

Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Taxa
2.2. Distribution
2.3. Study of Plant Communities
2.4. Study of Reproductive Traits
2.5. Age of Maturity
2.6. Statistical Analyses
3. Results
3.1. Distribution
3.2. Comparison of Reproductive Traits of Subspecies
3.2.1. Flowering and Fruiting
3.2.2. Maturation Age
3.3. Species Diversity in Invaded Communities
4. Discussion
4.1. Distribution
4.2. Comparison of Reproductive Traits of Subspecies
4.2.1. Flowering and Fruiting
4.2.2. Age of Maturity of Subspecies
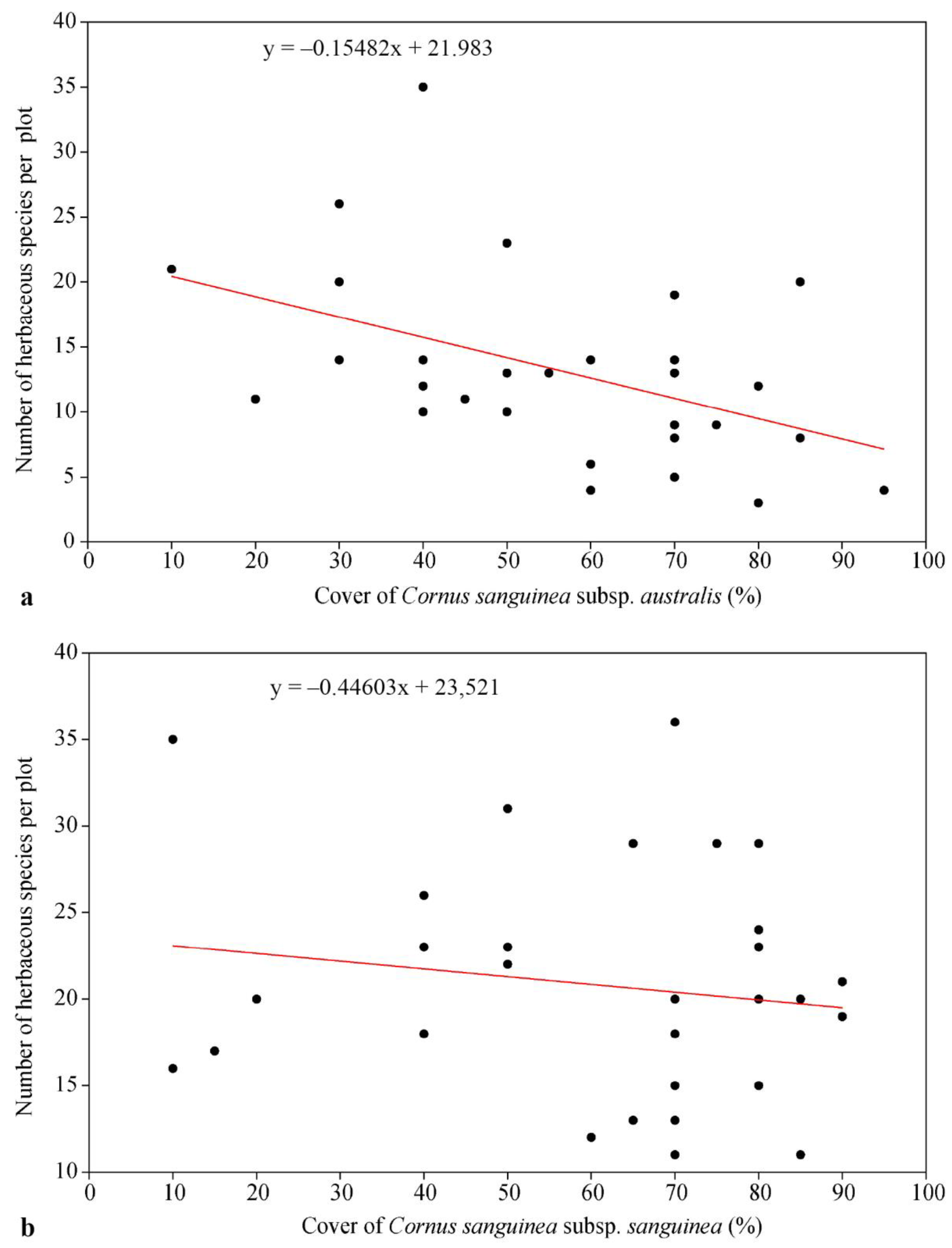
4.3. Species Diversity in Invaded Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Richardson, D.M. Pinus: A model group for unlocking the secrets of alien plant invasions? Preslia 2006, 78, 375–388. [Google Scholar]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Follak, S.; Bakacsy, L.; Essl, F.; Hochfellner, L.; Lapin, K.; Schwarz, M.; Tokarska-Guzik, B.; Wołkowycki, D. Monograph of invasive plants in Europe N°6: Asclepias syriaca L. Bot. Lett. 2021, 168, 422–451. [Google Scholar] [CrossRef]
- Verloove, F. Invaders in disguise. Conservation risks derived from misidentifications of invasive plants. Manag. Biol. Invasions 2010, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Saltonstall, K. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. PNAS 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [Green Version]
- Morais, P.; Reichard, M. Cryptic invasions: A review. Sci. Total Environ. 2018, 613, 1438–1448. [Google Scholar] [CrossRef]
- Jarić, I.; Heger, T.; Monzon, F.C.; Jeschke, J.M.; Kowarik, I.; McConke, K.R.; Pyšek, P.; Sagouis, A.; Essl, F. Crypticity in biological invasions. Trends Ecol. Evol. 2019, 34, 291–302. [Google Scholar] [CrossRef]
- Canavan, K.; Canavan, S.; Harms, N.E.; Lambertini, C.; Paterson, I.D.; Thum, R. The potential for biological control on cryptic plant invasions. Biol. Control 2020, 144, 104243. [Google Scholar] [CrossRef]
- Wahlsteen, E. Morphometrical characteristics of cryptic invasive and indigenous gene pools of field maple Acer campestre L. in southern Sweden. Nord. J. Bot. 2021, 39, e02901. [Google Scholar] [CrossRef]
- Hyldgaard, B.; Lambertini, C.; Brix, H. Phylogeography reveals a potential cryptic invasion in the Southern Hemisphere of Ceratophyllum demersum, New Zealand’s worst invasive macrophyte. Sci. Rep. 2017, 7, 16569. [Google Scholar] [CrossRef] [Green Version]
- Boeraeve, M.; D’hondt, B.; Verloove, F.; Vandekerkhove, K.; Van Landuyt, W.; Tamsyn, W.; Adriaens, T. “Lianification” or liana invasion? Commentary on Perring et al. (2020). Front. Ecol. Environ. 2021, 19, 375–376. [Google Scholar] [CrossRef]
- van Kleunen, M.; Weber, E.; Fischer, M. A meta analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, S.I.; Richardson, D.M. Invasive plants have broader physiological niches. PNAS 2014, 111, 10610–10614. [Google Scholar] [CrossRef] [Green Version]
- Rejmánek, M.; Richardson, D.M. What attributes make some plant species more invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Gudžinskas, Z.; Petrulaitis, L.; Žalneravičius, E. Emerging invasion threat of the liana Celastrus orbiculatus (Celastraceae) in Europe. NeoBiota 2020, 5, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Taura, L.; Gudžinskas, Z. Life stages and demography of invasive shrub Cytisus scoparius (Fabaceae) in Lithuania. Botanica 2020, 26, 1–14. [Google Scholar] [CrossRef]
- Xavier, C.N.; Granato-Souza, D.; Barbosa, A.C.; da Silva, J.R.M. Tropical dendrochronology applied to invasive tree species in the Brazilian Atlantic Forest. J. For. Res. 2021, 32, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Silveri, A.; Dunwiddie, P.W.; Michaels, H.J. Logging and edaphic factors in the invasion of an Asian woody vine in a mesic North American forest. Biol. Invasions 2001, 3, 379–389. [Google Scholar] [CrossRef]
- Mestre, L.; Chartier, M.; Renison, D. Effect of livestock on the growth of Rosa rubiginosa in a mountain range: A dendrochronological approach. Appl. Ecol. Environ. Res. 2014, 12, 855–866. [Google Scholar] [CrossRef]
- Richardson, D.M. Forestry trees as invasive aliens. Conserv. Biol. 1998, 12, 18–26. [Google Scholar] [CrossRef]
- van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.; González-Moreno, P.; Groom, Q.J.; Hulme, P.E.; et al. The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Křivánek, M.; Jarošík, V. Planting intensity, residence time, and species traits determine invasion success of alien woody species. Ecology 2009, 90, 2734–2744. [Google Scholar] [CrossRef] [PubMed]
- Collete, L.K.D.; Pither, J. Russian-olive (Elaeagnus angustifolia) Biology and ecology and its potential to invade Northern North American riparian ecosystems. Invasive Plant Sci. Manag. 2015, 8, 1–14. [Google Scholar] [CrossRef]
- Deckers, B.; Verheyen, K.; Hermy, M.; Muys, B. Effects of landscape structure on the invasive spread of black cherry Prunus serotina in an agricultural landscape in Flanders, Belgium. Ecography 2005, 28, 99–109. [Google Scholar] [CrossRef]
- Le Viol, I.; Julliard, R.; Kerbiriou, C.; De Redon, L.; Carnino, N.; Machon, N.; Porcher, E. Plant and spider communities benefit differently from the presence of planted hedgerows in highway verges. Biol. Conserv. 2008, 141, 1581–1590. [Google Scholar] [CrossRef] [Green Version]
- Blanusa, T.; Garratt, M.; Cathcart-James, M.; Hunt, L.; Cameron, R.W. Urban hedges: A review of plant species and cultivars for ecosystem service delivery in north-west Europe. Urban For. Urban Green 2019, 44, 126391. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Standish, R.J.; Stock, W.D.; Valladares, F. Plant functional traits of dominant native and invasive species in mediterranean-climate ecosystems. Ecology 2016, 97, 75–83. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Hejda, M.; Sádlo, J.; Kutlvašr, J.; Petřík, P.; Vítková, M.; Vojík, M.; Pyšek, P.; Pergl, J. Impact of invasive and native dominants on species richness and diversity of plant communities. Preslia 2021, 93, 181–201. [Google Scholar] [CrossRef]
- Tanner, R.; Branquart, E.; Brundu, G.; Buholzer, S.; Chapman, D.; Ehret, P.; van Valkenburg, J. The prioritisation of a short list of alien plants for risk analysis within the framework of the Regulation (EU) No. 1143/2014. NeoBiota 2017, 35, 87–118. [Google Scholar] [CrossRef]
- Yazlık, A.; Ambarlı, D. Do non-native and dominant native species carry a similar risk of invasiveness? A case study for plants in Turkey. NeoBiota 2022, 76, 53–72. [Google Scholar] [CrossRef]
- Kowarik, I. Time lags in biological invasions with regard to the success and failure of alien species. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 15–38. [Google Scholar]
- Petrulaitis, L.; Gudžinskas, Z. The first records of two alien woody species, Cornus alternifolia and Cornus amomum, in Lithuania. BioInv Rec. 2020, 9, 384–392. [Google Scholar] [CrossRef]
- Medvecká, J.; Kliment, J.; Májeková, J.; Halada, Ľ.; Zaliberová, M.; Gojdičová, E.; Feráková, V.; Jarolímek, I. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. [Google Scholar]
- Azola, T.; Bakan, B. Popis tujerodnih rastlinskih vrst v Mestni občini Maribor [Inventory of alien plant species in the Municipality of Maribor]. Hladnikia 2021, 47, 3–16. [Google Scholar]
- Gudžinskas, Z.; Petrulaitis, L. New alien taxa of the genus Cornus (Cornaceae) recorded in Lithuania and Latvia. Botanica 2021, 27, 160–169. [Google Scholar] [CrossRef]
- Kelly, D.L. Cornus sericea L. in Ireland: An incipient weed of wetlands. Watsonia 1990, 18, 33–36. [Google Scholar]
- Bačič, T.; Strgulc Krajšek, S.; Jogan, N. Red osier dogwood (Cornus sericea L.)—A new invasive species in Slovenian flora. Acta Biol. Slov. 2015, 58, 13–21. (In Slovenian) [Google Scholar]
- Bomanowska, A.; Adamowski, W.; Kirpluk, I.; Otreba, A.; Rewicz, A. Invasive alien plants in Polish national parks—Threats to species diversity. PeerJ 2019, 7, e8034. [Google Scholar] [CrossRef] [Green Version]
- Gregor, T.; Kasperek, G. Non-native woody plant species in urban forests of Frankfurt/Main (Germany). Tuexenia 2021, 41, 133–145. [Google Scholar] [CrossRef]
- Riebl, R.; Meve, U.; Aas, G. Morphologische Variabilität und taxonomische Differenzierung von Cornus sanguinea: Nordbayerische Naturstandorte und Strassenbegleitgrün im Vergleich. Ber. Bayer. Bot. Ges. Erforsch. Heim. Flora 2017, 87, 39–54. [Google Scholar]
- Verloove, F. Fraxinus pennsylvanica, Pterocarya fraxinifolia en andere opmerkelijke uitheemse rivierbegeleiders in België en NW-Frankrijk. Dumortiera 2011, 99, 1–10. [Google Scholar]
- Poyarkova, A.I. Cornaceae. In Flora of USSR; Shishkin, B.K., Ed.; Smithsonian Libraries: Washington, DC, USA, 1973; Volume 17, pp. 225–249. [Google Scholar]
- Eyde, R.H. Comprehending Cornus: Puzzles and progress in the systematics of dogwoods. Bot. Rev. 1988, 54, 233–351. [Google Scholar] [CrossRef]
- Woźnicka, A.; Melosik, I.; Morozowska, M. Quantitative and qualitative differences in morphological traits of endocarps revealed between Cornus L. species. Plant Syst. Evol. 2015, 301, 291–308. [Google Scholar] [CrossRef] [Green Version]
- Morozowska, M.; Woźnicka, A.; Nowińska, R. The morphology and internal structure of dogwood (Cornus L.) endocarps in the taxonomy and phylogeny of the genus. PeerJ 2021, 9, e12170. [Google Scholar] [CrossRef]
- Schulz, B. Die Gattung Cornus (Cornaceae), Hartriegel und Kornelkirsche, Teil 3. Mitt. Deutsch. Dendrol. Ges. 2012, 97, 91–132. [Google Scholar]
- Grosset, E.G. The geographical distribution of Cornus sanguinea s. l. and history of the broad-leaved forests of the European part of the U.S.S.R. Bull. Soc. Nat. Mosc. Biol. Ser. 1958, 63, 77–85. [Google Scholar]
- Novák, P.; Zukal, D.; Harásek, M.; Vlčková, P.; Abdaladze, O.; Willner, W. Ecology and vegetation types of oak-hornbeam and ravine forests of the Eastern Greater Caucasus, Georgia. Folia Geobot. 2020, 55, 333–349. [Google Scholar] [CrossRef]
- Essl, F.; Bacher, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kowarik, I.; Kühn, I.; Pyšek, P.; Rabitsch, W.; et al. Which taxa are alien? Criteria, applications, and uncertainties. Bioscience 2018, 68, 496–509. [Google Scholar] [CrossRef] [Green Version]
- Maes, B.; van Loon, R. Wilde en niet-wilde bomen en struiken in vegetatieopnamen. Een voorstel voor een methode van specifieke notering van bomen en struiken in vegetatieopnamen en overwegingen daarbij. Stratiotes 2020, 55, 19–31. [Google Scholar]
- Shynder, O.; Negrash, J. Checklist of the flora of the vicinity of Balakliya (Kharkiv region, Ukraine): Native and alien taxa, distribution of rare plants, new findings. Plant Introd. 2021, 89–90, 13–71. [Google Scholar] [CrossRef]
- Orlov, O.O.; Shynder, O.I.; Vorobjov, E.O.; Gryb, O.V. New floristic finds in the Forest-Steppe part of Zhytomyr Region. Ukr. Bot. J. 2022, 79, 6–26. [Google Scholar] [CrossRef]
- Convention on Biological Diversity. Pathways of Introduction of Invasive Species, Their Prioritization and Management. 2014. Available online: https://www.cbd.int/doc/meetings/sbstta/sbstta-18/official/sbstta-18-09-add1-en.pdf (accessed on 25 October 2022).
- Gudžinskas, Z. Genus Ambrosia L. (Asteraceae) in Lithuania. Thaiszia 1993, 3, 89–96. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde. Dritte Auflage; Springer: New York, NY, USA, 1964; p. 865. [Google Scholar] [CrossRef]
- Lenth, R.V. Some practical guidelines for effective sample size determination. Am. Stat. 2001, 55, 187–193. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Crooks, J. Lag times and exotic species: The ecology and management of biological invasions in slow-motion. Ecoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Simberloff, D. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Krüsi, B.O.; Debussche, M. The fate of flowers and fruits of Cornus sanguinea L. in three contrasting Mediterranean habitats. Oecologia 1988, 74, 592–599. [Google Scholar] [CrossRef]
- Kollmann, J.; Grubb, P.J. Biological flora of Central Europe: Cornus sanguinea L. Flora 2001, 196, 161–179. [Google Scholar] [CrossRef]
- Holub, J. Cornaceae Dumort.—Dřínovité. In Květena České republiky; Slavík, B., Chrtek, J., Tomšovic, P., Eds.; Academia: Praha, Czech Republic, 1997; Volume 5, pp. 252–264. [Google Scholar]
- Borowicz, V.A.; Stephenson, A.G. Fruit composition and patterns of fruit dispersal of two Cornus spp. Oecologia 1985, 67, 435–441. [Google Scholar] [CrossRef]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Parker, J.D.; Torchin, M.E.; Hufbauer, R.A.; Lemoine, N.P.; Alba, C.; Blumenthal, D.M.; Bossdorf, O.; Byers, J.E.; Dunn, A.M.; Heckman, R.W.; et al. Do invasive species perform better in their new ranges? Ecology 2013, 94, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudžinskas, Z.; Taura, L. Do Reproductive Traits of Invasive Populations of Scotch Broom, Cytisus scoparius (Fabaceae), Outperform Native Populations? Plants 2022, 11, 2158. [Google Scholar] [CrossRef] [PubMed]
- Guitián, J.; Guitián, P.; Navarro, L. Fruit set, fruit reduction, and fruiting strategy in Cornus sanguinea (Cornaceae). Am. J. Bot. 1996, 83, 744–748. [Google Scholar] [CrossRef]
- Schurr, F.M.; Steinitz, O.; Nathan, R. Plant fecundity and seed dispersal in spatially heterogeneous environments: Models, mechanisms and estimation. J. Ecol. 2008, 96, 628–641. [Google Scholar] [CrossRef]
- Petrulaitis, L.; Rašomavičius, V.; Uogintas, D.; Gudžinskas, Z. Soil seed bank of alien and native Cornus (Cornaceae) taxa in Lithuania: What determines seed density and vertical distribution in soil? Diversity 2022, 14, 488. [Google Scholar] [CrossRef]
- Verloove, F. Cornus sanguinea. From: Manual of the Alien Plants of Belgium. Botanic Garden Meise, Belgium. 2021. Available online: www.alienplantsbelgium.be (accessed on 15 October 2021).
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 193, pp. 97–125. [Google Scholar] [CrossRef]
- Zalba, S.M.; Amodeo, M.R. Site-specific reproductive performance of an invasive fleshy-fruited tree in remnant grasslands of the Argentinean Pampas. Weed Res. 2015, 55, 546–554. [Google Scholar] [CrossRef]
- Serviss, B.E.; Tumlison, R. Guide to the naturalized, escaped, and adventive woody flora of Arkansas. Phytoneuron 2021, 29, 1–193. [Google Scholar]
- Grubb, P.J.; Kollmann, J.; Lee, W.G. A Garden Experiment on Susceptibility to Rabbit-Grazing, Sapling Growth Rates, and Age at First Reproduction for Eleven European Woody Species. Plant Biol. 1999, 1, 226–234. [Google Scholar] [CrossRef]
- Chevin, L.-M.; Collins, S.; Lefèvre, F. Phenotypic plasticity and evolutionary demographic responses to climate change: Taking theory out to the field. Funct. Ecol. 2013, 27, 967–979. [Google Scholar] [CrossRef] [Green Version]
- He, Y. The Relationship between an Invasive Shrub and Soil Moisture: Seasonal Interactions and Spatially Covarying Relations. ISPRS Int. J. Geo-Inf. 2014, 3, 1139–1153. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Classen, A.T.; Simberloff, D. Two co-occurring invasive woody shrubs alter soil properties and promote subdominant invasive species. J. Appl. Ecol. 2014, 51, 124–133. [Google Scholar] [CrossRef] [Green Version]
- De Barros Ruas, R.; Costa, L.M.S.; Bered, F. Urbanization driving changes in plant species and communities—A global view. Glob. Ecol. Conserv. 2022, 38, e02243. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subspecies | Site Name | Administrative Unit | Longitude (°E) | Latitude (°N) |
|---|---|---|---|---|
| Cornus sanguinea subsp. sanguinea | Piepaliai | Kaunas distr. | 55.07423 | 23.78346 |
| Cornus sanguinea subsp. sanguinea | Veliuona | Jurbarkas distr. | 55.08996 | 23.18326 |
| Cornus sanguinea subsp. sanguinea | Šilinė | Jurbarkas distr. | 55.08856 | 22.94780 |
| Cornus sanguinea subsp. australis | Ginkūnai | Šiauliai distr. | 55.95343 | 23.37109 |
| Cornus sanguinea subsp. australis | Kaišiadorys | Kaišiadorys city | 54.87099 | 24.43677 |
| Cornus sanguinea subsp. australis | Maišiagala | Vilnius distr. | 54.86959 | 25.05138 |
| Taxa and Sites | Age (years) | Shoot Diameter (mm) | ||||
|---|---|---|---|---|---|---|
| Min–Max | Mean | Median | Min–Max | Mean | Median | |
| Cornus sanguinea subsp. australis | ||||||
| Kaišiadorys | 3–6 | 3.9 ± 0.7 a | 4 | 5.0–8.8 | 7.18 ± 1.28 a | 7.7 |
| Vievis | 3–5 | 3.5 ± 0.6 a | 3 | 4.9–8.8 | 6.01 ± 0.90 b | 5.7 |
| Pooled | 3–6 | 3.7 ± 0.7 A | 4 | 4.9–8.8 | 6.60 ± 1.24 A | 6.5 |
| Cornus sanguinea subsp. sanguinea | ||||||
| Piepaliai | 4–6 | 5.4 ± 0.7 b | 5 | 7.5–13.0 | 10.90 ± 1.62 c | 11.3 |
| Veliuona | 4–6 | 5.2 ± 0.6 b | 5 | 8.0–14.6 | 12.18 ± 1.79 d | 12.5 |
| Pooled | 4–6 | 5.3 ± 0.1 B | 5 | 7.5–14.6 | 11.55 ± 1.80 B | 11.9 |
| Factor | Sum of Squares | df | Mean Square | F | p |
|---|---|---|---|---|---|
| Shoot age | |||||
| Taxon (f) | 76.80 | 1 | 76.80 | 168.30 | <0.001 |
| Habitat (r) | 2.26 | 2 | 1.13 | 2.48 | 0.088 |
| Within | 52.93 | 116 | 0.46 | ||
| Total | 132.00 | 119 | |||
| Shoot diameter | |||||
| Taxon (f) | 737.55 | 1 | 737.55 | 357.00 | <0.001 |
| Habitat (r) | 43.84 | 2 | 21.92 | 10.61 | <0.001 |
| Within | 239.67 | 116 | 2.07 | ||
| Total | 1021.06 | 119 |
| Parameters | All Plots | Woodlands | Shrublands | Woodlands | Shrublands | |
|---|---|---|---|---|---|---|
| Cornus sanguinea | Cornus sanguinea | Cornus sanguinea | ||||
| subsp. australis | subsp. sanguinea | subsp. australis | subsp. australis | subsp. sanguinea | subsp. sanguinea | |
| Cover of Cornus (%) | 56.6 ± 21.6 ns | 63.5 ± 23.5 ns | 59.2 ± 21.2 a | 52.3 ± 22.6 a | 65.9 ± 24.2 a | 60.7 ± 23.0 a |
| Cover of trees (%) | 37.2 ± 25.7 ns | 31.6 ± 30.7 ns | 53.7 ± 14.9 a | 8.6 ± 10.0 b | 57.3 ± 14.5 a | 0.4 ± 1.3 c |
| Cover of shrubs (%) | 64.7 ± 24.9 * | 78.1 ± 17.1 * | 65.8 ± 26.1 a | 62.7 ± 23.7 a | 76.5 ± 19.0 a | 80.0 ± 14.8 a |
| Cover of herbaceous species (%) | 38.5 ± 31.8 ns | 42.1 ± 28.0 ns | 32.0 ± 29.1 a | 49.6 ± 34.4 ab | 31.7 ± 22.9 a | 54.6 ± 29.2 b |
| Cover of bryophytes (%) | 13.7 ± 20.3 ns | 26.1 ± 30.8 ns | 13.2 ± 17.9 a | 14.6 ± 24.9 a | 22.2 ± 27.9 a | 30.8 ± 34.4 a |
| Total number of plant species | 22.6 ± 8.2 ns | 24.6 ± 6.4 ns | 22.9 ± 7.6 a | 22.2 ± 9.5 a | 23.4 ± 7.1 a | 26.1 ± 5.3 a |
| Number of native vascular plant species | 15.4 ± 8.8 *** | 22.7 ± 6.2 *** | 15.9 ± 6.4 ns | 14.4 ±12.1 ns | 21.5 ± 6.7 ns | 24.1 ± 5.3 ns |
| Total number of woody species | 8.2 ± 4.3 ns | 7.0 ± 2.7 ns | 9.9 ± 4.1 a | 5.3 ± 2.9 b | 8.3 ± 2.5 a | 5.4 ± 2.0 b |
| Total number of herbaceous plant species | 12.9 ± 6.8 * | 15.8 ± 5.8 * | 11.6 ± 5.0 a | 15.3 ± 8.9 ab | 13.0 ± 4.7 a | 19.2 ± 5.3 b |
| Number of alien species | 3.5 ± 1.5 *** | 0.6 ± 1.0 *** | 3.8 ± 1.6 a | 2.9 ± 1.4 a | 0.5 ± 0.9 b | 0.8 ± 1.2 b |
| Number of alien woody species | 3.1 ± 1.6 *** | 0.3 ± 0.5 *** | 3.6 ± 1.6 a | 2.3 ± 1.0 b | 0.2 ± 0.4 c | 0.4 ± 0.6 c |
| Number of alien herbaceous species | 0.4 ± 0.7 ns | 0.4 ± 0.6 ns | 0.3 ± 0.4 a | 0.6 ± 1.0 a | 0.3 ± 0.6 a | 0.4 ± 0.8 a |
| Number of bryophyte species | 1.5 ± 1.5 ns | 1.2 ± 1.6 ns | 1.4 ± 1.5 a | 1.6 ± 1.6 a | 1.6 ± 1.6 a | 0.7 ± 1.3 a |
| Group of Habitats | Woodlands | Shrublands | ||
|---|---|---|---|---|
| Subspecies | Cornus sanguinea | Cornus sanguinea | ||
| subsp. australis | subsp. sanguinea | subsp. australis | subsp. sanguinea | |
| Number of study plots | 19 | 16 | 11 | 14 |
| Total number of species | 160 | 136 | 140 | 170 |
| Total number of woody species | 55 | 33 | 29 | 30 |
| Total number of herbaceous species | 94 | 95 | 90 | 132 |
| Number of bryophyte species | 11 | 8 | 11 | 8 |
| Total number of alien species | 27 | 6 | 15 | 9 |
| Number of alien woody species | 25 | 3 | 10 | 4 |
| Number of alien herbaceous species | 2 | 3 | 5 | 5 |
| Woody species | ||||
| Cornus sanguinea L. subsp. australis (C.A.Mey.) Jáv. | 100 | 0 | 100 | 0 |
| Cornus sanguinea L. subsp. sanguinea | 0 | 100 | 0 | 100 |
| Acer platanoides L. | 90 | 65 | 27 | 21 |
| Rubus caesius L. | 53 | 63 | 18 | 36 |
| Prunus padus L. | 47 | 59 | 18 | 43 |
| Sorbus aucuparia L. | 42 | 6 | 27 | 21 |
| Corylus avelana L. | 32 | 59 | 0 | 21 |
| Euonymus europaeus L. | 21 | 12 | 9 | 64 |
| Lonicera xylosteum L. | 16 | 41 | 9 | 14 |
| Rhamnus cathartica L. | 11 | 29 | 0 | 50 |
| Herbaceous species | ||||
| Veronica chamaedrys L. | 58 | 31 | 64 | 36 |
| Geum urbanum A. Gray | 53 | 31 | 18 | 21 |
| Galium aparine L. | 42 | 19 | 18 | 7 |
| Aegopodium podagraria L. | 32 | 44 | 9 | 36 |
| Carex hirta L. | 26 | 19 | 45 | 7 |
| Calamagrostis epigejos (L.) Roth. | 21 | 13 | 55 | 29 |
| Taraxacum officinale F.H. Wigg. | 21 | 6 | 55 | 14 |
| Vicia cracca L. | 21 | 0 | 45 | 36 |
| Equisetum arvense L. | 11 | 31 | 55 | 43 |
| Dactylis glomerata L. | 11 | 0 | 45 | 21 |
| Poa pratensis L. | 11 | 0 | 45 | 43 |
| Agrimonia eupatoria L. | 0 | 6 | 0 | 43 |
| Achillea millefolium L. | 0 | 0 | 27 | 43 |
| Briza media L. | 0 | 0 | 0 | 43 |
| Medicago falcata L. | 0 | 0 | 9 | 43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrulaitis, L.; Gudžinskas, Z. Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania. Diversity 2023, 15, 107. https://doi.org/10.3390/d15010107
Petrulaitis L, Gudžinskas Z. Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania. Diversity. 2023; 15(1):107. https://doi.org/10.3390/d15010107
Chicago/Turabian StylePetrulaitis, Lukas, and Zigmantas Gudžinskas. 2023. "Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania" Diversity 15, no. 1: 107. https://doi.org/10.3390/d15010107
APA StylePetrulaitis, L., & Gudžinskas, Z. (2023). Drivers and Effects of Cryptic Invasion of Cornus sanguinea subsp. australis in Lithuania. Diversity, 15(1), 107. https://doi.org/10.3390/d15010107