

Fatty Acid Spectra in Mesopelagic Fishes of the Myctophidae and Stomiidae Families Collected in the North East Atlantic

 , ,
, ,  and
and 
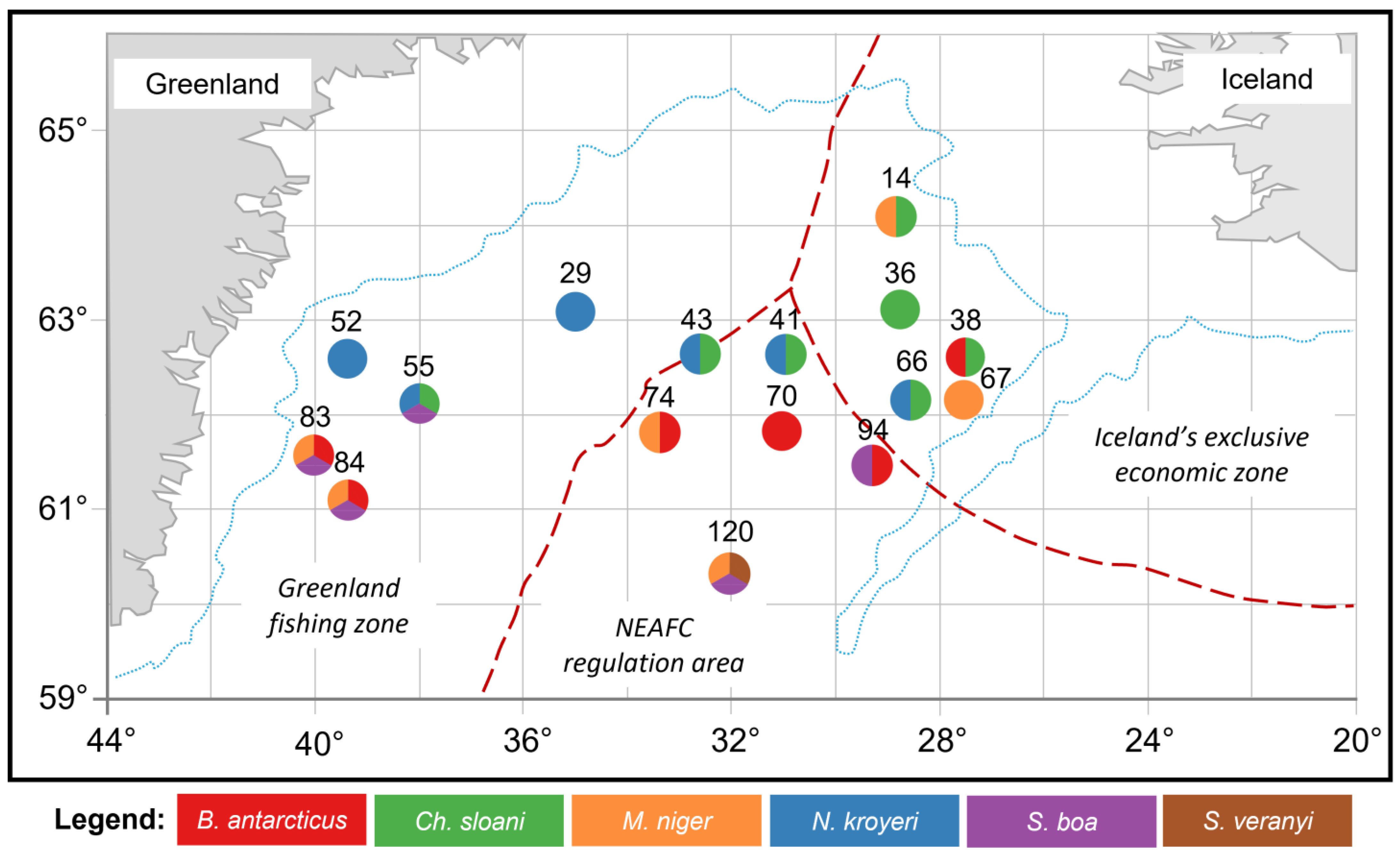{kind=link}
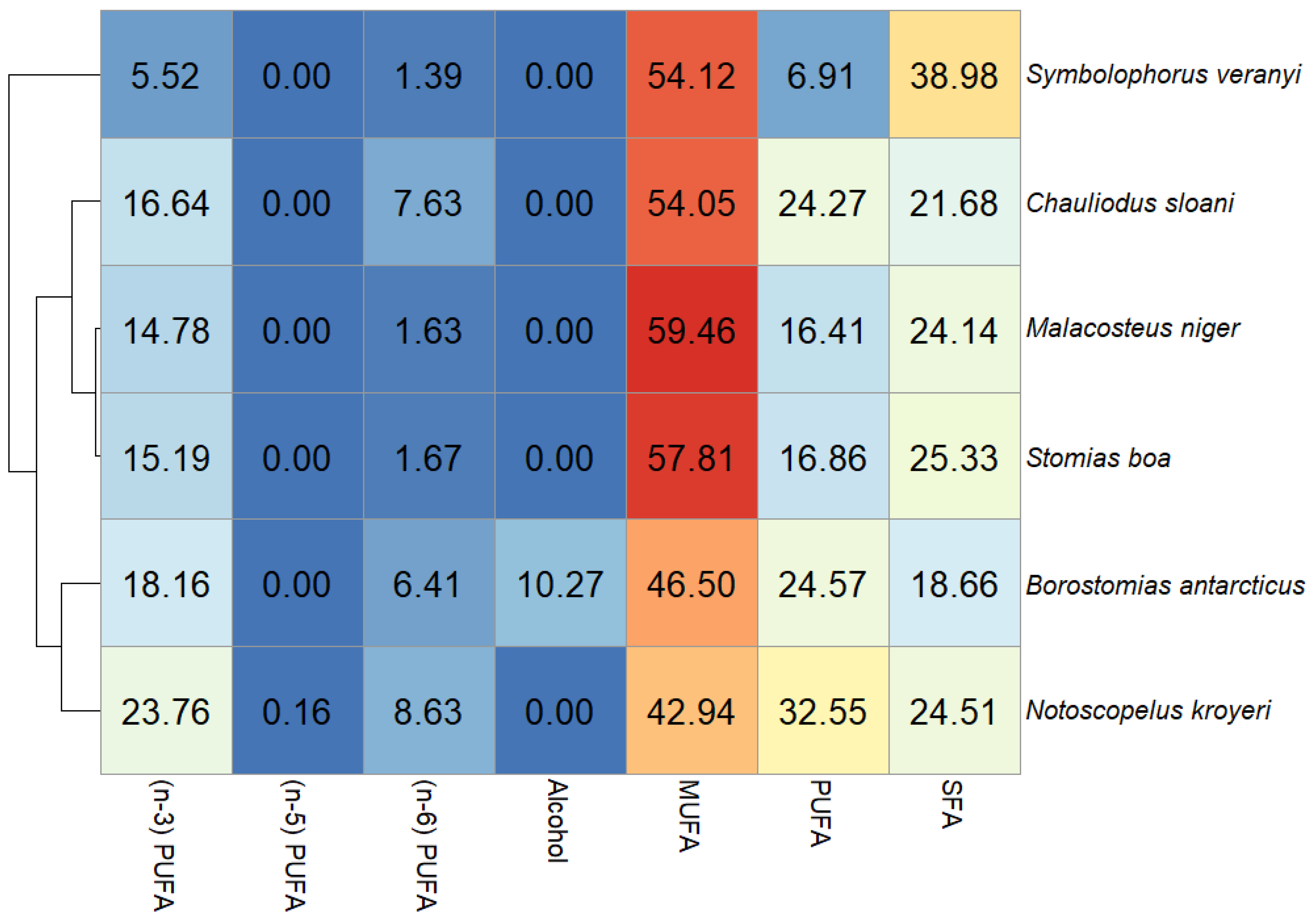{kind=link}
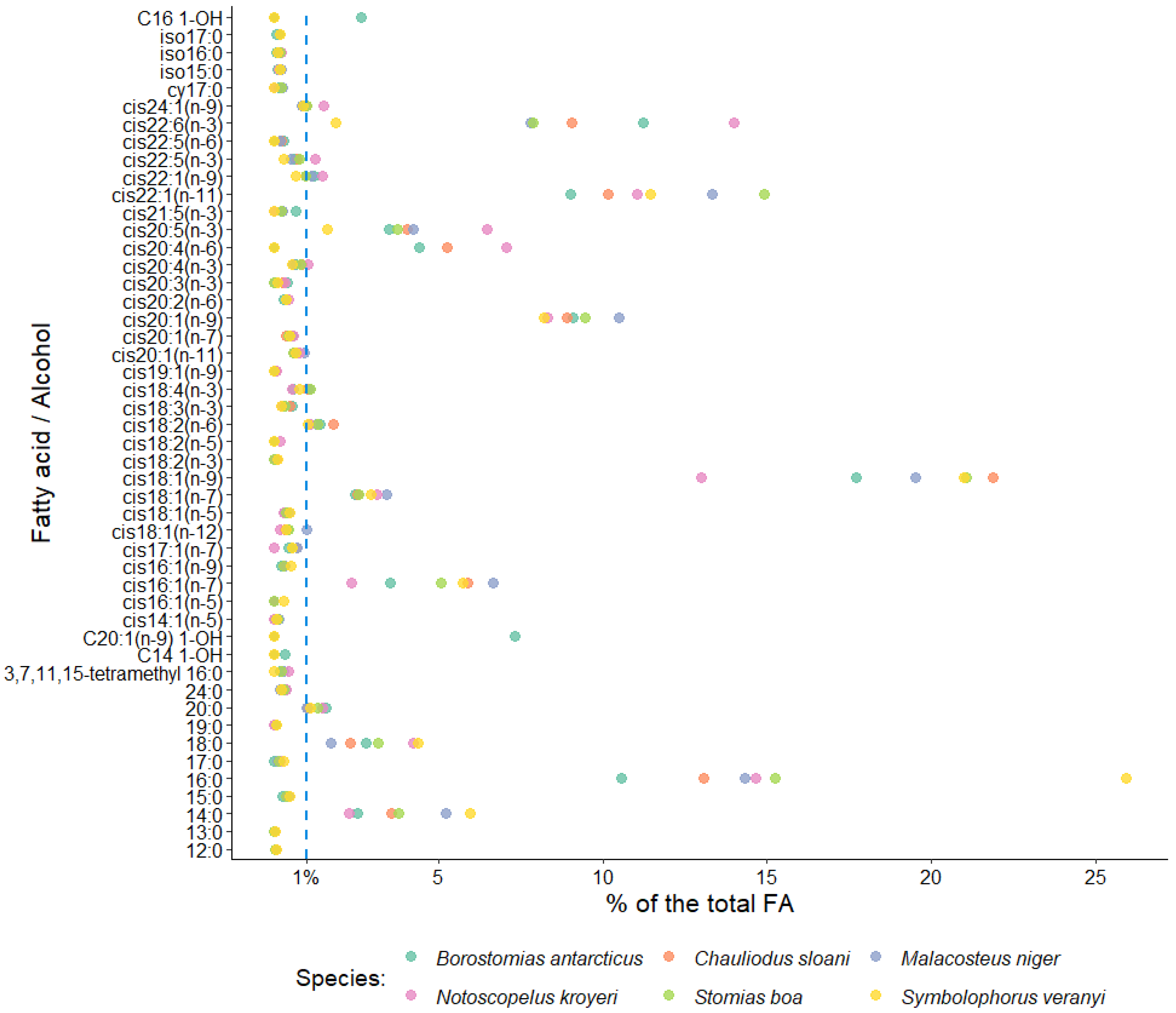{kind=link}
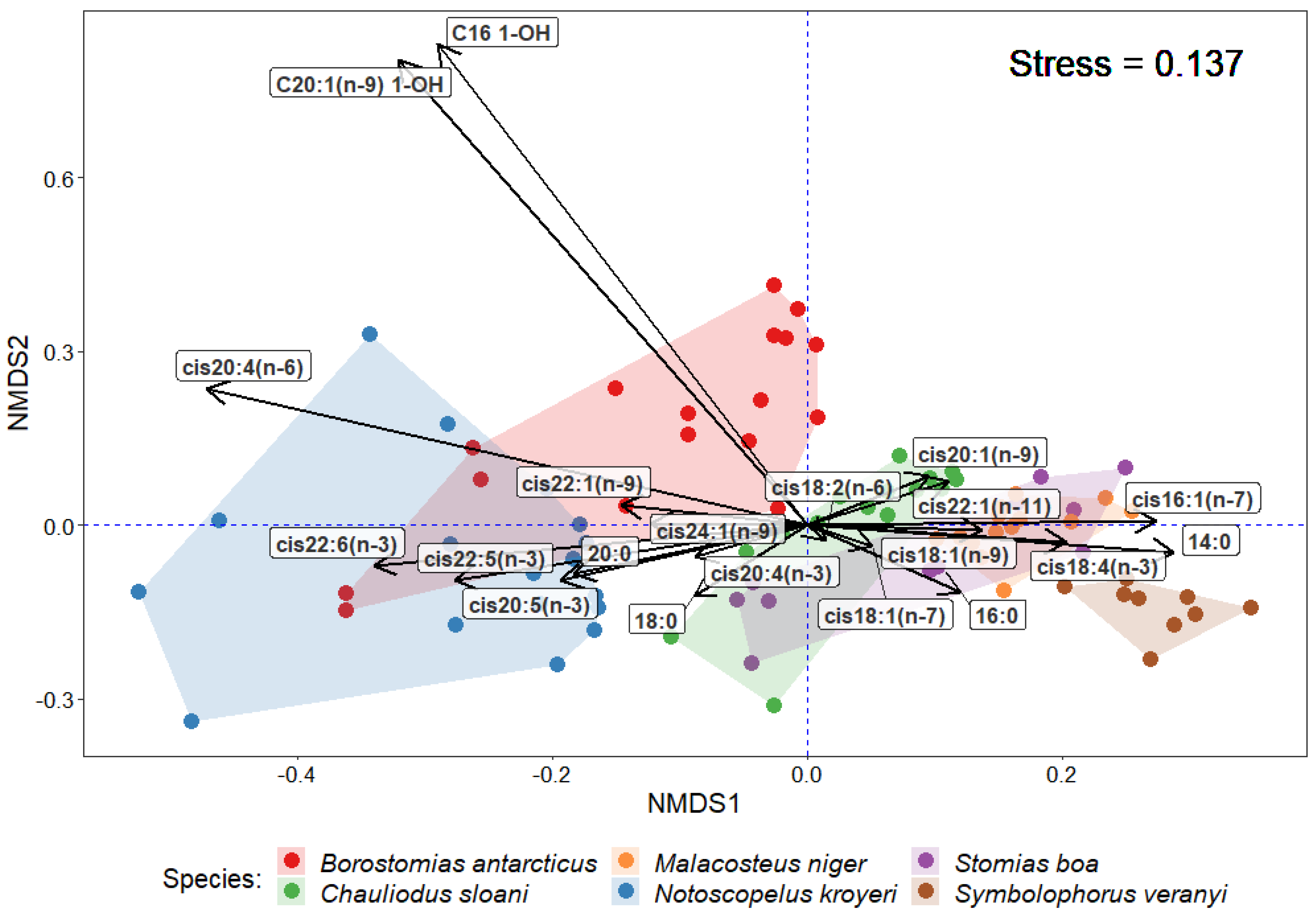{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Lipid Extraction
2.3. Gas Chromatography
2.3.1. Fatty Acid Analysis of the Total Lipids
2.3.2. Fatty Acid Analysis of the Individual Lipid Classes—Phospholipids, Triacylglycerols, and Cholesterol Esters and Waxes
2.4. Statistical Analysis
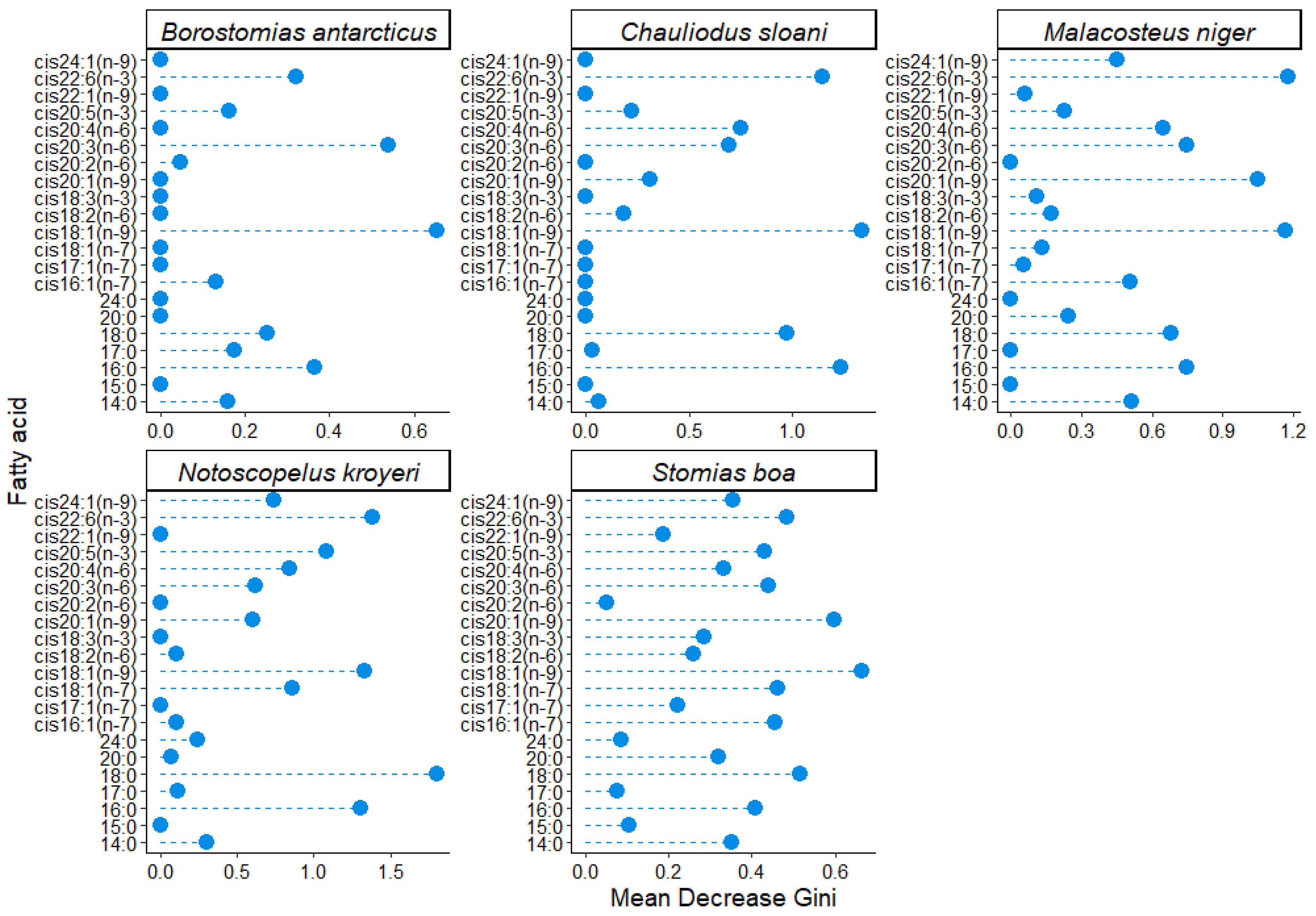
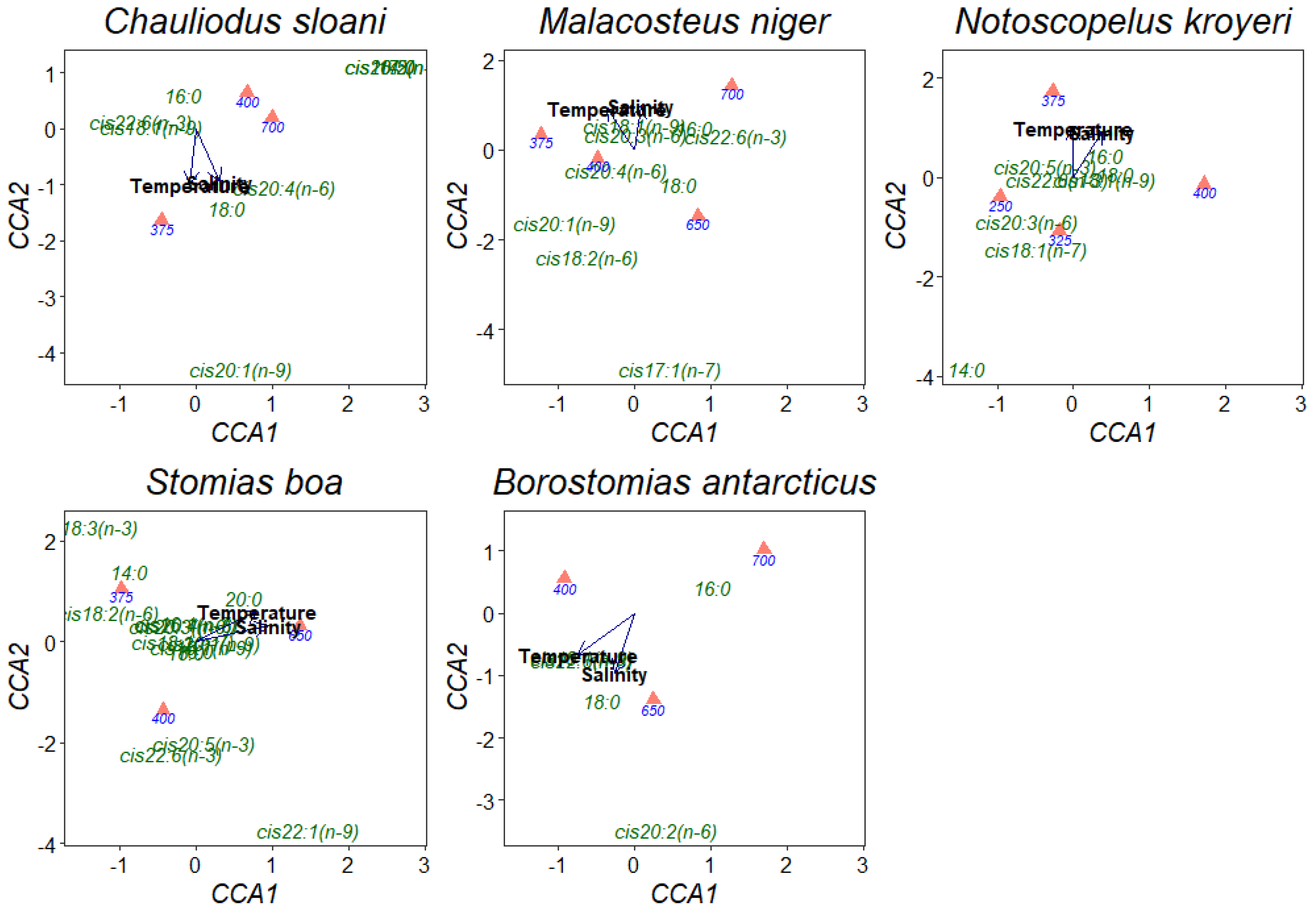
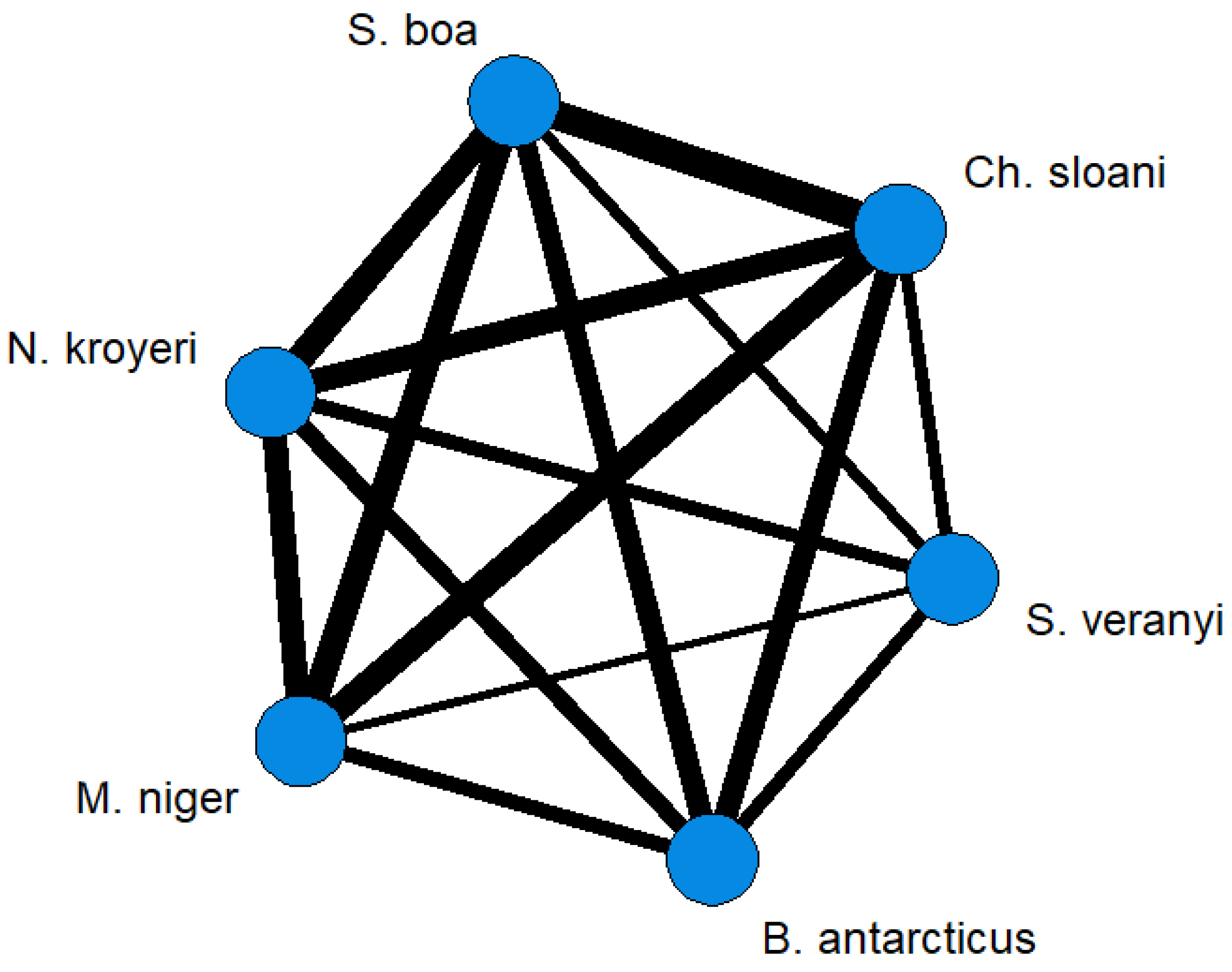
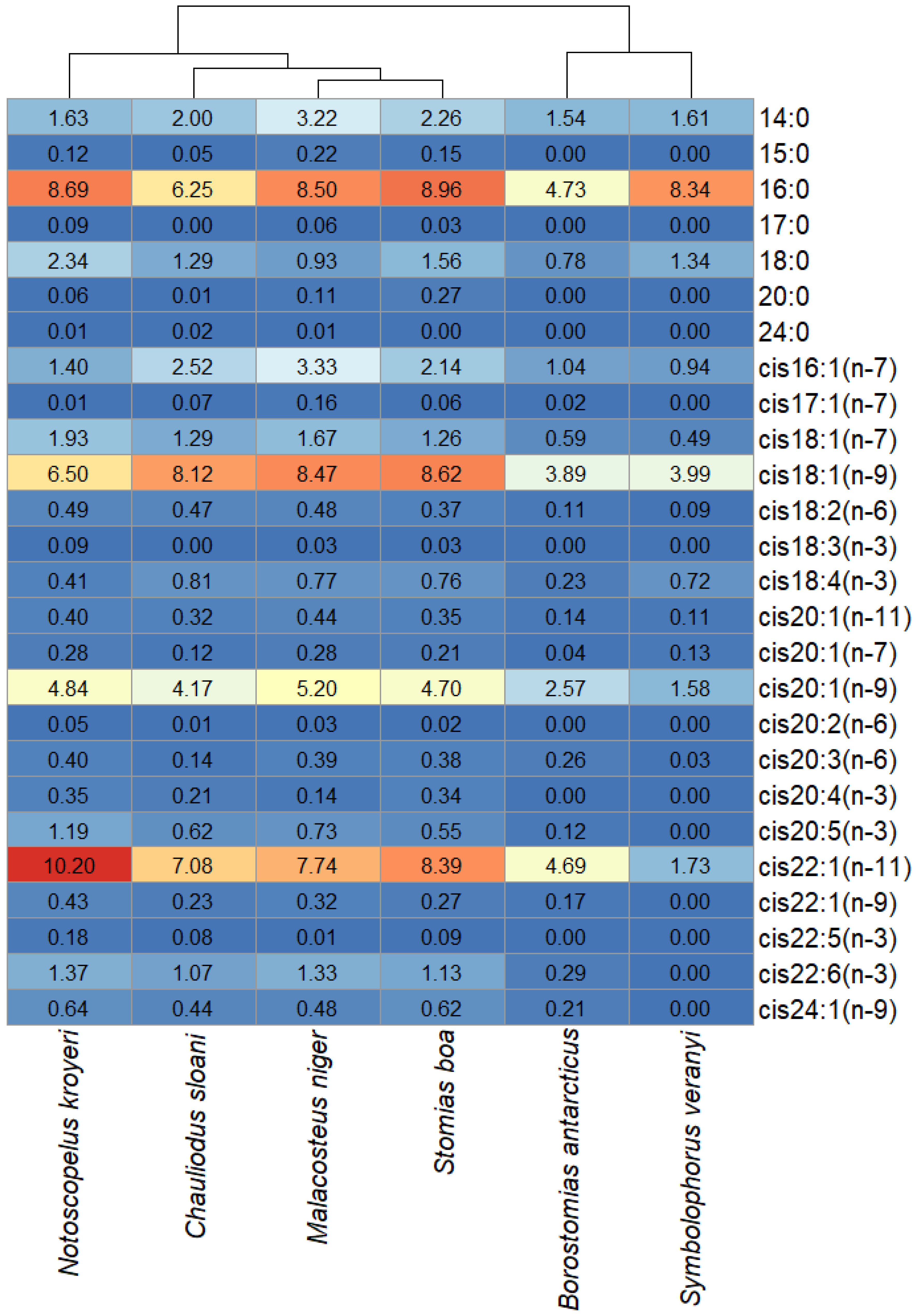
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Irigoien, X.; Klevjer, T.A.; Røstad, A.; Martinez, U.; Boyra, G.; Acuña, J.L.; Bode, A.; Echevarria, F.; Gonzalez-Gordillo, J.I.; Hernandez-Leon, S.; et al. Large mesopelagic fishes biomass and trophic efficiency in the open ocean. Nat. Commun. 2014, 5, 3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, M.; Browman, H.I. Developing the knowledge base needed to sustainably manage mesopelagic resources. ICES J. Mar. Sci. 2019, 76, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Aumont, O.; Maury, O.; Lefort, S.; Bopp, L. Evaluating the potential impacts of the diurnal vertical migration by marine organisms on marine biogeochemistry. Glob. Biogeochem. Cycles 2018, 32, 1622–1643. [Google Scholar] [CrossRef]
- Salvanes, A.G.V.; Kristoffersen, J.B. Mesopelagic Fishes; Academic Press: Cambridge, MA, USA, 2001; pp. 1711–1717. [Google Scholar]
- Catul, V.; Gauns, M.; Karuppasamy, P.K. A review on mesopelagic fishes belonging to family Myctophidae. Rev. Fish Biol. Fish 2011, 21, 339–354. [Google Scholar] [CrossRef]
- Tocher, D.R.; Bell, J.G.; Dick, J.R.; Henderson, R.J.; McGhee, F.; Michell, D.; Morris, P.C. Polyunsaturated fatty acid metabolism in Atlantic salmon (Salmo salar) undergoing parr-smolt transformation and the effects of dietary linseed and rapeseed oils. Fish Physiol. Biochem. 2000, 23, 59–73. [Google Scholar] [CrossRef]
- Arts, M.T.; Kohler, C.C. Health and conditions in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: Heidelberg, Germany; Dordrecht, The Netherlands; London, UK; New York, NY, USA, 2009; pp. 237–257. [Google Scholar]
- Shillito, B.; Desurmont, C.; Barthélémy, D.; Farabos, D.; Després, G.; Ravaux, J.; Zbinden, M.; Lamazière, A. Lipidome variations of deep-sea vent shrimps according to acclimation pressure: A homeoviscous response? Deep-Sea Res. Part I Oceanogr. Res. Pap. 2020, 161, 103285. [Google Scholar] [CrossRef]
- Kreps, E.M. Lipids of Cellular Membranes. Evolution of Brain Lipids. Adaptive Function of Lipids; Nauka: St. Petersburg, Russia, 1981; p. 339. [Google Scholar]
- Ashjian, C.J.; Campbell, R.G.; Welch, H.E.; Butler, M.; Van Keuren, D. Annual cycle in abundance, distribution, and size in relation to hydrography of important copepod species in the western Arctic Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 1235–1261. [Google Scholar] [CrossRef]
- Petursdottir, H.; Gislason, A.; Falk-Petersen, S. Lipid classes and fatty acid composition of muscle, liver and skull oil in deep-sea redfish Sebastes mentella over the Reykjanes Ridge. J. Fish Biol. 2008, 73, 2485–2496. [Google Scholar] [CrossRef]
- Petursdottir, H.; Gislason, A.; Falk-Petersen, S.; Hop, H.; Svavarsson, J. Trophic interaction of the pelagic ecosystem over the Reykjanes Ridge as evaluated by fatty acid and stable isotope analyses. Deep-Sea Research. Part II 2008, 55, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Graeve, M.; Kattner, G.; Piepenburg, D. Lipids in Arctic benthos: Does the fatty acid and alcohol composition reflect feeding and trophic interactions? Polar Biol. 1997, 18, 53–61. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Mayzaud, P.; Chevallier, J.; Tavernier, E.; Moteki, M.; Koubbi, P. Lipid composition of the Antarctic fish Pleuragramma antarcticum. Influence of age class. Polar Sci. 2011, 5, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Krutetskaya, Z.I.; Lebedev, O.E. Arachidonic acid and its products: Pathways of formation and metabolism in cells. Cell Tissue Biol. 1993, 35, 3–27. [Google Scholar]
- Ghioni, C.; Tocher, D.R.; Bell, M.V.; Dick, J.R.; Sargent, J.R. Low C18 to C20 fatty acid elongase activity and limited conversion of stearidonic acid, 18: 4 (n–3), to eicosapentaenoic acid, 20: 5 (n–3), in a cell line from the turbot, Scophthalmus maximus. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 1993, 1437, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Sergeeva, M.G.; Varfolomeeva, A.T. Arachidonic Acid Cascade; Public education: Moscow, Russia, 2006; p. 256. [Google Scholar]
- Boldyrev, A.A.; Kyayvaryainen, E.I.; Ilyukha, V.A. Biomembranology: A Textbook; KarRC RAS: Petrozavodsk, Russia, 2006; p. 226. [Google Scholar]
- Bell, M.V.; Dick, J.R.; Porter, A.E.A. Biosynthesis and tissue deposition of docosahexaenoic acid (22:6n−3) in rainbow trout (Oncorhynchus mykiss). Lipids 2001, 36, 1153–1159. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R.; Kelly, M.S. Biosynthesis of eicosapentaenoic acid in the sea urchin Psammechinus miliaris. Lipids 2001, 36, 79–82. [Google Scholar] [CrossRef]
- Monroig, Ó.; Rotllant, J.; Sánchez, E.; Cerdá-Reverter, J.M.; Tocher, D.R. Expression of long-chain polyunsaturated fatty acid (LC-PUFA) biosynthesis genes during zebrafish Danio rerio early embryogenesis. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2009, 1791, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Monroig, O.; Tocher, D.R.; Castro, L.F.C. Polyunsaturated fatty acid biosynthesis and metabolism in fish. In Polyunsaturated Fatty Acid Metabolism; AOCS Press: Urbana, IL, USA, 2018; pp. 31–60. [Google Scholar]
- Hazel, J.R. Thermal adaptation in biological membranes: Is homeoviscous adaptation the explanation? Annu. Rev. Physiol. 1995, 57, 19–42. [Google Scholar] [CrossRef]
- Hulbert, A.J. The under-appreciated fats of life: The two types of polyunsaturated fats. J. Exp. Biol. 2021, 224, jeb232538. [Google Scholar] [CrossRef]
- Biederman, A.M.; O’Brien, K.M.; Crockett, E.L. Homeoviscous adaptation occurs with thermal acclimation in biological membranes from heart and gill, but not the brain, in the Antarctic fish Notothenia coriiceps. J. Comp. Physiol. B 2021, 191, 289–300. [Google Scholar] [CrossRef]
- Macdonald, A. Life at High Pressure; Springer International Publishing: Cham, Switzerland, 2021; 445p. [Google Scholar]
- Olsen, R.E.; Strand, E.; Melle, W.; Nørstebø, J.T.; Lall, S.P.; Ringø, E.; Tocher, D.R.; Sprague, M. Can mesopelagic mixed layers be used as feed sources for salmon aquaculture? Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 180, 104722. [Google Scholar] [CrossRef]
- Yang, Z.H.; Emma-Okon, B.; Remaley, A.T. Dietary marine-derived long-chain monounsaturated fatty acids and cardiovascular disease risk: A mini review. Lipids Health Dis. 2016, 15, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eduardo, L.N.; Lucena-Frédou, F.; Mincarone, M.M.; Soares, A.; Loc’h, L.; Frédou, T.; Ménard, F.; Bertrand, A. Trophic ecology, habitat, and migratory behaviour of the viperfish Chauliodus sloani reveal a key mesopelagic player. Sci. Rep. 2020, 10, 20996. [Google Scholar] [CrossRef] [PubMed]
- Voronin, V.P.; Nemova, N.N.; Ruokolainen, T.R.; Artemenkov, D.V.; Rolskii, A.Y.; Orlov, A.M.; Murzina, S.A. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules 2021, 11, 704. [Google Scholar] [CrossRef]
- Voronin, V.P.; Artemenkov, D.V.; Orlov, A.M.; Murzina, S.A. Lipids and Fatty Acids in Some Mesopelagic Fish Species: General Characteristics and Peculiarities of Adaptive Response to Deep-Water Habitat. J. Mar. Sci. Eng. 2022, 10, 949. [Google Scholar] [CrossRef]
- Murzina, S.A.; Voronin, V.P.; Ruokolainen, T.R.; Artemenkov, D.V.; Orlov, A.M. Comparative Analysis of Lipids and Fatty Acids in Beaked Redfish Sebastes mentella Travin, 1951 Collected in Wild and in Commercial Products. J. Mar. Sci. Eng. 2022, 10, 59. [Google Scholar] [CrossRef]
- McGinnis, R.F. Biogeography of the lantern fishes (Myctophidae) south of 30°S. Antarct. Res Ser 1982, 35, 110. [Google Scholar]
- ICES. Manual for the International Deep Pelagic Ecosystem Survey in the Irminger Sea and Adjacent Waters. In Series of ICES Survey Protocols SISP 11- IDEEPS VI, Copenhagen, Denmark; 2015; 49p, Available online: https://ices-library.figshare.com/articles/report/SISP_11_-_Manual_for_the_International_Deep_Pelagic_Ecosystem_Survey_in_the_Irminger_Sea_and_Adjacent_Waters_Version_1/19050851 (accessed on 15 December 2022).
- Kukuev, E.I.; Gushchin, A.V.; Gomolitsky, V.D. Methodical Materials for the Identification of Fish in the Open Waters of the North Atlantic; AtlantNIRO: Kaliningrad, Russia, 1980; 145p. [Google Scholar]
- Dolgov, A.V. Atlas-Key to Fishes of the Barents Sea; PINRO: Murmansk, Russia, 2011; 187p. [Google Scholar]
- Schilling, B.; Couperus, A.S.; Bos, O.G. Fotogids Mesopelagische Vissen: Noordoost Atlantische Oceaan = Photo Guide Mesopelagic Fish: North East Atlantic Ocean; Wageningen Marine Research: Yerseke, The Netherlands, 2019; 121p. [Google Scholar]
- Sutton, T.T.; Hulley, P.A.; Wienerroither, R.; Zaera-Perez, D.; Paxton, J.R. Identification Guide to the Mesopelagic Fishes of the Central and South East Atlantic Ocean; FAO: Rome, Italy, 2020. [Google Scholar]
- Folch, J.; Lees, M.; Sloan-Syanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue (for brain, liver and muscle). J. Biol. Chem 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Cyganov, E.P. Method for direct lipid methylation after TLC without elution from silica gel. Labor. Matter 1971, 8, 490–493. [Google Scholar]
- Murzina, S.A.; Pekkoeva, S.N.; Kondakova, E.A.; Nefedova, Z.A.; Filippova, K.A.; Nemova, N.N.; Orlov, A.M.; Berge, J.; Falk-Petersen, S. Tiny but Fatty: Lipids and Fatty Acids in the Daubed Shanny (Leptoclinus maculatus), a Small Fish in Svalbard Waters. Biomolecules 2020, 10, 368. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double development HPTLC and scanning densitometry. J. Exp. Mar. Biol. Ecol 1989, 129, 189–197. [Google Scholar] [CrossRef]
- ISO/TS 17764-2-2015; Feeds, Compound Feeds. Determination of the Content of Fatty Acids. Part 2. Gas Chromatographic Method. Russia. ISO: Geneva, Switzerland, 2017; 16p.
- Kabakoff, R. R in Action: Data Analysis and Graphics with R. TRANSL. from English P. A. Volkova; DMK Press: Moscow, Russia, 2014; p. 588. [Google Scholar]
- Shitikov, V.K.; Mastitsky, S.E. Classification, Regression, Data Mining Algorithms Using R. 2017. Available online: https://github.com/ranalytics/data-mining (accessed on 1 December 2022).
- Bruce, P.; Bruce, A. Practical Statistics for Data Scientists; Spb. BHV-Peterburg: Saint Petersburg, Russia, 2020; 304p. [Google Scholar]
- Lantz, B. Machine Learning with R: Expert Techniques for Predictive Modeling; Spb.: Saint Petersburg, Russia, 2020; 464p. [Google Scholar]
- FAO 2020. The Mesopelagic Fish Guide: Shedding Light on 550 Fish Species in One of the Largest Ecosystems on Earth. EAF—Nansen Programme. Available online: http://www.fao.org/in-action/eaf-nansen/news-events/detail-events/en/c/1311820/ (accessed on 10 December 2022).
- Choy, C.A.; Popp, B.N.; Hannides, C.C.; Drazen, J.C. Trophic structure and food resources of epipelagic and mesopelagic fishes in the North Pacific Subtropical Gyre ecosystem inferred from nitrogen isotopic compositions. Limnol. Oceanogr. 2015, 60, 1156–1171. [Google Scholar] [CrossRef]
- Cavallaro, M.; Ammendolia, G.; Andaloro, F.; Battaglia, P. First record of the mesopelagic fish Diaphus dumerilii (Bleeker, 1856) in the Mediterranean Sea. Mar. Biodivers. 2017, 47, 585–588. [Google Scholar] [CrossRef]
- Choy, C.A.; Portner, E.; Iwane, M.; Drazen, J.C. Diets of five important predatory mesopelagic fishes of the central North Pacific. Mar. Ecol. Prog. Ser. 2013, 492, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Olaso, I.; Velasco, F.; Sánchez, F.; Serrano, A.; Rodríguez-Cabello, C.; Cendrero, O. Trophic relations of lesser-spotted catshark (Scyliorhinus canicula) and blackmouth catshark (Galeus melastomus) in the Cantabrian Sea. J. Northwest Atl. Fish. Sci. 2005, 35, 481–494. [Google Scholar] [CrossRef]
- Pusineri, C.; Chancollon, O.; Ringelstein, J.; Ridoux, V. Feeding niche segregation among the Northeast Atlantic community of oceanic top predators. Mar. Ecol. Prog. Ser. 2008, 361, 21–34. [Google Scholar] [CrossRef]
- Olivar, M.P.; Hulley, P.A.; Castellón, A.; Emelianov, M.; López, C.; Tuset, V.M.; Contreras, T.; Molí, B. Mesopelagic fishes across the tropical and equatorial Atlantic: Biogeographical and vertical patterns. Prog. Oceanogr. 2017, 151, 116–137. [Google Scholar] [CrossRef]
- Duhamel, G.; Hulley, P.A.; Causse, R.; Koubbi, P.; Vacchi, M.; Pruvost, P.; Vigetta, S.; Irisson, J.O.; Mormede, S.; Belchier, M.; et al. Biogeographic Patterns of Fish. Biogeographic Atlas of the Southern Ocean; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 328–362. [Google Scholar]
- Iverson, S.J. Tracing aquatic food webs using fatty acids: From qualitative indicators to quantitative determination. In Lipids in Aquatic Ecosystems, 3rd ed.; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 281–308. [Google Scholar]
- Ozdemir, N.S.; Parrish, C.C.; Parzanini, C.; Mercier, A. Neutral and polar lipid fatty acids in five families of demersal and pelagic fish from the deep Northwest Atlantic. ICES J. Mar. Sci. 2019, 76, 1807–1815. [Google Scholar] [CrossRef]
- Saito, H.; Murata, M. Origin of the monoene fats in the lipid of midwater fishes: Relationship between the lipids of myctophids and those of their prey. Mar. Ecol. Prog. Ser 1998, 168, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Lea, M.A.; Nichols, P.D.; Wilson, G. Fatty acid composition of lipid-rich myctophids and mackerel icefish (Champsocephalus gunnari)–Southern Ocean food-web implications. Polar Biol. 2002, 25, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Falk-Petersen, S.; Hagen, W.; Kattner, G.; Clarke, A.; Sargent, J.R. Lipids, trophic relationships, and biodiversity in Arctic and Antarctic krill. Can. J. Fish. Aq. Sci 2000, 57, 178–191. [Google Scholar] [CrossRef]
- Dalsgaard, J.; St. John, M.; Kattner, G.; Muller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar]
- Stegeman, J.J.; Schlezinger, J.J.; Craddock, J.E.; Tillitt, D.E. Cytochrome P450 1A expression in midwater fishes: Potential effects of chemical contaminants in remote oceanic zones. Environ. Sci. Technol. 2001, 35, 54–62. [Google Scholar] [CrossRef]
- Sutton, T.T. Trophic ecology of the deep-sea fish Malacosteus niger (Pisces: Stomiidae): An enigmatic feeding ecology to facilitate a unique visual system? Deep Sea Res. Part I: Oceanogr. Res. Pap. 2005, 52, 2065–2076. [Google Scholar] [CrossRef]
- Munschy, C.; Spitz, J.; Bely, N.; Héas-Moisan, K.; Olivier, N.; Pollono, C.; Chouvelon, T. A large diversity of organohalogen contaminants reach the meso-and bathypelagic organisms in the Bay of Biscay (northeast Atlantic). Mar. Pollut. Bull. 2022, 184, 114180. [Google Scholar] [CrossRef] [PubMed]
- Klimpel, S.; Palm, H.W.; Busch, M.W.; Kellermanns, E.; Rückert, S. Fish parasites in the Arctic deep-sea: Poor diversity in pelagic fish species vs. heavy parasite load in a demersal fish. Deep Sea Res. Part I Oceanogr. Res. Pap. 2006, 53, 1167–1181. [Google Scholar] [CrossRef]
- Butler, M.; Bollens, S.M.; Burkhalter, B.; Madin, L.P.; Horgan, E. Mesopelagic fishes of the Arabian Sea: Distribution, abundance and diet of Chauliodus pammelas, Chauliodus sloani, Stomias affinis, and Stomias nebulosus. Deep Sea Res. II 2001, 48, 1369–1383. [Google Scholar] [CrossRef]
- Sargent, J.R.; Falk-Petersen, S. The lipid biochemistry of calanoid copepods. Hydrobiologia 1988, 167/168, 101–114. [Google Scholar] [CrossRef]
- Scott, C.L.; Kwasniewski, S.; Falk-Petersen, S.; Sargent, J.R. Species differences, origins andfunctions of fatty alcohols and fatty acids in the wax esters and phospholipids of Calanus hyperboreus, C. glacialis and C. finmarchicus from Arctic waters. Mar. Ecol. Prog. Ser. 2002, 235, 127–134. [Google Scholar] [CrossRef]
- Lee, R.F. Lipid composition of the copepod Calanus hyperboreas from the ArcticOcean. Changes with depth and season. Mar. Biol. 1974, 26, 313–318. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Mayzaud, P.; Kattner, G.; Sargent, J.R. Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 2009, 5, 18–39. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, N.M.; Wanders, R.J.A.; Saudubray, J.M.; Jakobs, C. The metabolism of phytanic acid and pristanic acid in man: A review. J. Inherit. Metab. Dis. 1998, 21, 697–728. [Google Scholar] [CrossRef] [PubMed]
- Bode, M.; Hagen, W.; Schukat, A.; Teuber, L.; Fonseca-Batista, D.; Dehairs, F.; Auel, H. Feeding strategies of tropical and subtropical calanoid copepods throughout the eastern Atlantic Ocean–Latitudinal and bathymetric aspects. Prog. Oceanogr. 2015, 138, 268–282. [Google Scholar] [CrossRef]
- Gershanovich, A.D. Lipid mobilization during early development of surgeons. In Proc. First Intarnational Symposium Sturgeon; Bordeaux: Gironde, France, 1991; pp. 41–52. [Google Scholar]
- Atlasova, M. Estimates of the total food composition Notoscopelus kroyeri in the Irminger Sea in 2003–2013 years. Nat. Math. Sci. Mod. World 2015, 32, 54–58. [Google Scholar]
- Auel, H.; Harjes, M.; Da Rocha, R.; Stübing, D.; Hagen, W. Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipods Themisto abyssorum and T. libellula. Polar Biol. 2002, 25, 374–383. [Google Scholar] [CrossRef]
- Schukat, A.; Auel, H.; Teuber, L.; Lahajnar, N.; Hagen, W. Complex trophic interactions of calanoid copepods in the Benguela upwelling system. J. Sea Res. 2014, 85, 186–196. [Google Scholar] [CrossRef]
- Gaskett, A.C.; Bulman, C.; He, X.; Goldsworthy, S.D. Diet composition and guild structure of mesopelagic and bathypelagic fishes near Macquarie Island, Australia. N. Z. J. Mar. Freshw. Res 2001, 35, 469–476. [Google Scholar] [CrossRef]
- Rabinovich, A.L.; Ripatti, P.O. Polyunsaturated carbon chins of lipids: Structure, properties, functions. Biol. Bull. Rev 1994, 114, 581–594. [Google Scholar]
- Shulman, G.E.; Yuneva, T.V. Role of docosahexaenoic acid in adaptations fishes (review). Hydrobiol. J. 1990, 26, 43–51. [Google Scholar]
- Isanta Navarro, J.; Fromherz, M.; Dietz, M.; Zeis, B.; Schwarzenberger, A.; Martin--Creuzburg, D. Dietary polyunsaturated fatty acid supply improves Daphnia performance at fluctuating temperatures, simulating diel vertical migration. Freshw. Biol. 2019, 64, 1859–1866. [Google Scholar] [CrossRef]
- Murzina, S.A.; Nefedova, Z.A.; Pekkoeva, S.N.; Veselov, A.E.; Baryshev, I.A.; Ripatti, P.O.; Nemova, N.N. Content of fatty acids in forage objects of juveniles of salmonids from rivers of the Lake Onega basin. Inland Water Biol. 2019, 12, 96–103. [Google Scholar] [CrossRef]
- Almansa, E.; Martian, M.V.; Cejas, J.R.; Badi, P.; Jerez, S.; Lorenzo, A. Lipid and fatty acid composition of female gilthead seabream during their reproductive cycle: Effects of a diet lacking n--3 HUFA. J. Fish Biol. 2001, 59, 267–286. [Google Scholar] [CrossRef]
- Buda, C.; Dey, I.; Balogh, N.; Horvath, L.I.; Maderspach, K.; Juhasz, M.; Yeo, Y.K.; Farkas, T. Structural order of membranes and composition of phospholipids in fish brain cells during thermal acclimatization. Proc. Natl. Acad. Sci. USA 1994, 91, 8234–8238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemova, N.N.; Nefedova, Z.A.; Murzina, S.A.; Veselov, A.E.; Ripatti, P.O. Comparative characteristics of the lipid and fatty acid status of eyed-stage atlantic salmon embryos reared in natural and artificial environments. Biol. Bull. 2015, 42, 493–499. [Google Scholar] [CrossRef]
- Murzina, S.A.; Nefedova, Z.A.; Veselov, A.E.; Ripatti, P.O.; Nemova, N.N.; Pavlov, D.S. Changes in fatty acid composition during embryogenesis and in young age groups (0+) of Atlantic Salmon Salmo salar L.: The role of rheotactic behavior and lipid composition of fry in the formation of phenotypic groups of Salmon in large arctic rivers. In Salmon: Biology, Ecological Impacts and Economic Importance, 2nd ed.; Woo, P.T.K., Noakes, D.J., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2014; pp. 47–65. [Google Scholar]
- Winnikoff, J.R.; Haddock, S.H.; Budin, I. Depth-and temperature-specific fatty acid adaptations in ctenophores from extreme habitats. J. Exp. Biol. 2021, 224, jeb242800. [Google Scholar] [CrossRef]
- Neighbors, M.A. Triacylglycerols and wax esters in the lipids of deep midwater teleost fishes of the Southern California Bright. Mar. Biol. 1988, 98, 15–22. [Google Scholar] [CrossRef]
- Phleger, C.F.; Nelson, M.M.; Mooney, B.D.; Nichols, P.D. Wax esters versus triacylglycerols in myctophid fishes from the Southern Ocean. Antarct. Sci. 1999, 11, 436–444. [Google Scholar] [CrossRef]
- Roe, H.S.J.; Badcock, J. The diel migrations and distributions within a mesopelagic community in the North East Atlantic. 5. Vertical migrations and feeding of fish. Prog. Oceanogr. 1984, 13, 389–424. [Google Scholar] [CrossRef]
- Podrazhanskaya, S.G. Feeding habits of mesopelagic species of fish and estimation of plankton graze in the Northwest Atlantic. NAFO Sci. Counc. Stud. 1993, 19, 79–85. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronin, V.P.; Artemenkov, D.V.; Orlov, A.M.; Murzina, S.A. Fatty Acid Spectra in Mesopelagic Fishes of the Myctophidae and Stomiidae Families Collected in the North East Atlantic. Diversity 2023, 15, 166. https://doi.org/10.3390/d15020166
Voronin VP, Artemenkov DV, Orlov AM, Murzina SA. Fatty Acid Spectra in Mesopelagic Fishes of the Myctophidae and Stomiidae Families Collected in the North East Atlantic. Diversity. 2023; 15(2):166. https://doi.org/10.3390/d15020166
Chicago/Turabian StyleVoronin, Viktor P., Dmitrii V. Artemenkov, Alexei M. Orlov, and Svetlana A. Murzina. 2023. "Fatty Acid Spectra in Mesopelagic Fishes of the Myctophidae and Stomiidae Families Collected in the North East Atlantic" Diversity 15, no. 2: 166. https://doi.org/10.3390/d15020166
APA StyleVoronin, V. P., Artemenkov, D. V., Orlov, A. M., & Murzina, S. A. (2023). Fatty Acid Spectra in Mesopelagic Fishes of the Myctophidae and Stomiidae Families Collected in the North East Atlantic. Diversity, 15(2), 166. https://doi.org/10.3390/d15020166