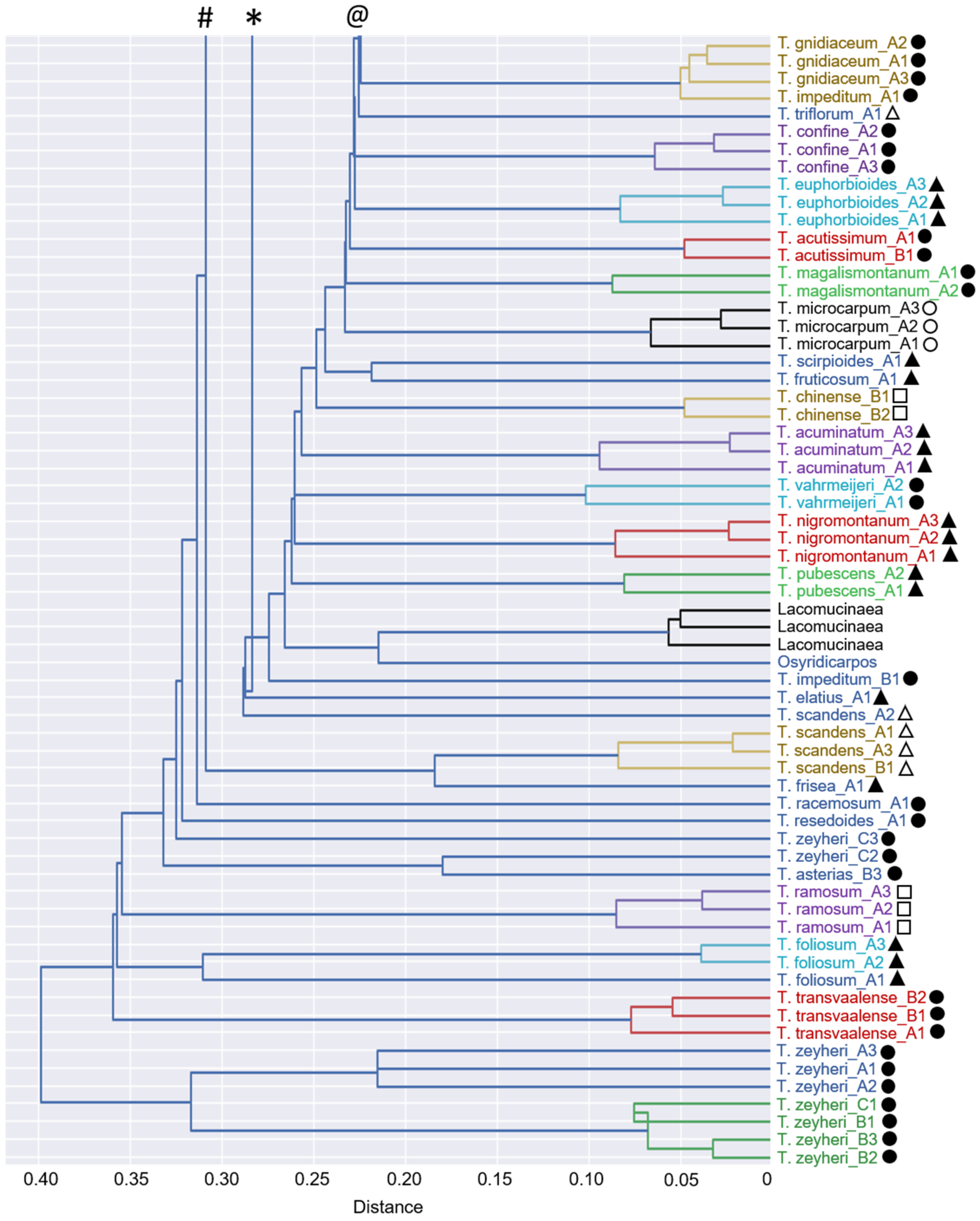
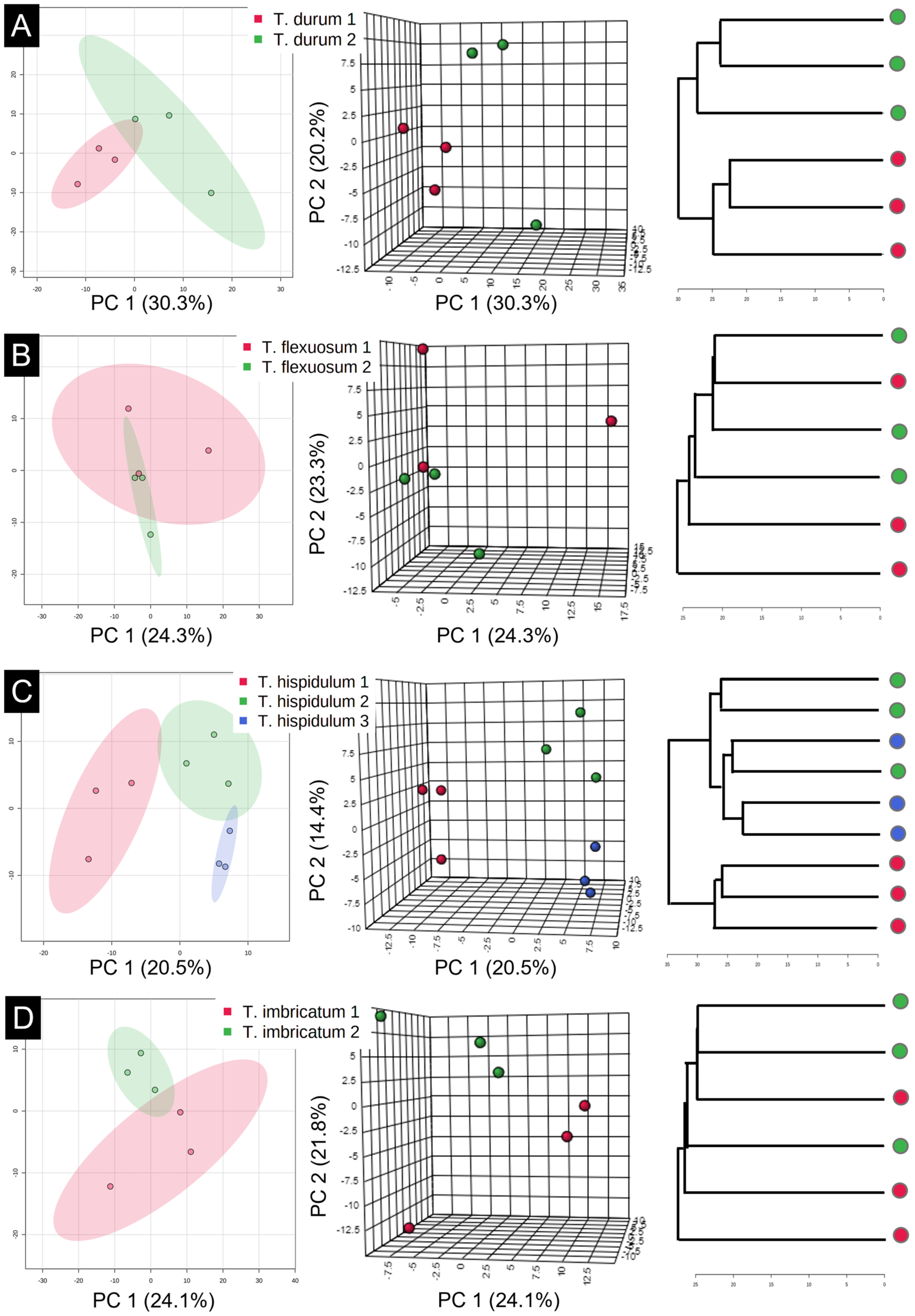
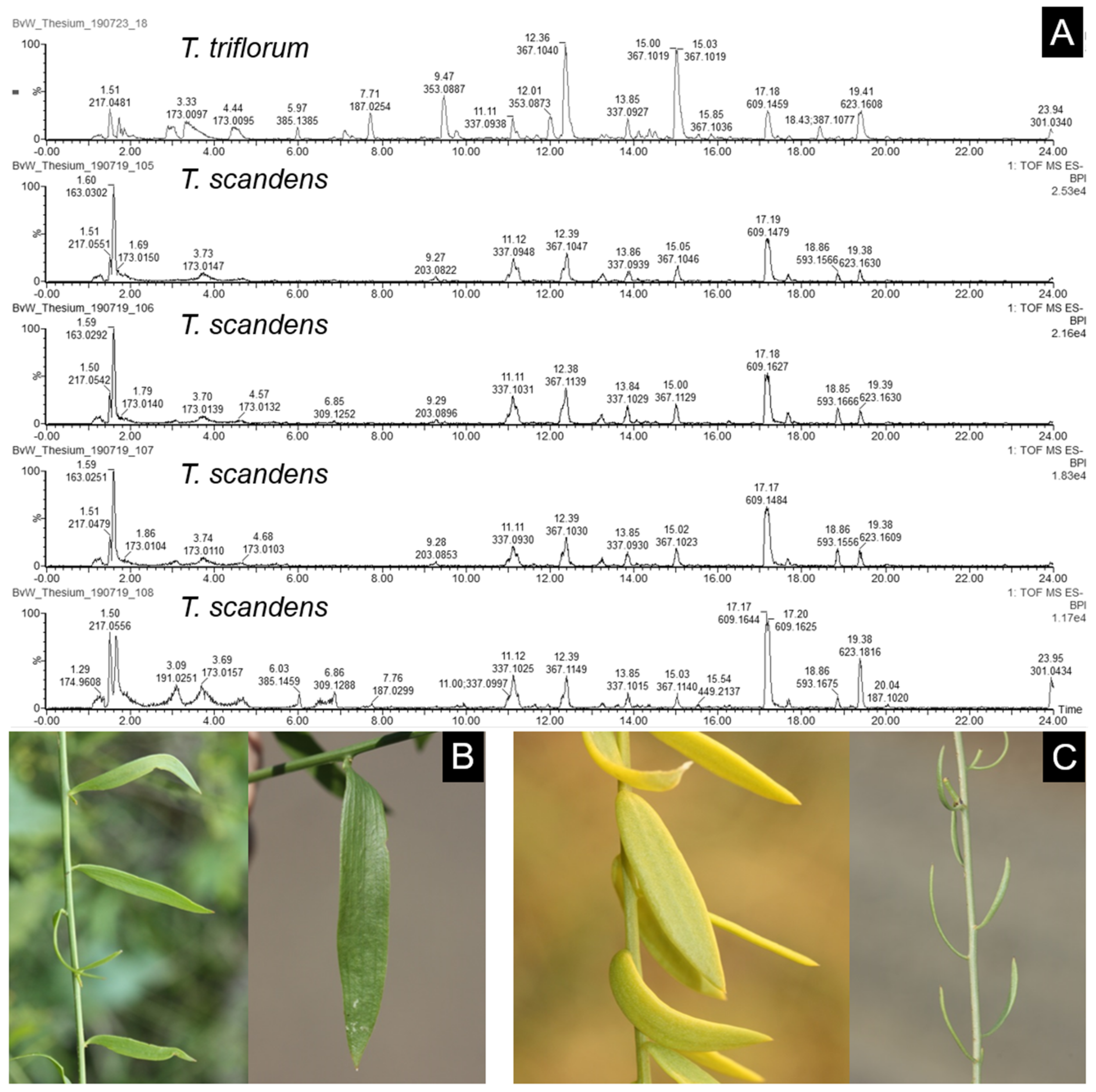
3.4. Chemophenetic Value at Interspecific Level
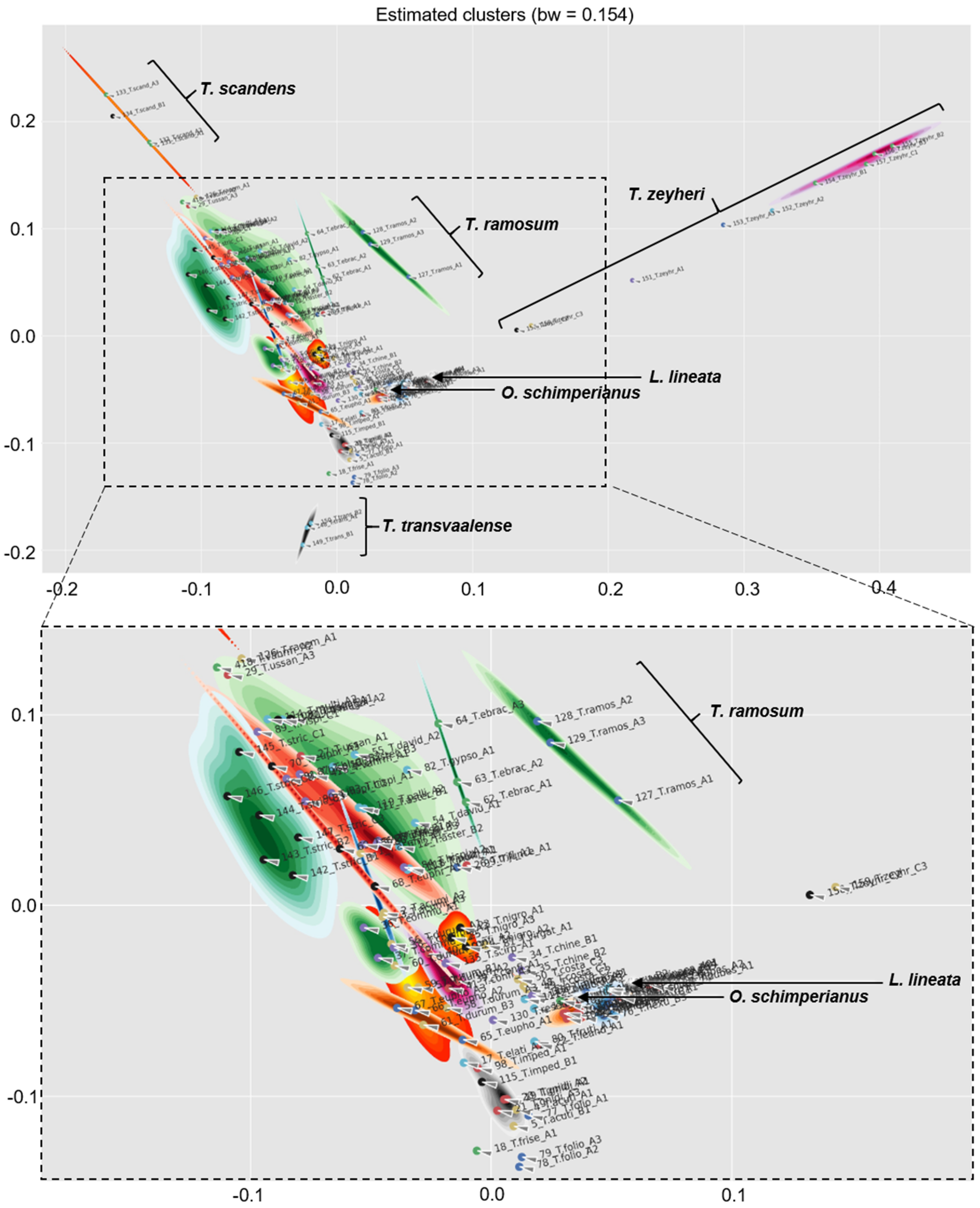
3.5. Chemophenetic Value at Infraspecific Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nickrent, D.L.; García, M.A. Lacomucinaea, a new monotypic genus in Thesiaceae (Santalales). Phytotaxa 2015, 224, 173–184. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lombard, N.; Van Wyk, B.-E.; le Roux, M.M. A review of the ethnobotany, contemporary uses, chemistry and pharmacology of the genus Thesium (Santalaceae). J. Ethnopharmacol. 2020, 256, 112745. [Google Scholar] [CrossRef] [PubMed]
- García, M.A.; Nickrent, D.L.; Mucina, L. Thesium nautimontanum, a new species of Thesiaceae (Santalales) from South Africa. PhytoKeys 2018, 109, 41–51. [Google Scholar] [CrossRef]
- De Candolle, A. Santalaceae. In Prodromus Systematis Naturalis; De Candolle, A., Ed.; Treuttel et Würtz: Paris, France, 1857; pp. 619–692. [Google Scholar]
- Hill, A.W. Order CXX. Santalaceae. In Flora Capensis, Volume 5; Thiselton-Dyer, W.T., Ed.; L. Reeve & Co. Ltd.: London, UK, 1925; pp. 135–212. [Google Scholar] [CrossRef]
- Zhigila, D.A.; Verboom, G.A.; Muasya, A.M. An infrageneric classification of Thesium (Santalaceae) based on molecular phylogenetic data. Taxon 2020, 69, 100–123. [Google Scholar] [CrossRef]
- Linnaeus, C. Species Plantarum: Exhibentes Plantas Rite Cognitas, Ad Genera Relatas, Cum Differentiis Specificis, Nominibus Trivialibus, Synonymis Selectis, Locis Natalibus, Secundum Systema Sexuale Digestas; Tomus I; Laurentius Salvius: Stockholm, Sweden, 1753; pp. 207–208. [Google Scholar]
- Hill, A.W. The genus Thesium in South Africa, with a key and descriptions of new species. Bull. Misc. Inf. 1915, 1, 1–43. [Google Scholar] [CrossRef]
- Pilger, R. Santalaceae. In Die Natürlichen Pflanzen Familien, 2nd ed.; Engler, A., Ed.; W. Engelmann: Leipzig, Germany, 1935; pp. 52–91. [Google Scholar]
- Bobrov, E.G. Family XLIX. Santalaceae R.Br. In 5. Flora of the U.S.S.R; Komarov, V.L., Ed.; Israel Program for Scientific Translations: Jerusalem, Israel, 1970; pp. 325–340. [Google Scholar]
- Hendrych, R. The natural history and systematic of the genus Thesium L. Acta Univ. Carol. Biol. 1970, 4, 293–358. [Google Scholar]
- Stander, M.A.; Brendler, T.; Redelinghuys, H.; Van Wyk, B.-E. The commercial history of Cape herbal teas and the analysis of phenolic compounds in historic teas from a depository of 1933. J. Food Compos. Anal. 2019, 76, 66–73. [Google Scholar] [CrossRef]
- De Kock, P.C.; Rapson, W.S. A correlation between the chemical constituents and morphological structure of certain Thesium species. Nature 1938, 142, 1078–1079. [Google Scholar] [CrossRef]
- Moore, T.E.; Verboom, G.A.; Forest, F. Phylogenetics and biogeography of the parasitic genus Thesium L. (Santalaceae), with an emphasis on the Cape of South Africa. Bot. J. Linn. Soc. 2010, 162, 435–452. [Google Scholar] [CrossRef] [Green Version]
- Willems, J.L.; Khamis, M.M.; Mohammed-Saeid, W.; Purves, R.; Katselis, G.; Low, N.H.; El-Aneed, A. Analysis of a series of chlorogenic acid isomers using differential ion mobility and tandem mass spectrometry. Anal. Chim. Acta 2016, 933, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Assanga, S.B.I.; Luján, L.M.L.; Ruiz, J.C.G.; McCarty, M.F.; Cota-Arce, J.M.; Espinoza, C.L.L.; Gil Salido, A.A.; Ángulo, D.F. Comparative analysis of phenolic content and antioxidant power between parasitic Phoradendron californicum (toji) and their hosts from Sonoran Desert. Results Chem. 2020, 2, 100079. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, J. Study on chemical constituents of Thesium chinense. Chin. Tradit. Herb. Drugs 2004, 35, 491–493. [Google Scholar]
- Parveen, Z.; Deng, Y.; Saeed, M.K.; Dai, R.; Ahamad, W.; Yu, Y.H. Anti-inflammatory and analgesic activities of Thesium chinense Turcz. extracts and its major flavonoids, kaempferol and kaempferol-3-O-glucoside. Yakugaku Zasshi 2007, 127, 1275–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.K.; Kim, K.H.; Choi, S.U.; Lee, J.H.; Lee, K.R. Phytochemical constituents of Thesium chinense Turcz. and their cytotoxic activities in vitro. Nat. Prod. Sci. 2009, 15, 246–249. [Google Scholar]
- Liu, Y.; Zahida, P.; Deng, Y.L.; Zhang, J.H.; Dai, R.J. Study on the flavonoids compounds of Thesium chinense. Zhong Yao Cai 2009, 32, 518–520. [Google Scholar]
- Liu, C.; Li, X.-T.; Cheng, R.-R.; Han, Z.-Z.; Yang, L.; Song, Z.-C.; Wang, Z.-T. Anti-oral common pathogenic bacterial active acetylenic acids from Thesium chinense Turcz. J. Nat. Med. 2018, 72, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Hong, M.; Yang, X. Isolation of chemical components from Thesium chinense. Zhongguo Shiyan Fangjixue Zazhi 2016, 22, 74–77. [Google Scholar]
- Forest, F.; Manning, J.C. The minor genera Kunkeliella and Thesidium included in Thesium. Bothalia 2013, 43, 214–216. [Google Scholar] [CrossRef] [Green Version]
- Kuttan, R.; Pattabhiraman, K.S.V.; Radhakrishnan, A.N. Possible chemotaxonomic significance of the occurrence of cis-4-hydroxy-L-proline in Santalaceae. Phytochemistry 1974, 13, 453–454. [Google Scholar] [CrossRef]
- Hopkins, C.Y.; Chisholm, M.J.; Cody, W.J. Fatty acid components of some Santalaceae seed oils. Phytochemistry 1969, 8, 161–165. [Google Scholar] [CrossRef]
- Hilliard, O.M. Thesium. In Flora Zambesiaca, Vol. 9, Part 3: Polygonaceae and Myriaceae; Pope, G.V., Polhill, R.M., Martins, E.S., Eds.; Royal Botanic Gardens, Kew: Richmond, UK, 2006; pp. 206–241. [Google Scholar]
- Jadhav, R.B.; Anarthe, S.J.; Surana, S.J.; Gokhale, S.B. Host-hemiparasite transfer of the C-glucosyl xanthone mangiferin between Mangifera indica and Dendrophthoe falcate. J. Plant Interact. 2005, 1, 171–177. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Species | Subgenus | Sample Code | Collection Details | |||
|---|---|---|---|---|---|---|---|
| Country | Locality | Date | Voucher | ||||
| 1 | Thesium acuminatum A.W.Hill | Frisea (Rchb.) Hendrych | 1_T.acumi_A1 * | South Africa | Western Cape, Silvermine | 2018/09/13 | Visser and van Wyk 305 |
| 2 | 2_T.acumi_A2 * | ||||||
| 3 | 3_T.acumi_A3 * | ||||||
| 4 | Thesium acutissimum A.DC. | Psilothesium (A.DC.) Zhigila, Verboom and Muasya | 4_T.acuti_A1 * | South Africa | Eastern Cape, Grahamstown area | 2018/11/04 | Visser et al. 369 |
| 5 | 5_T.acuti_B1 * | South Africa | Eastern Cape, Grahamstown | 2018/11/04 | Visser et al. 370 | ||
| 6 | Thesium asterias A.W.Hill | Psilothesium | 6_T.aster_A1 | South Africa | KwaZulu-Natal, Cape Vidal | 2018/02/02 | van Wyk s.n. |
| 7 | 7_T.aster_A2 | ||||||
| 8 | 8_T.aster_A3 | ||||||
| 9 | 9_T.aster_A4 | ||||||
| 10 | 10_T.aster_A5 | ||||||
| 11 | 11_T.aster_B1 * | South Africa | Limpopo, Lekgalameetse Nature Reserve | 2017/12/05 | Visser and le Roux 291 | ||
| 12 | 12_T.aster_B2 * | ||||||
| 13 | 13_T.aster_B3 * | ||||||
| 14 | Thesium carinatum A.DC. | Frisea | 14_T.carin_A1 | South Africa | Western Cape | 2017 | van Wyk s.n. |
| 15 | 15_T.carin_A2 | ||||||
| 16 | 16_T.carin_A3 | ||||||
| 17 | Thesium chinense Turcz. var. longipedunculatum Chu | Thesium | 31_T.chine_A1 | China | - | 2004/05/04 | Guo 1506034 |
| 18 | 32_T.chine_A2 | ||||||
| 19 | Thesium chinense Turcz. var. chinense | Thesium | 34_T.chine_B1 * | Japan | Honshu, Tsukuba City | 1997/05/07 | Konta and Momose 18144 |
| 20 | 35_T.chine_B2 * | ||||||
| 21 | Thesium commutatum Sond. | Frisea | 36_T.commu_A1 * | South Africa | Western Cape, Silvermine | 2018/09/13 | Visser and van Wyk 304 |
| 22 | 37_T.commu_A2 * | ||||||
| 23 | 38_T.commu_A3 * | ||||||
| 24 | Thesium confine Sond. | Psilothesium | 39_T.confi_A1 * | South Africa | Eastern Cape, Middelburg area | 2016/10/26 | Visser and le Roux 208 |
| 25 | 40_T.confi_A2 * | ||||||
| 26 | 41_T.confi_A3 * | ||||||
| 27 | Thesium costatum A.W.Hill | Psilothesium | 42_T.costa_A1 | South Africa | Gauteng, Pretoria National Botanical Garden | 2016/10/18 | le Roux 184 |
| 28 | 43_T.costa_A2 | ||||||
| 29 | 44_T.costa_A3 | ||||||
| 30 | 48_T.costa_C1 * | South Africa | Limpopo, Haenertsburg | 2017/12/05 | Visser and le Roux 293 | ||
| 31 | 49_T.costa_C2 * | ||||||
| 32 | 50_T.costa_C3 * | ||||||
| 33 | Thesium cupressoides A.W.Hill | Psilothesium | 51_T.cupre_A1 | South Africa | KwaZulu-Natal, Sani Pass | 2017/01/31 | Visser et al. 252 |
| 34 | 52_T.cupre_B1 | South Africa | KwaZulu-Natal, Umtamvuna Nature Reserve area | 2017/02/01 | Visser et al. 253 | ||
| 35 | 53_T.cupre_C1 | South Africa | KwaZulu-Natal, Umtamvuna Nature Reserve | 2017/02/02 | Visser et al. 262 | ||
| 36 | Thesium cymosum A.W.Hill | Psilothesium | 30_T.cymos_A1 | Mozambique | Manica, Mavita | 1944/10/25 | Mendonça 2565 |
| 37 | Thesium davidsoniae Brenan | Psilothesium | 54_T.david_A1 * | South Africa | Limpopo, Abel Erasmus Pass | 2017/12/04 | Visser and le Roux 289 |
| 38 | 55_T.david_A2 * | ||||||
| 39 | Thesium durum Hilliard and B.L.Burtt | Frisea | 56_T.durum_A1 * | South Africa | Eastern Cape, Lady Grey | 2018/10/30 | Visser et al. 352 |
| 40 | 57_T.durum_A2 * | ||||||
| 41 | 58_T.durum_A3 * | ||||||
| 42 | 59_T.durum_B1 * | South Africa | Eastern Cape, Rossouw-Indwe area | 2018/11/02 | Visser et al. 362 | ||
| 43 | 60_T.durum_B2 * | ||||||
| 44 | 61_T.durum_B3 * | ||||||
| 45 | Thesium ebracteatum Hayne | Thesium | 62_T.ebrac_A1 * | Lithuania | - | 2018/06 | Gudzinskas s.n. |
| 46 | 63_T.ebrac_A2 * | ||||||
| 47 | 64_T.ebrac_A3 * | ||||||
| 48 | Thesium cf. elatius Sond. | Frisea | 17_T.elati_A1 * | South Africa | Western Cape, Betty’s Bay | 2018/09/14 | Visser et al. 312 |
| 49 | Thesium euphorbioides P.J.Bergius | Frisea | 65_T.eupho_A1 * | South Africa | Western Cape, Fernkloof Nature Reserve | 2018/09/14 | Visser et al. 316 |
| 50 | 66_T.eupho_A2 * | ||||||
| 51 | 67_T.eupho_A3 * | ||||||
| 52 | Thesium euphrasioides A.DC. | Frisea | 68_T.euphr_A1 * | South Africa | Western Cape, Franschoek area | 2018/09/12 | Visser and van Wyk 300 |
| 53 | 69_T.euphr_A2 * | ||||||
| 54 | 70_T.euphr_A3 * | ||||||
| 55 | Thesium flexuosum A.DC. | Frisea | 71_T.flexu_A1 * | South Africa | Eastern Cape, Hankey area | 2018/12/30 | Visser 379 |
| 56 | 72_T.flexu_A2 * | ||||||
| 57 | 73_T.flexu_A3 * | ||||||
| 58 | 74_T.flexu_B1 * | South Africa | Eastern Cape, Grahamstown area | 2018/11/04 | Visser et al. 368 | ||
| 59 | 75_T.flexu_B2 * | ||||||
| 60 | 76_T.flexu_B3 * | ||||||
| 61 | Thesium foliosum A.DC. | Frisea | 77_T.folio_A1 * | South Africa | Eastern Cape, Van Stadens Wild Flower Local Nature Reserve | 2018/12/30 | Visser 380 |
| 62 | 78_T.folio_A2 * | ||||||
| 63 | 79_T.folio_A3 * | ||||||
| 64 | Thesium cf. frisea L. | Frisea | 18_T.frise_A1 * | South Africa | Western Cape, Betty’s Bay | 2018/09/14 | Visser et al. 310 |
| 65 | Thesium fruticosum A.W.Hill | Frisea | 80_T.fruti_A1 * | South Africa | Eastern Cape, Grahamstown | 2018/11/04 | Visser et al. 371 |
| 66 | Thesium cf. gnidiaceum A.DC. | Psilothesium | 19_T.gnidi_A1 * | South Africa | Eastern Cape, Elliot area | 2018/11/01 | Visser et al. 360 |
| 67 | 20_T.gnidi_A2 * | ||||||
| 21_T.gnidi_A3 * | |||||||
| 69 | Thesium gypsophiloides A.W.Hill | Psilothesium | 82_T.gypso_A1 * | South Africa | KwaZulu-Natal, Vernon Crookes Nature Reserve | 2017/02/03 | Visser et al. 269 |
| 70 | Thesium hispidulum Lam. | Frisea | 83_T.hispi_A1 * | South Africa | Western Cape, Central Cederberg | 2018/09/17 | Visser and van Wyk 337 |
| 71 | 84_T.hispi_A2 * | ||||||
| 72 | 85_T.hispi_A3 * | ||||||
| 73 | 86_T.hispi_B1 * | South Africa | Western Cape, Piekernieskloof Pass | 2018/09/16 | Visser and van Wyk 329 | ||
| 74 | 87_T.hispi_B2 * | ||||||
| 75 | 88_T.hispi_B3 * | ||||||
| 76 | 89_T.hispi_C1 * | South Africa | Western Cape, Bain’s Kloof Pass | 2018/09/16 | Visser and van Wyk 327 | ||
| 77 | 90_T.hispi_C2 * | ||||||
| 78 | 91_T.hispi_C3 * | ||||||
| 79 | Thesium imbricatum Thunb. | Frisea | 92_T.imbri_A1 * | South Africa | Eastern Cape, Rossouw-Indwe area | 2018/11/02 | Visser et al. 363 |
| 80 | 93_T.imbri_A2 * | ||||||
| 81 | 94_T.imbri_A3 * | ||||||
| 82 | 95_T.imbri_B1 * | South Africa | Eastern Cape, Maclear area | 2018/10/31 | Visser et al. 357 | ||
| 83 | 96_T.imbri_B2 * | ||||||
| 84 | 97_T.imbri_B3 * | ||||||
| 85 | Thesium impeditum A.W.Hill | Psilothesium | 98_T.imped_A1 * | South Africa | Eastern Cape, Hogsback area | 2018/11/03 | Visser et al. 366 |
| 86 | 115_T.imped_B1 * | South Africa | Free State, Frankfort area | 2016/12/12 | Visser and le Roux 233 | ||
| 87 | Thesium junceum Bernh. | Frisea | 99_T.junce_A1 * | South Africa | Eastern Cape, Maclear area | 2018/10/31 | Visser et al. 356 |
| 88 | Thesium karooicum Compton | Frisea | 100_T.karoo_A1 * | South Africa | Western Cape, Montagu | 2018/09/15 | Visser et al. 318 |
| 89 | 101_T.karoo_A2 * | ||||||
| 90 | 102_T.karoo_A3 * | ||||||
| 91 | Thesium cf. leandrianum Cavaco and Keraudren | Psilothesium | 22_T.leand_A1 * | Madagascar | Marojejy National Park | 1989/02/15 | Miller and Lowry 4171 |
| 92 | Thesium magalismontanum Sond. | Psilothesium | 108_T.magal_A1 * | South Africa | Gauteng, Dinokeng Nature Reserve | 2017/12/12 | Nel 482 |
| 93 | 109_T.magal_A2 * | ||||||
| 94 | Thesium microcarpum A.DC. | Hagnothesium (A.DC.) Zhigila, Verboom and Muasya | 110_T.micro_A1 * | South Africa | Western Cape, Op-de-Tradouw | 2018/09/15 | Visser et al. 323 |
| 95 | 111_T.micro_A2 * | ||||||
| 96 | 112_T.micro_A3 * | ||||||
| 97 | Thesium multiramulosum Pilg. | Psilothesium | 113_T.multi_A1 * | South Africa | Limpopo, Uitsig | 2017/12/04 | Visser and le Roux 282 |
| 98 | 114_T.multi_A2 * | ||||||
| 99 | Thesium cf. nigromontanum Sond. | Frisea | 23_T.nigro_A1 * | South Africa | Western Cape, Franschoek Pass | 2018/09/12 | Visser and van Wyk 301 |
| 100 | 24_T.nigro_A2 * | ||||||
| 101 | 25_T.nigro_A3 * | ||||||
| 102 | Thesium ovatifolium N.Lombard and M.M.leRoux | Psilothesium | 116_T.ovati_A1 * | South Africa | KwaZulu-Natal, Ngome area | 2018/10/14 | Visser and Lombard 346 |
| 103 | 117_T.ovati_A2 * | ||||||
| 104 | Thesium pallidum A.DC. | Psilothesium | 118_T.palli_A1 * | South Africa | Free State, Platberg Nature Reserve | 2016/12/14 | Visser and le Roux 239 |
| 105 | 119_T.palli_A2 * | ||||||
| 106 | 120_T.palli_B1 * | South Africa | Eastern Cape, Maclear area | 2018/11/01 | Visser et al. 359 | ||
| 107 | Thesium procerum N.E.Br. | Psilothesium | 121_T.proce_A1 | South Africa | Gauteng, Pretoria National Botanical Garden | 2017/10/13 | Visser 276 |
| 108 | 122_T.proce_A2 | ||||||
| 109 | 123_T.proce_A3 | ||||||
| 110 | Thesium pubescens A.DC. | Frisea | 124_T.pubes_A1 * | South Africa | Western Cape, Citrusdal area | 2018/09/16 | Visser and van Wyk 331 |
| 111 | 125_T.pubes_A2 * | ||||||
| 112 | Thesium racemosum Bernh. | Psilothesium | 126_T.racem_A1 * | South Africa | Limpopo, Woodbush Forest Reserve area | 2017/12/04 | Visser and le Roux 288 |
| 113 | Thesium ramosum Hayne | Thesium | 127_T.ramos_A1 * | Lithuania | - | 2018/06 | Gudzinskas s.n. |
| 114 | 128_T.ramos_A2 * | ||||||
| 115 | 129_T.ramos_A3 * | ||||||
| 116 | Thesium resedoides A.W.Hill | Psilothesium | 130_T.resed_A1 * | South Africa | Gauteng, Dinokeng Nature Reserve | 2017/12/12 | Nel 481 |
| 117 | Thesium scandens Sond. | Discothesium (A.DC.) Zhigila, Verboom and Muasya | 131_T.scand_A1 * | South Africa | Eastern Cape, Paterson area | 2018/11/04 | Visser et al. 373 |
| 118 | 132_T.scand_A2 * | ||||||
| 119 | 133_T.scand_A3 * | ||||||
| 120 | 134_T.scand_B1 * | South Africa | Eastern Cape, Baviaan River Conservancy | 2018/11/05 | Visser et al. 376 | ||
| 121 | Thesium scirpioides A.W.Hill | Frisea | 135_T.scirp_A1 * | South Africa | Gauteng, Suikerbosrand Nature Reserve | 2016/12/12 | Visser and le Roux 232 |
| 122 | Thesium sonderianum Schltr. | Frisea | 136_T.sonde_A1 * | South Africa | Eastern Cape, Nieu-Bethesda area | 2018/11/06 | Visser et al. 378 |
| 123 | 137_T.sonde_A2 * | ||||||
| 124 | 138_T.sonde_A3 * | ||||||
| 125 | Thesium spicatum L. | Frisea | 103_T.spica_A1 | South Africa | Western Cape | 2017 | van Wyk s.n. |
| 126 | 104_T.spica_A2 | ||||||
| 127 | 105_T.spica_A3 | ||||||
| 128 | 106_T.spica_A4 | ||||||
| 129 | 107_T.spica_A5 | ||||||
| 130 | Thesium strictum P.J.Bergius | Frisea | 139_T.stric_A1 | South Africa | Western Cape | 2017 | van Wyk s.n. |
| 131 | 140_T.stric_A2 | ||||||
| 132 | 141_T.stric_A3 | ||||||
| 133 | 142_T.stric_B1 * | South Africa | Western Cape, Tradouw Pass | 2018/09/15 | Visser et al. 325 | ||
| 134 | 143_T.stric_B2 * | ||||||
| 135 | 144_T.stric_B3 * | ||||||
| 136 | 145_T.stric_C1 * | South Africa | Western Cape, Rooi-Els | 2018/09/14 | Visser et al. 309 | ||
| 137 | 146_T.stric_C2 * | ||||||
| 138 | 147_T.stric_C3 * | ||||||
| 139 | Thesium transvaalense Schltr. | Psilothesium | 148_T.trans_A1 * | South Africa | Gauteng, Waverley | 2016/10/22 | Visser and le Roux 201 |
| 140 | 149_T.trans_B1 * | South Africa | Gauteng, Waverley | 2016/10/22 | Visser and le Roux 200 | ||
| 141 | 150_T.trans_B2 * | ||||||
| 142 | Thesium triflorum L.f. | Discothesium | 26_T.trifl_A1 * | South Africa | KwaZulu-Natal, St. Lucia | 2018/02/02 | van Wyk s.n. |
| 143 | Thesium cf. ussanguense Engl. | Frisea | 27_T.ussan_A1 * | Mozambique | Manica, Serra Zuira | 1965/11/09 | Torre and Pereira 12.809 |
| 144 | 29_T.ussan_A3 * | ||||||
| 145 | Thesium vahrmeijeri Brenan | Psilothesium | 415_T.vahrm_A1 * | South Africa | KwaZulu-Natal, iSimangaliso Wetland Park | 2020/01/14 | Lombard and le Roux 415 |
| 146 | 418_T.vahrm_A2 * | South Africa | KwaZulu-Natal, iSimangaliso Wetland Park | 2020/01/15 | Lombard and le Roux 418 | ||
| 147 | Thesium virgatum Lam. | Frisea | 81_T.virgat_A1 * | South Africa | Western Cape, Franschoek area | 2018/09/12 | Visser and van Wyk 299B |
| 148 | Thesium zeyheri A.DC. | Psilothesium | 151_T.zeyhr_A1 * | South Africa | Free State, Platberg Nature Reserve | 2016/12/14 | Visser and le Roux 240 |
| 149 | 152_T.zeyhr_A2 * | ||||||
| 150 | 153_T.zeyhr_A3 * | ||||||
| 151 | 154_T.zeyhr_B1 * | South Africa | Eastern Cape, Maclear area | 2018/10/31 | Visser et al. 354 | ||
| 152 | 155_T.zeyhr_B2 * | ||||||
| 153 | 156_T.zeyhr_B3 * | ||||||
| 154 | 157_T.zeyhr_C1 * | South Africa | Eastern Cape, Rossouw-Indwe area | 2018/11/02 | Visser et al. 364 | ||
| 155 | 158_T.zeyhr_C2 * | ||||||
| 156 | 159_T.zeyhr_C3 * | ||||||
| 157 | Osyridicarpos schimperianus (Hochst. ex A.Rich.) A.DC. | - | Osyridicarpos_292 * | South Africa | Limpopo, Lekgalameetse Nature Reserve | 2017/12/05 | Visser and le Roux 292 |
| 158 | Lacomucinaea lineata (L.f.) Nickrent and M.A.García | - | Lacomucinaea_394 * | South Africa | Northern Cape, Postmasburg area | 2019/11/28 | Lombard and le Roux 394 |
| 159 | Lacomucinaea_401 * | South Africa | Northern Cape, Bergenaarspadpas area | 2019/11/29 | Lombard and le Roux 401 | ||
| 160 | Lacomucinaea_405 * | South Africa | Northern Cape, Witsand area | 2019/11/29 | Lombard and le Roux 405 | ||
| Retention Time | m/z [M-H]- | Formula | MS/MS | Standard | Identification | Reference | Number of Samples | Number of Species | |
|---|---|---|---|---|---|---|---|---|---|
| F1 | 3.05 | 191.0195 | C6H7O7 | 170, 155 | Citric acid | Citric acid | 38 | 26 | |
| F6 | 4.30 | 612.1365 | C26H30NO14S | 559, 377, 325, 279, 161, 97 | Unknown sulphated compound | 12 | 4 | ||
| F11 | 5.99 | 389.1207 | C20H21O8 | 165, 183, 209 | Unknown | 9 | 6 | ||
| F12 | 6.01 | 339.1283 | C14H25O12/C13H23O10 | 339, 215, 193/193 | Dehydrodiferulic acid | 20 | 10 | ||
| F16 | 6.32 | 215.0564 | C9H11O6 | 156, 155, 137, 111, 93 | Trimethyl Aconitate | 10 | 4 | ||
| F18 | 6.65 | 191.0555 | C7H11O6 | 129, 115, 101 | Quinic acid | 35 | 14 | ||
| F20 | 6.86 | 527.2552 | C19H43O16 | 361, 309, 263, 191, 179, 119, 89 | Unknown | 22 | 5 | ||
| F28 | 9.50 | 353.0873 | C16H17O9 | 191, 179, 135 | Neochlorogenic acid (3-caffeoylquinic acid) | [16] | 11 | 7 | |
| F33 | 10.01 | 173.0459 | C7H9O5 | 129, 111, 85 | Shikimic acid derivative | 18 | 7 | ||
| F41 | 11.08 | 337.0923 | C16H17O8 | 191, 163, 119 | Coumaroyl quinic acid | 19 | 9 | ||
| F49 | 11.67 | 353.0871 | C16H17O9 | 191 | Chlorogenic acid (5-caffeoylquinic acid) | 20 | 7 | ||
| F50 | 11.90 | 353.0865 | C16H17O9 | 191, 179, 173, 135 | Cryptochlorogenic acid (4-caffeoylquinic acid) | [16] | 25 | 12 | |
| F58 | 12.39 | 367.1033 | C17H19O9 | 193, 134 | Feruloylquinic acid | 11 | 7 | ||
| F78 | 13.78 | 337.0955 | C16H17O8 | 191, 173 | Coumaroyl quinic acid | 12 | 6 | ||
| F90 | 15.05 | 367.1046 | C17H19O9 | 193, 173 | Feruloylquinic acid | 7 | 6 | ||
| F94 | 15.10 | 367.1024 | C17H19O9 | 191 | Feruloylquinic acid | 14 | 7 | ||
| F100 | 15.49 | 755.2222 | C33H39O20 | 300, 271, 255 | Quercetin rhamnosyl-rhamnosyl-glucoside | 18 | 11 | ||
| F111 | 16.28 | 609.1619 | C27H29O16 | 300, 271, 255 | Quercetin robinobioside | 18 | 10 | ||
| F113 | 16.42 | 739.2306 | C30H43O21 | 284, 255, 227 | Kaempferol O-rhamnosyl-rhamnosyl-hexoside | 11 | 5 | ||
| F122 | 17.20 | 609.1416 | C27H29O16 | 300, 271, 255 | Rutin | Rutin | 80 | 27 | |
| F125 | 17.27 | 449.1227 | C21H21O11 | 287, 151, 135 | Eriodictyol -O-glucoside | 10 | 4 | ||
| F133 | 17.55 | 1043.2959 | C48H51O26 | 755, 609, 447 | Quercetin glycoside | 11 | 4 | ||
| F138 | 17.99 | 579.1522 | C23H31O17 | 300, 271, 255 | Quercetin pentoside hexoside | 11 | 5 | ||
| F145 | 18.81 | 593.1492 | C27H29O15 | 285, 255 | Kaempferol O-rutinoside | [17] | 36 | 15 | |
| F154 | 19.28 | 355.1417 | C17H23O8 | 193, 161 | Ferulic acid derivative | 12 | 6 | ||
| F156 | 19.36 | 623.1616 | C28H31O16 | 315, 300, 271 | Isorhamnetin O-glucoside O-rhamnoside | 51 | 20 | ||
| F173 | 21.46 | 933.269 | C43H49O23 | 661, 353 | Unknown | 10 | 4 | ||
| F177 | 22.11 | 933.2667 | C43H49O23 | 771, 661, 353 | Unknown | 10 | 3 | ||
| F193 | 23.93 | 301.0364 | C15H9O7 | 299, 271, 151 | Quercetin | 11 | 5 |
| Retention Time | m/z [M-H]- | Formula | MS/MSc | Lacomucinaea | Osyridicarpos | Thesium | Identification | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| F1 | 3.05 | 191.0195 | C6H7O7 | 170, 155 | + | + | + | Citric acid | |
| F18 | 6.65 | 191.0555 | C7H11O6 | 129, 115, 101 | + | Quinic acid | |||
| F25.4 | 8.76 | 463.1452 | C19H27O13 | 293, 191, 169, 149, 125, 89 | + | Unknown | |||
| F26 | 8.90 | 315.1080 | C14H19O8 | 153, 123 | + | + | Hydroxytyrosol O-glucoside | ||
| F28 | 9.50 | 353.0873 | C16H17O9 | 191, 179, 135 | + | + | + | Neochlorogenic acid (3-caffeoylquinic acid) | [16] |
| F50 | 11.90 | 353.0865 | C16H17O9 | 191, 179, 173, 135 | + | + | + | Cryptochlorogenic acid (4-caffeoylquinic acid) | [16] |
| F57 | 12.29 | 385.1160 | C17H21O10 | 223, 208, 179, 164 | + | + | Unknown, possibly sinapoyl hexose | ||
| F58 | 12.39 | 367.1033 | C17H19O9 | 193, 134 | + | + | + | Feruloylquinic acid | |
| F78.5 | 13.81 | 769.1800 | C33H37O21 | 667, 625 | + | Unknown | |||
| F90 | 15.05 | 367.1046 | C17H19O9 | 193, 173 | + | + | + | Feruloylquinic acid | |
| F94.1 | 15.15 | 753.1843 | C40H33O15 | 591, 489, 447, 191, 179, 161 | + | Unknown | |||
| F94.2 | 15.15 | 625.1400 | C27H29O17 | 367, 300, 191 | + | Quercetin-O-dihexoside | |||
| F114.1 | 16.58 | 601.2300 | C31H37O12 | 479, 171 | + | + | Unknown | ||
| F121.2 | 17.15 | 639.1600 | C28H31O17 | 315 | + | Unknown | |||
| F122 | 17.20 | 609.1416 | C27H29O16 | 300, 271, 255 | + | Rutin | |||
| F136 | 17.67 | 463.0978 | C25H19O9 | 300, 271, 255 | + | + | Quercetin-O-hexoside | ||
| F145 | 18.81 | 593.1492 | C27H29O15 | 285, 255 | + | Kaempferol O-rutinoside | [17] | ||
| F156 | 19.36 | 623.1616 | C28H31O16 | 315, 300, 271 | + | Isorhamnetin O-glucoside O-rhamnoside | |||
| F207 | 24.89 | 265.1429 | C15H21O4 | 97, 183 | + | + | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombard, N.; Stander, M.A.; Redelinghuys, H.; Le Roux, M.M.; Van Wyk, B.-E. A Study of Phenolic Compounds and Their Chemophenetic Value in the Genus Thesium (Santalaceae). Diversity 2022, 14, 590. https://doi.org/10.3390/d14080590
Lombard N, Stander MA, Redelinghuys H, Le Roux MM, Van Wyk B-E. A Study of Phenolic Compounds and Their Chemophenetic Value in the Genus Thesium (Santalaceae). Diversity. 2022; 14(8):590. https://doi.org/10.3390/d14080590
Chicago/Turabian StyleLombard, Natasha, Maria A. Stander, Herman Redelinghuys, Margaretha Marianne Le Roux, and Ben-Erik Van Wyk. 2022. "A Study of Phenolic Compounds and Their Chemophenetic Value in the Genus Thesium (Santalaceae)" Diversity 14, no. 8: 590. https://doi.org/10.3390/d14080590
APA StyleLombard, N., Stander, M. A., Redelinghuys, H., Le Roux, M. M., & Van Wyk, B.-E. (2022). A Study of Phenolic Compounds and Their Chemophenetic Value in the Genus Thesium (Santalaceae). Diversity, 14(8), 590. https://doi.org/10.3390/d14080590