
Functional Trait Responses of C4 Bunchgrasses to Fire Return Intervals in the Semi-Arid Savanna of South Africa

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
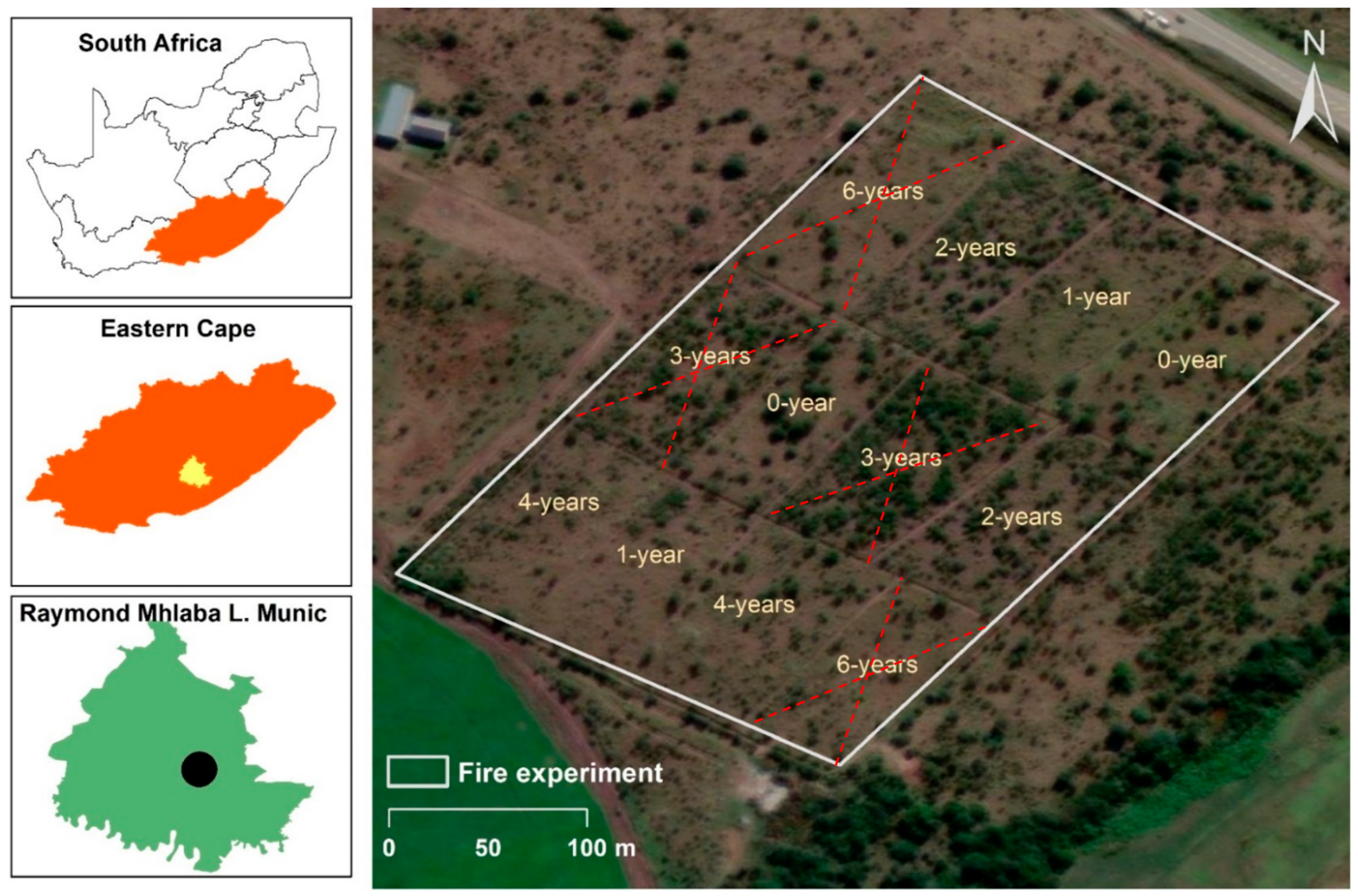
2.1. Study Area
2.2. Experimental Design and Layout
2.3. Species and Data Collection
2.4. Statistical Analysis
3. Results
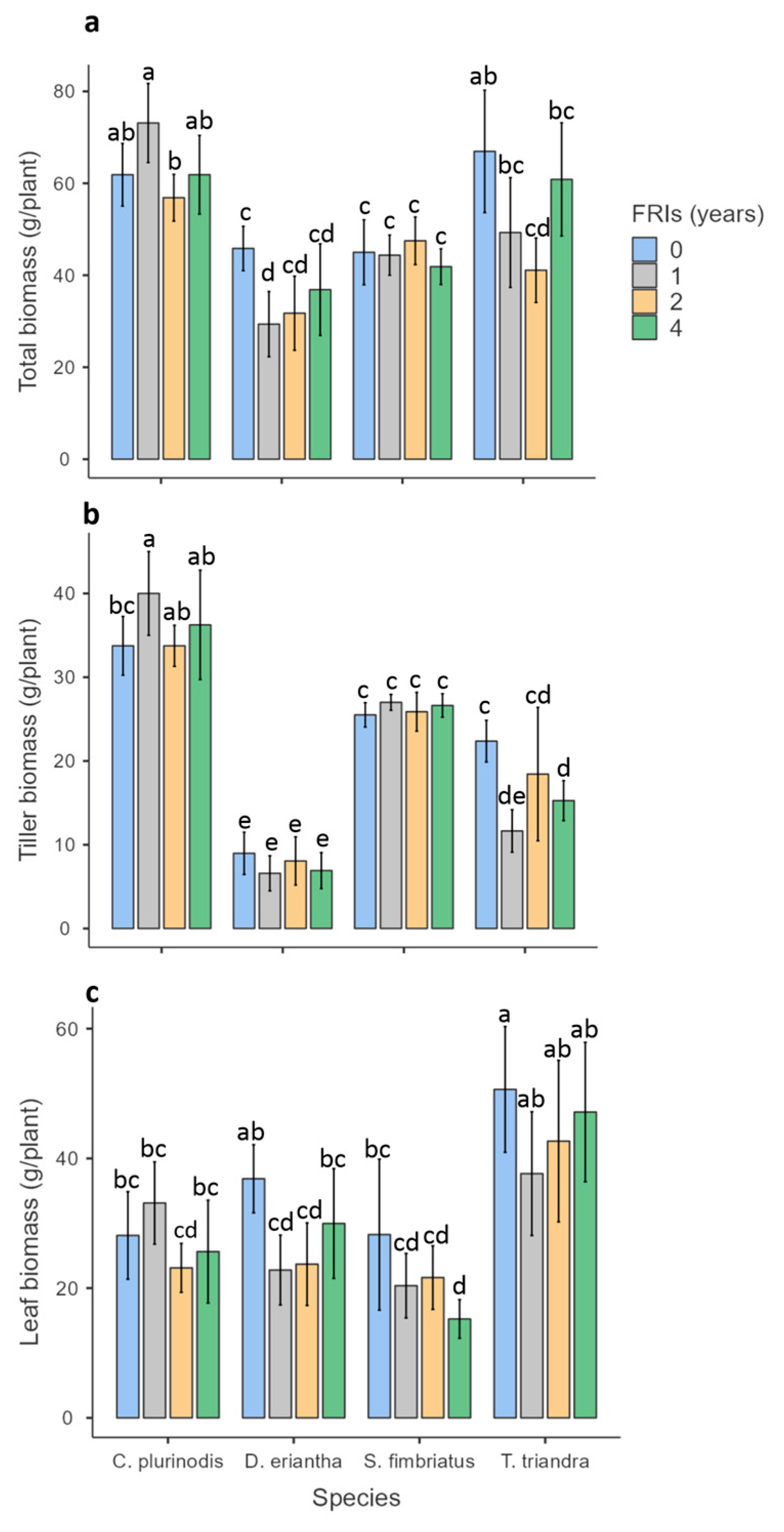
3.1. Biomass Production
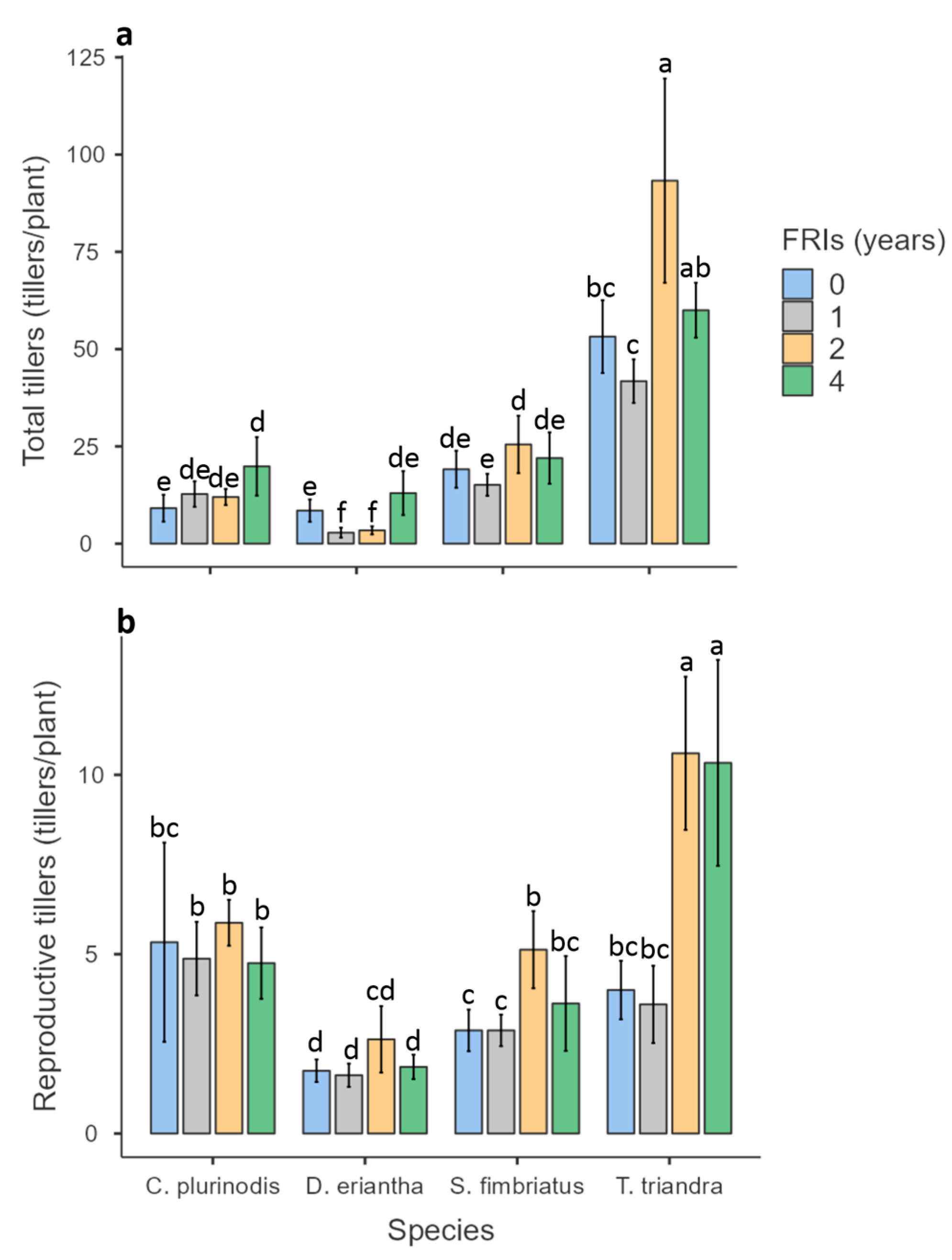
3.2. Tiller Production
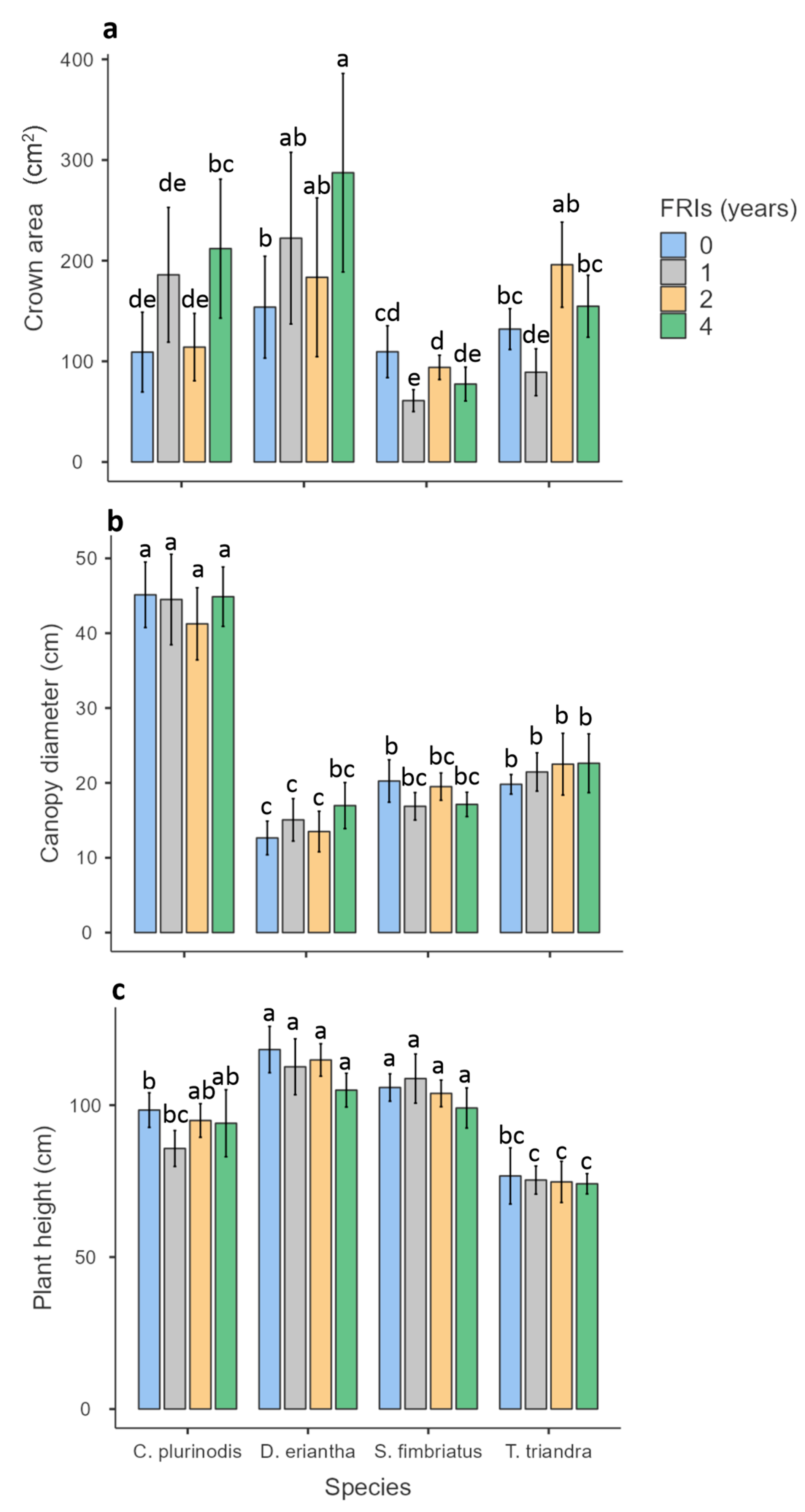
3.3. Canopy and Crown Size
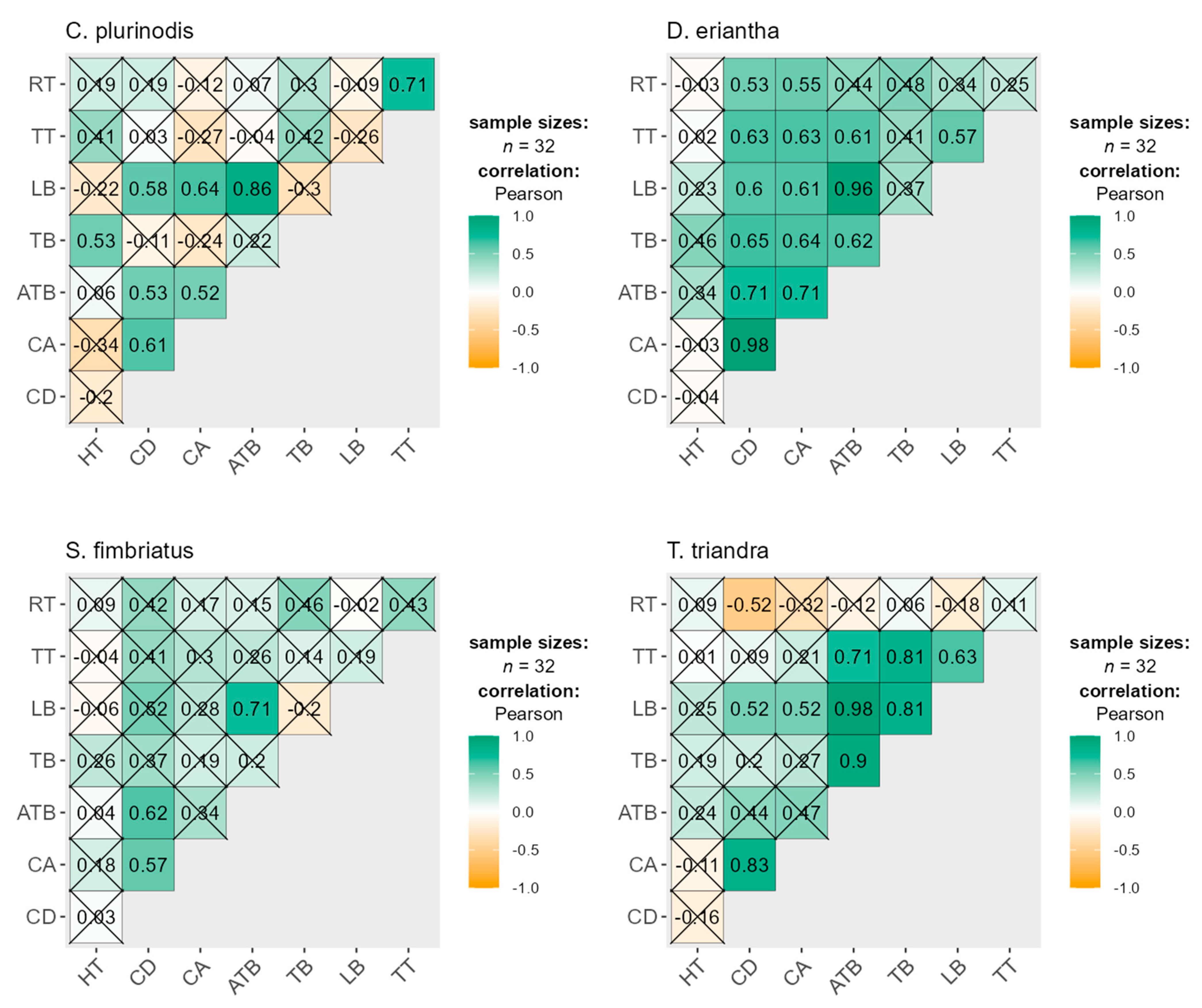
3.4. Relationships between Functional Traits
4. Discussion
4.1. Effect of Varying Fire Return Intervals on Grass Biomass Production
4.2. Effect of Fire Return Intervals on Tillering, Crown Size, and Flowering
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durigan, G.; Pilon, N.A.L.; Abreu, R.C.R.; Hoffmann, W.A.; Martins, M.; Fiorillo, B.F.; Antunes, A.Z.; Carmignotto, A.P.; Maravalhas, J.B.; Vieira, J.; et al. No Net Loss of Species Diversity After Prescribed Fires in the Brazilian Savanna. Front. For. Glob. 2020, 3, 13. [Google Scholar] [CrossRef]
- Baggio, R.; Overbeck, G.E.; Durigan, G.; Pillar, V.D. To graze or not to graze: A core question for conservation and sustainable use of grassy ecosystems in Brazil. Perspect. Ecol. Conserv. 2021, 19, 256–266. [Google Scholar] [CrossRef]
- Mndela, M.; Thamaga, K.H.; Gusha, B. A global perspective of functional trait responses of graminoids to seasonality of fire. Fire 2023, 9, 329. [Google Scholar] [CrossRef]
- Simpson, K.J.; Olofsson, J.K.; Ripley, B.S.; Osborne, C.P. Frequent fires prime plant developmental responses to burning. Proc. Royal Soc. 2019, 286, 20191315. [Google Scholar] [CrossRef] [PubMed]
- Hoetzel, S.; Dupont, L.; Schefub, E.; Rommerskirchen, F.; Wefer, G. The role of fire in Miocene to Pliocene C4 grassland and ecosystem evolution. Nat. Geosci. 2013, 6, 1027–1030. [Google Scholar] [CrossRef]
- Moore, N.A.; Camac, J.S.; Morgan, J.W. Effects of drought and fire on resprouting capacity of temperate Australian perennial native grasses. New Phytol. 2018, 221, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Ripley, B.; Visser, V.; Christin, P.A.; Archibald, S.; Martin, T.; Osborne, C. Fire ecology of C3 and C4 grasses depends on evolutionary history and frequency of burning but not photosynthetic type. Ecology 2015, 96, 2679–2691. [Google Scholar] [CrossRef]
- Russell, M.L.; Vermeire, L.T.; Ganguli, A.C.; Hendrickson, J.R. Fire Return Interval and Season of Fire Alter Bud Banks. Range Ecol. Manag. 2019, 72, 542–550. [Google Scholar] [CrossRef]
- Dagleish, H.J.; Ott, J.P.; Setshogo, M.P.; Hartnet, D.C. Inter-specific variation in bud banks and flowering effort among semi-arid African savanna grasses. S. Afr. J. Bot. 2012, 83, 127–133. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimešová, J.; Hartnett, D.C. The ecology and signifcance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.J.; Jardine, E.C.; Archibald, S.; Forrestel, E.J.; Lehmann, C.E.R.; Thomas, G.H.; Osborne, C.P. Resprouting grasses are associated with less frequent fire than seeders. New Phytol. 2021, 230, 832–844. [Google Scholar] [CrossRef]
- Hiers, Q.A.; Treadwell, M.L.; Dickinson, M.B.; Kavanagh, K.L.; Lodge, A.G.; Starns, H.D.; Tolleson, D.R.; Twidwell, D.; Wonkka, C.L.; Rogers, W.E. Grass bud responses to fire in a semiarid savanna system. Ecol. Evol. 2021, 11, 6620–6633. [Google Scholar] [CrossRef]
- Bombo, A.B.; Appezzato da Glória, B.; Fidelis, A. Fire exclusion changes belowground bud bank and bud bearing organ composition jeopardizing open savanna resilience. Oecologia 2022, 199, 153–164. [Google Scholar] [CrossRef]
- Keleey, J.E.; Pausas, J.G.; Rundell, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.J.; Ripley, B.S.; Christin, P.A.; Belcher, C.M.; Lehmann, C.E.R.; Thomas, G.H.; Osborne, C.P. Determinants of flammability in savanna grass species. J. Ecol. 2016, 104, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Enright, N.J.; Fontaine, J.B.; Lamont, B.B.; Miller, B.P.; Westcott, V.C. Resistance and resilience to changing climate and fire regime depend on plant functional traits. J. Ecol. 2014, 102, 1572–1581. [Google Scholar] [CrossRef]
- Te Beest, M.; Kleinjan, A.; Tuijnman, V.; Findlay, N.; Mvelase, T.; le Roux, E.; Tedder, M.; Gordijn, P.; Janse van Rensburg, S. Grass functional trait responses to experimental warming and fire in Afromontane grasslands. Afr. J. Range Forag. Sci. 2021, 38, 88–101. [Google Scholar] [CrossRef]
- Oluwole, F.A.; Sambo, J.M.; Dube, D. Long-term effects of different burning frequencies on the dry savannah grassland in South Africa. Afr. J. Agric. Res. 2008, 3, 147–153. [Google Scholar]
- Maroyi, A. Diversity of use and local knowledge of wild and cultivated plants in the Eastern Cape province, South Africa. J. Ethnobiol. Ethnomed. 2017, 13, 2–16. [Google Scholar] [CrossRef]
- Parwada, C.; Magomani, M.I.; van Tol, J.J. Impacts of different prescribed fire frequencies on selected soil chemical properties in a semi-arid savannah thornveld. Cogent Environ. Sci. 2020, 6, 1868171. [Google Scholar] [CrossRef]
- Mandiringana, O.T.; Mnkeni, P.N.S.; Mkile, Z.; Van Averbeke, W.; Van Ranste, V.H. Mineralogy and fertility status of selected soils of the Eastern Cape Province, South Africa. Commun. Soil Sci. Plant Anal. 2005, 36, 2431–2446. [Google Scholar] [CrossRef]
- Buitenwerf, R.; Bond, W.J.; Stevens, N.; Trollope, W.S.W. Increased tree densities in south african savannas: >50 years of data suggests CO2 as a driver. Glob. Chang. Biol. 2011, 18, 675–684. [Google Scholar] [CrossRef]
- Van Oudtshoorn, F. Guide to Grasses of Southern Africa; Briza Publications: Pretoria, South Africa, 1999. [Google Scholar]
- Wigley, B.J.; Charles-Dominique, T.; Hempson, G.P.; Stevens, N.; Te Beest, M.; Archibald, S.; Bond, W.J.; Bunney, K.; Coetsee, C.; Donaldson, J.; et al. A handbook for the standardised sampling of plant functional traits in disturbance-prone ecosystems, with a focus on open ecosystems. Aust. J. Bot. 2020, 68, 473–531. [Google Scholar] [CrossRef]
- Gordijn, P.J.; O’Connor, T.G. Multidecadal effects of fire in a grassland biodiversity hotspot: Does pyrodiversity enhance plant diversity? Ecol. Appl. 2021, 3, e02391. [Google Scholar] [CrossRef] [PubMed]
- Benson, E.J.; Hartnett, D.C.; Mann, K.H. Belowground bud banks and meristem limitation in tallgrass prairie plant populations. Am. J. Bot. 2004, 91, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Paula, S. Grasses and fire: The importance of hiding buds: A response to Moore et al. (2019) ‘Effects of drought and fire on resprouting capacity of 52 temperate Australian perennial native grasses’. New Phytol. 2020, 226, 957–959. [Google Scholar] [CrossRef]
- Rowe, H.I. Tricks of the trade: Techniques and opinions from 38 experts in tallgrass prairie restoration. Restor. Ecol. 2010, 18, 253–262. [Google Scholar] [CrossRef]
- Ellsworth, L.M.; Wrobleski, D.W.; Kauffman, J.B.; Reis, S.A. Ecosystem resilience is evident 17 years after fire in Wyoming big sagebrush ecosystems. Ecosphere 2016, 7, e01618. [Google Scholar] [CrossRef]
- Everson, C.S.; Everson, T.M.; Tainton, N.M. The dynamics of Themeda triandra tillers in relation to burning in the Natal Drakensberg. J. Grass Soc. S. Afr. 1985, 2, 18–25. [Google Scholar] [CrossRef]
- Everson, T.M.; Yeaton, R.I.; Everson, C.S. Seed dynamics of Themeda triandra in the montane grasslands of South Africa. Afr. J. Range Forage Sci. 2009, 26, 19–26. [Google Scholar] [CrossRef]
- Morgan, J.W.; Lunt, I.D. Effects of time-since-fire on the tussock dynamics of a dominant grass (Themeda triandra) in a temperate Australian grassland. Biol. Conserv. 1999, 88, 379–386. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Beveridge, C.A. Apical dominance. Curr. Biol. 2017, 27, R864–R865. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ATB | TB | LB | TT | RT | CA | CD | HT | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOV | DF | F | p | F | p | F | p | F | p | F | p | F | p | F | p | F | p |
| Spp | 3 | 5.00 | 0.003 | 42.71 | <0.001 | 0.73 | <0.001 | 45.75 | <0.022 | 14.1 | <0.001 | 6.77 | <0.01 | 44.6 | <0.001 | 2.84 | <0.001 |
| FRI | 3 | 3.84 | 0.006 | 18.88 | <0.001 | 6.40 | 0.537 | 3.34 | <0.001 | 7.99 | <0.001 | 0.83 | 0.481 | 1.05 | 0.384 | 0.69 | 0.557 |
| Spp * FRI | 9 | 1.09 | 0.027 | 0.84 | 0.003 | 0.51 | 0.863 | 1.84 | 0.050 | 2.24 | 0.025 | 0.63 | 0.770 | 2.76 | 0.047 | 0.34 | 0.960 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mndela, M.; Moss, S.; Gusha, B.; Thamaga, K.H.; Afuye, G.A.; Abdo, H.G.; Almohamad, H. Functional Trait Responses of C4 Bunchgrasses to Fire Return Intervals in the Semi-Arid Savanna of South Africa. Diversity 2023, 15, 1201. https://doi.org/10.3390/d15121201
Mndela M, Moss S, Gusha B, Thamaga KH, Afuye GA, Abdo HG, Almohamad H. Functional Trait Responses of C4 Bunchgrasses to Fire Return Intervals in the Semi-Arid Savanna of South Africa. Diversity. 2023; 15(12):1201. https://doi.org/10.3390/d15121201
Chicago/Turabian StyleMndela, Mthunzi, Siphenamhla Moss, Bukho Gusha, Kgabo Humphrey Thamaga, Gbenga Abayomi Afuye, Hazem Ghassan Abdo, and Hussein Almohamad. 2023. "Functional Trait Responses of C4 Bunchgrasses to Fire Return Intervals in the Semi-Arid Savanna of South Africa" Diversity 15, no. 12: 1201. https://doi.org/10.3390/d15121201
APA StyleMndela, M., Moss, S., Gusha, B., Thamaga, K. H., Afuye, G. A., Abdo, H. G., & Almohamad, H. (2023). Functional Trait Responses of C4 Bunchgrasses to Fire Return Intervals in the Semi-Arid Savanna of South Africa. Diversity, 15(12), 1201. https://doi.org/10.3390/d15121201