Amphibians are represented in Rwanda by at least 61 anuran and one caecilian species. Some additional candidate species have already been identified by us but are not treated in the following accounts. First, we list the 62 species with validated country records, and then we give an account of species that have been assumed to occur in Rwanda by several authors but have never actually been collected within the borders of the country and are therefore rejected.
3.1. Species with Validated Country Record
The anuran species of Rwanda belong to ten families: (1)
Arthroleptidae Mivart, 1869 with three genera,
Arthroleptis Smith, 1849,
Cardioglossa Boulenger, 1900, and
Leptopelis Günther, 1859; (2)
Bufonidae Gray, 1825 with two genera,
Schismaderma Smith, 1849 and
Sclerophrys Tschudi, 1838; (3)
Dicroglossidae Anderson, 1871 with the genus
Hoplobatrachus Peters, 1863; (4)
Hyperoliidae Laurent, 1943 with five genera,
Afrixalus Laurent, 1944,
Callixalus Laurent, 1950,
Hylambates Duméril, 1853,
Hyperolius Rapp, 1842, and
Kassina Girard, 1853; (5)
Microhylidae Günther, 1858 (1843) with the genus
Phrynomantis Peters, 1867; (6)
Phrynobatrachidae Laurent, 1941 with the genus
Phrynobatrachus Günther, 1862; (7)
Pipidae Gray, 1825 with the genus
Xenopus Wagler, 1827; (8)
Ptychadenidae Dubois, 1987 with the genus
Ptychadena Boulenger, 1917; (9)
Pyxicephalidae Bonaparte, 1850 with two genera,
Amietia Dubois, 1987 and
Cacosternum Boulenger, 1887; (10)
Ranidae Batsch, 1796 with the genus
Amnirana Dubois, 1992 (
Table 1). The only caecilian species of Rwanda belongs to the genus
Boulengerula Tornier, 1896 in the family
Herpelidae Laurent, 1984 (
Table 1). The species are treated in the order of the families and genera, and then alphabetically within the genus to which they belong.
Key to the families and genera of the amphibians of Rwanda based on morphological characters:
- 1
Limbless........................... Herpelidae (only Boulengerula; only B. fischeri)
- 1′
With limbs ...................................................................................... 2
- 2
Toes with claw-like tips ........................................ Pipidae (only Xenopus)
- 2′
Toes without claw-like tips ................................................................. 3
- 3
Dorsum black with bright red dorsolateral bands and red spots on limbs ................................. Microhylidae (only Phrynomantis; only P. bifasciatus)
- 3′
Dorsal colour and pattern different ...................................................... 4
- 4
Distinct pale transverse groove behind the eyes, tooth-like projections in the upper and lower jaw .... Dicroglossidae (only Hoplobatrachus; only H. occipitalis)
- 4′
Transverse groove behind eyes and tooth-like projections in jaw absent ...... 5
- 5
Dorsum with three to four pairs of continuous longitudinal dermal ridges .................................................................. Ptychadenidae (only Ptychadena)
- 5′
Dorsum without or with one pair of continuous ridges, sometimes with additional interrupted ridges ..................................................................... 6
- 6
Mid-tarsal tubercle present ....... Phrynobatrachidae (only Phrynobatrachus)
- 6′
Mid-tarsal tubercle absent .................................................................. 7
- 7
Dermal fold extending along the tarsus to inner metatarsal tubercle ..................................................................................................... Bufonidae (8)
- 7′
Dermal fold along tarsus absent .......................................................... 9
- 8
Large paratoid gland present behind eye ...... Bufonidae (part.): Sclerophrys
- 8′
Paratoid glands absent ...... Bufonidae (part.): Schismaderma (only S. carens)
- 9
Venter white with large black spots ......................................................................................... Pyxicephalidae (part.): Cacosternum (only C. plimptoni)
- 9′
Venter without large black spots ........................................................ 10
- 10
Dorsolateral ridge extending from posterior end of upper eyelid, separated from well-developed supratympanic fold .......... Pyxicephalidae (part.): Amietia
- 10′
Dorsolateral ridge absent or extending from supratympanic fold ............. 11
- 11
Webbing between toes absent ............................................................ 12
- 11′
Webbing between toes present ........................................................... 13
- 12
Narrow vertebral dermal ridge on dorsum, dorsal colouration brown or grey with or without symmetric darker brown markings ......................................................................................................... Arthroleptidae (part.): Arthroleptis
- 12′
Vertebral ridge absent, dorsal colour bluish grey with irregularly shaped black blotches ............. Arthroleptidae (part.): Cardioglossa (only C. cyaneospila)
- 13
Conspicuous dorsolateral ridge running from eye as anterior part of supratympanic fold, then continuing straight-lined to end of body, tympanum as large or nearly as large as eye ..................................... Ranidae (only Amnirana)
- 13′
Dorsolateral ridge absent or reaching at most to middle of trunk, tympanum smaller................................................................................................... 14
- 14
Tympanum concealed or reduced ...................................................... 15
- 14′
Tympanum visible ........................................................................... 17
- 15
Pupil horizontal ..................................... Hyperoliidae (part.): Hyperolius
- 15′
Pupil vertical ................................................................................. 16
- 16
Dorsal colouration light brown to yellowish brown with darker brown markings ....................................................... Hyperoliidae (part.): Afrixalus
- 16′
Dorsal colouration chocolate brown with yellow blotches ........................................................................ Hyperoliidae (part.): Callixalus (only C. pictus)
- 17
Dorsal skin tuberculate, sides of thigh with conspicuous black and yellow transverse bars ................ Hyperoliidae (part.): Hylambates (only H. verrucosus)
- 17′
Dorsal skin smooth or shagreen, colouration of thighs different .............. 18
- 18
Canthus rostralis and supratympanic fold indistinct, dorsal colour yellowish brown to cream with contrasting large black stripes and blotches ................................................................. Hyperoliidae (part.): Kassina (only K. senegalensis)
- 18′
Canthus rostralis and supratympanic fold distinctly visible, dorsal colour brown or green without contrasting pattern, dorsal skin shagreen ................................................................................. Arthroleptidae (part.): Leptopelis
Eight arthroleptid species have been recorded from Rwanda. While the morphological features of the genera Arthroleptis, Cardioglossa and Leptopelis allow for a reliable assignment of species to a genus, species within each genus are not always readily distinguished from each other. Yet, species are well differentiated with respect to partial 16S sequences. The advertisement call (unknown for C. cyaneospila and L. cf. cynnamomeus) allows for a reliable assignment of reproductive males to species of Arthroleptis and Leptopelis in the field.
Key to the species of Arthroleptis of Rwanda based on morphological characters:
- 1
Large, SVL of males > 27 mm, females > 37 mm ................ A. adolfifriederici
- 1′
Small, SVL of males and females < 24 mm .............................. A. schubotzi
Key to the species of Leptopelis of Rwanda based on morphological characters:
- 1
Thick-bodied, toes webbed only at the bases ................................ L. bocagii
- 1′
Body sturdy but not thick, toes webbed to one-third ................................ 2
- 2
Canthus rostralis and supratympanic fold sharply defined, the latter continued as a short dorsolateral ridge .............................................. L. cf. cynnamomeus
- 2′
Canthus rostralis and supratympanic fold distinct but without a sharp edge, dorsolateral ridge absent ........................................................................... 3
- 3
Males with a white ventral side including the vocal sac ............. L. kivuensis
- 3′
Males with a bluish ventral side including the vocal sac ...... L. karissimbensis
Key to the arthroleptid species of Rwanda based on advertisement-call features:
- 1
Call tonal ................................................. Arthroleptis adolfifriederici
- 1′
Call pulsed .................................................................................... 2
- 2
Dominant frequency < 3000 Hz .......................................... Leptopelis (3)
- 2′
Dominant frequency > 4000 Hz ............................... Arthroleptis schubotzi
- 3
Dominant frequency < 1000 Hz .................................... Leptopelis bocagii
- 3′
Dominant frequency > 1000 Hz ........................................................... 4
- 4
Dominant frequency 1000–1500 Hz ..................... Leptopelis karissimbensis
- 4′
Dominant frequency > 1500 Hz .................................. Leptopelis kivuensis
Figure 2.
Rwandan species of Arthroleptidae (A) Arthroleptis adolfifriedrici, Gishwati-Mukura NP (photo: J.M.D.), (B) Arthroleptis schubotzi, Nyungwe NP (photo: J.M.D.), (C) Cardioglossa cyaneospila, Nyungwe NP (photo: Protais Niyigaba), (D) Leptopelis bocagii, Akagera NP (photo: J.M.D.), (E) Leptopelis cf. cynnamomeus, Cyamudongo Forest (photo: Eberhard Fischer), (F) Leptopelis karissimbensis, Nyungwe NP (photo: J.M.D.), (G) Leptopelis kivuensis, Nyungwe NP (photo: J.M.D.); and Dicroglossidae (H) Hoplobatrachus occipitalis, Bugarama (photo: J.M.D.).
Figure 2.
Rwandan species of Arthroleptidae (A) Arthroleptis adolfifriedrici, Gishwati-Mukura NP (photo: J.M.D.), (B) Arthroleptis schubotzi, Nyungwe NP (photo: J.M.D.), (C) Cardioglossa cyaneospila, Nyungwe NP (photo: Protais Niyigaba), (D) Leptopelis bocagii, Akagera NP (photo: J.M.D.), (E) Leptopelis cf. cynnamomeus, Cyamudongo Forest (photo: Eberhard Fischer), (F) Leptopelis karissimbensis, Nyungwe NP (photo: J.M.D.), (G) Leptopelis kivuensis, Nyungwe NP (photo: J.M.D.); and Dicroglossidae (H) Hoplobatrachus occipitalis, Bugarama (photo: J.M.D.).
Figure 3.
Advertisement calls of Rwandan Arthroleptidae (A) Arthroleptis adolfifriederici, (B) Arthroleptis schubotzi, (C) Leptopelis bocagii, (D) Leptopelis karissimbensis, (E) Leptopelis kivuensis; and Dicroglossidae (F) Hoplobatrachus occipitalis. Calls are shown as representative spectrograms and oscillograms.
Figure 3.
Advertisement calls of Rwandan Arthroleptidae (A) Arthroleptis adolfifriederici, (B) Arthroleptis schubotzi, (C) Leptopelis bocagii, (D) Leptopelis karissimbensis, (E) Leptopelis kivuensis; and Dicroglossidae (F) Hoplobatrachus occipitalis. Calls are shown as representative spectrograms and oscillograms.
3.1.1. Arthroleptis adolfifriederici Nieden, 1911 (Figure 2A and Figure 3A)
Original name and description: Arthroleptis adolfi-friederici—F. Nieden (1911 “1910”). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1910: 436–441 [p. 440].
Type locality: “Bugoiewald” (=“Bugoie Forest”, now Gishwati Forest, Rwanda), “Rugegewald” (=“Rugege Forest”, in the northeastern part of Nyungwe Forest, Rwanda).
External morphology: The Rugege Forest Squeaker is a stout, medium-sized frog with long and slender limbs. Pronounced sexual size dimorphism, SVL of males 27.6–32.0 mm, SVL of females 37.5–46.0 mm [
4,
6,
11,
102]. In reproductive males, spines on the inner side of the second and third fingers. Finger and toe tips with small discs. Coloration in life: Dorsum variable from brown to reddish, mimicking local ground cover, sometimes with a vertebral stripe and dark spots, the venter whitish to marbled. Dorsal skin shagreen with scattered tubercles, ventral skin areolate. Tympanum distinct, half the eye size.
Advertisement call: Males produce series of loud, short whistles. The tonal calls within a series are more or less regularly spaced in time (intercall interval 0.9–1.3 s depending on temperature or social interactions with conspecifics). A single call has a duration of 52–65 ms (average: 58 ms) and a dominant frequency of about 2900 Hz (
Figure 3A).
Distribution: Isolated populations of the Rugege Forest Squeaker inhabit the pristine montane forests (altitudinal range: 1780–2815 m) of extreme eastern DR Congo, Rwanda, Burundi, and southwestern Uganda [
103]. In Rwanda known to occur in Nyungwe (including Cyamudongo Forest), Gishwati-Mukura, and Volcanoes National Parks. Altitudinal range and province records in Rwanda: 1780–2700 m; Southern, Western, and Northern Provinces.
Habitat preference and ecology: Terrestrial. Dwells in the leaf litter of moist montane forest and pristine bamboo forest. The breeding is terrestrial, eggs develop directly within moist leaf litter.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Fragmented range with populations in protected areas. Locally abundant, but population trend unknown. Proposed national conservation status: Least Concern, due to its frequent occurrence in National Parks.
3.1.2. Arthroleptis schubotzi Nieden, 1911 (Figure 2B and Figure 3B)
Original name and description: Arthroleptis schubotzi—F. Nieden (1911 “1910”). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1910: 436–441 [p. 440].
Type locality: “Usumbura” (=Bujumbura, Burundi).
External morphology: Schubotz’s Squeaker is a very small frog. Sexual size dimorphism absent; SVL of males 16.0–23.1 mm, SVL of females up to 23 mm [
6,
32]. In reproductive males, throat is black, third finger elongated with spines on the inner edge. Finger and toe tips with small discs. First finger shorter than the second. Coloration in life: dorsum brown or grey, often with dark spot on head, venter white, mottled or marbled with grey, thighs frequently orange. Dorsal skin granular. Tympanum distinct, half the eye size.
Advertisement call: The calls consist of 5–6 notes that are each composed of three pulses. Call duration is 246–429 ms. A single note has a duration of 23–29 ms (average: 27 ms). Internote interval 34–40 ms. Dominant frequency 4331–4450 Hz (average: 4372 Hz) (
Figure 3B).
Distribution: Schubotz’s Squeaker inhabits the Rift Valley from the northern shores of Lake Tanganyika in western Tanzania to northern Kivu (DR Congo), Rwanda, Burundi, and the western part of Uganda at elevation of 1460–2800 m [
11]. Altitudinal range and province records in Rwanda: 1583–3000 m, Northern and Western Provinces.
Habitat preference and ecology: Terrestrial. Inhabits closed forest, forest edges, and agricultural areas outside forest. The species is found on the ground. Males hide in leaf litter and under fallen tree branches. The breeding is terrestrial, eggs develop directly within moist leaf litter.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Locally abundant, but population trend unknown. Proposed national conservation status: Least Concern, due to its occurrence in national parks (Volcanoes, Nyungwe, and Gishwati-Mukura) and in anthropogenically altered habitats.
3.1.3. Arthroleptis sp.
Taxonomic remark: A small species of Arthroleptis. It differs in morphological characters, advertisement call, and 16S rRNA sequence from the other Arthroleptis species from Rwanda. The taxonomic status of the species will be addressed elsewhere (Dehling et al., in preparation).
Distribution in Rwanda: So far, only known from Western Province.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Population trend and local abundance unknown. Proposed national conservation status: Data Deficient.
3.1.4. Cardioglossa cyaneospila Laurent, 1950 (Figure 2C)
Original name and description: Cardioglossa nigromaculata cyaneospila—R. Laurent (1950). Revue de Zoologie et de Botanique Africaines, 44: 1–18 [p. 4].
Type locality: “rivière Mukozira (Alt. 1800 m.), à 5 km. de Bururi, Urundi” (=Mukazira, south of Bururi, Burundi [
104]).
External morphology: The Blue-spotted Long-fingered Frog is named for its unusual bluish gray color [
28]. The medium-sized species exhibits a pronounced sexual size dimorphism: SVL of males up to 31 mm, SVL of females up to 39 mm [
104]. Males have elongated third fingers. Coloration in life: Dorsum pale purple with three to four large and several smaller dark spots, venter similar with irregular blotches, dark mask from snout tip along the side of the head to tympanum.
Advertisement call: Unknown.
Distribution: Endemic to the Albertine Rift, occurring in the mountains (1467–2248 m) of eastern DR Congo, southwestern Uganda, Rwanda, and southwestern Burundi [
104]. In Rwanda, it inhabits the Gishwati-Mukura and Nyungwe National Parks. Altitudinal range and province records in Rwanda: 1950–2248 m; Southern and Western Provinces.
Habitat preference and ecology: Terrestrial. In the vicinity of streams within montane forests. Breeding takes place in slow-flowing streams.
Conservation status and population trend: IUCN Red List: Near Threatened [
7]. Population trend and local abundance unknown. Proposed national conservation status: Vulnerable, because, although the species occurs in two national parks, it has lost a large part of its former geographic range at medium altitudes due to habiat conversion.
3.1.5. Leptopelis bocagii (Günther, 1865) (Figure 2D and Figure 3C)
Original name and description: Cystignathus bocagii—A. Günther (1865 “1864”). Proceedings of Scientific Meetings of the Zoological Society of London, 1864: 479–482 [p. 481].
Type locality: “Province of Duque de Bragance” [nowadays Calandula], Angola.
Taxonomic remark: The nominal taxon
L. bocagii is probably a species complex [
2,
105].
External morphology: Bocage’s Burrowing Frog is a moderate-sized, robust tree frog [
2,
4,
6,
105,
106,
107]. Moderate sexual size dimorphism: SVL of males 43–52 mm, SVL of females up to 58 mm. Pectoral glands in males well developed. Coloration in life: Dorsum green to brown with an inverted Y-shape dark pattern, venter white without any spotting. Discs on toes and fingers weakly developed, unlike the arboreal
Leptopelis spp.
Advertisement call: The call is a slow, low-pitched croak, often produced in pairs. Males emit pulsed calls lasting 54–107 ms (average: 77 ms) and consisting of 25–38 pulses (average: 33) (
Figure 3C). Pulse rate ranges from 182 to 198 pulses/s. Pulse duration 1–3 ms (average: 2 ms). Interpulse interval 1–4 ms (average: 3 ms). Dominant frequency 709–929 Hz (average: 781 Hz).
Distribution: A widespread species, ranging from Namibia and Angola to Malawi and northwards to Ethiopia at elevations from sea level to about 1900 m [
108,
109]. Altitudinal range and province records in Rwanda: 962–1673 m; Southern, Western, and Eastern Provinces.
Habitat preference and ecology: Fossorial, climbs into vegetation when above the ground. The species inhabits dry and humid savanna and grassland that enters occasionally marginally into clearings in tropical deciduous forest. It lives in anthropogenically disturbed habitats like the surrounding of Ruzizi and Huye (Rwanda) and Nairobi (Kenya) [
7]. The species breeds in temporary pools.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. The species is likely to be overlooked because of its burrowing habits but may be quite common. During the breeding season frogs can occur in great numbers. Proposed national conservation status: Least Concern, because of its frequent occurence in protected sites (Akagera National Park) and in anthropogenically altered environments.
3.1.6. Leptopelis cf. cynnamomeus (Bocage, 1893) (Figure 2E)
Original name and description: Hylambates cynnamomeus—J. V. B. du Bocage (1893). Jornal de sciencias mathematicas, physicas e naturaes, Ser. 2, 3: 115–121 [p. 120].
Type locality: “Quillengues” [Angola] (=Quilengues, Angola; 14.07° S, 14.07° E).
Taxonomic remark: The nominal taxon is probably a complex of cryptic species [
2] and the Rwandan population might not be identical with the topotypic population from Angola.
External morphology: The Cinnamon Tree Frog is a medium-sized
Leptopelis [
2,
6]. SVL of males up to 40 mm, sexual dimorphism unknown. Extralimital populations with brown dorsum with indistinct dark pattern consisting of a dark bar between the eyes that may extend as a triangle backwards to join a pair of diverging dorsolateral bands and a dark line along the side of the head from the tip of the snout to the level of tympanum [
6]. Dorsum of Rwandan specimens greyish green. Males with weakly developed pectoral glands.
Advertisement call: The call has been described as “a clack followed by a whine”, as “a second-long whine followed immediately by a chirp”, and as “a scream followed by a clack” [
2,
6]. Call descriptions may refer to distinct taxa within the species complex. The call of the Rwandan population is unknown.
Distribution: This species (complex) occurs from southern DR Congo and northwestern Zambia to central and eastern Angola. Further records from eastern DR Congo, Rwanda, and Burundi appear to refer to another taxon than the nominal one. Altitudinal range and province records in Rwanda: The only locality currently known is a wetland near Cyamudongo Forest at 1839 m; Western Province.
Habitat preference and ecology: Arboreal. Occurs in savanna, near trees. Breeds in burrows in the ground [
2,
6]. The Rwandan specimens were found in farmland close to forest edge.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed national conservation status: Data Deficient.
3.1.7. Leptopelis karissimbensis Ahl, 1929 (Figure 2F and Figure 3D)
Original name and description: Leptopelis karissimbensis—E. Ahl (1929). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1929: 185–222 [p. 195].
Type locality: “Dorf des Mtulia Gahama am Karissimbi [Deutsch-Ost-Afrika]” (=“Village of Mtulia Gahama on Karissimbi [German East Africa]”, nowadays Mount Karisimbi, Rwanda).
External morphology: The Karisimbi Tree Frog is a moderate-sized, stout species [
4,
6,
55]. Pronounced sexual size dimorphism, SVL of males 27.2–47.1 mm, SVL of females 42.7–47.1 mm. Reproductive males have pectoral glands, a blue vocal sac and often blue pigmentation inside the mouth and on the ventral side of fore and hind limbs. Coloration in life: Dorsum uniformly brownish gray, venter white to cream with brown spots. Juveniles and subadults are bright green with yellow spots at elbow and heel.
Advertisement call: Males emit pulsed calls, often a long one followed by a shorter. A single call has a duration of 534–1305 ms (average: 1105 ms) and consists of 56–132 pulses (average: 88) (
Figure 3D). Pulse duration is 1–3 ms (average: 2 ms), interpulse interval 6–8 ms (average: 7 ms). Dominant frequency at 1100–1395 Hz (average: 1312 Hz). The call has been described elsewhere, but most numerical data have been confounded with those of
L. kivuensis [
55].
Distribution: The species occurs on both sides of the Albertine Rift in Burundi, DR Congo, Rwanda, and Uganda at elevations of 1800–3200 m [
55,
56]. Altitudinal range and province records in Rwanda: 2006–3200 m; Western, Southern, and Northern Provinces.
Habitat preference and ecology: Arboreal. In montane forests including the bamboo zone, near standing water bodies or slow-flowing streams. Abundant in seasonally flooded marshes within forest [
56]. Clutches of eggs are deposited into troughs of wet soil near small temporary pools, semipermanent ponds, and slow-moving streams, tadpoles develop within the water bodies.
Conservation status and population trend: IUCN Red List: Vulnerable [
7]. The species is locally abundant. Proposed national conservation status: Near Threatened, because of the limited geographic distribution in the country, restricted to few localities in Volcanoes and Nyungwe National Parks.
3.1.8. Leptopelis kivuensis Ahl, 1929 (Figure 2G and Figure 3E)
Original name and description: Leptopelis kivuensis—E. Ahl (1929). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1929: 185–222 [p. 206].
Type locality: “Kissenji, südlich des Kivu-See’s” (=“Gisenyi, south of Lake Kivu” [nowadays Rubavu, Western Province, Rwanda]; in error, as Rubavu is situated at the northeastern shore of Lake Kivu).
Taxonomic remark: The nominal taxon is a complex of cryptic species and up to three different species might be present in Rwanda [
72].
External morphology: The Kivu Tree Frog is a moderate-sized, stout species [
2,
4,
6,
55]. Pronounced sexual size dimorphism, males 27.7–37.0 mm, females 45.1–46.5 mm [
55]. Reproductive males are similar to those of
L. karissimbensis but differ by the white coloration of the gular sac. Coloration in life: Dorsum pale brown with a darker brown irregular pattern. Juveniles and subadults brightly green.
Advertisement call: Males emit pulsed calls in series of 2–6 single calls (intercall interval 0.14–0.18 s depending on temperature or social interactions with conspecifics). A single call has a duration of 55–88 ms (average: 69 ms) and includes 5–12 pulses (average: 8) (
Figure 3E). Pulse duration is 2–3 ms (average: 3 ms), interpulse interval 2–4 ms (average: 3 ms). Dominant frequency at 1880–1975 Hz (average: 1922 Hz). The call has been described elsewhere, but most numerical data have been confounded with those of
L. karissimbensis [
55].
Distribution: The species occurs on both sides of the Albertine Rift, in the highlands of western Rwanda and Burundi, North and South Kivu Provinces, DR Congo, and the Ruwenzori Mountains in western Uganda at elevations of 1481–3400 m [
54,
55,
56]. Altitudinal range and province records in Rwanda: 1533–3400 m; Southern, Western, and Northern Provinces.
Habitat preference and ecology: Arboreal. In montane wetlands and forest ecotones, also in farmland and tea plantations near forest. Seasonally flooded forest swamps and shallow pools are used for reproduction.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. This species can be abundant where it occurs. Proposed national conservation status: Least Concern, because of its occurrence within protected areas (Volcanoes, Gishwati-Mukura, and Nyungwe National Parks including Cyamudongo Forest) as well as in farmland.
Four bufonid species have been recorded in Rwanda. While Schismaderma carens is readily distinguished from the three Sclerophrys species by the absence of paratoid glands, the latter species are morphologically similar, but differ in size and color pattern and are well differentiated with respect to partial 16S rRNA sequences. The advertisement call allows for a reliable distinction of reproductive males in the field.
Key to the species of Sclerophys of Rwanda based on morphological characters:
- 1
Large, SVL of adult males >50 mm, adult females 53–120 mm, paratoid gland about 3 times as long as wide, its surface rather smooth without keratinous spines ........................................................................................................... 2
- 1′
Small, SVL of adult males <49 mm, adult females < 55 mm, paratoid gland four times as long as wide, its surface rough with keratinous spines ...... S. berghei
- 2
Light band from middle of paratoid gland to arm insertion, red markings on thigh, breeding males with warty back and normal dorsal colouration ............... S. gutturalis
- 2′
Light band from paratoid gland to arm insertion and red thigh markings absent, breeding males with smooth back and yellow dorsal colouration ............ S. kisoloensis
Key to the bufonid species of Rwanda based on advertisement call features:
- 1
Call consisting of 2–3 pulse groups ............................ Sclerophrys berghei
- 1′
Call consisting of a single pulse group ................................................. 2
- 2
Number of pulses per call < 20 ................................ Sclerophrys gutturalis
- 2′
Number of pulses per call greater......................................................... 3
- 3
Pulse rate > 73 pulses/s ........................................ Sclerophrys kisoloensis
- 3′
Pulse rate lower .................................................... Schismaderma carens
Figure 4.
Species of Bufonidae in Rwanda (A) Schismaderma carens, Eastern Province (photo: Eberhard Fischer), (B) Sclerophrys berghei, Nyungwe NP (photo: J.M.D.), (C) Sclerophrys gutturalis, Akagera NP (photo: J.M.D.), (D) Sclerophrys kisoloensis, Cyamudongo Forest (photo: J.M.D.).
Figure 4.
Species of Bufonidae in Rwanda (A) Schismaderma carens, Eastern Province (photo: Eberhard Fischer), (B) Sclerophrys berghei, Nyungwe NP (photo: J.M.D.), (C) Sclerophrys gutturalis, Akagera NP (photo: J.M.D.), (D) Sclerophrys kisoloensis, Cyamudongo Forest (photo: J.M.D.).
Figure 5.
Advertisement calls of Bufonidae in Rwanda. (A) Schismaderma carens, (B) Sclerophrys berghei, (C) Sclerophrys gutturalis, (D) Sclerophrys kisoloensis. Calls are shown as representative spectrograms and oscillograms.
Figure 5.
Advertisement calls of Bufonidae in Rwanda. (A) Schismaderma carens, (B) Sclerophrys berghei, (C) Sclerophrys gutturalis, (D) Sclerophrys kisoloensis. Calls are shown as representative spectrograms and oscillograms.
3.1.9. Schismaderma carens (Smith, 1848) (Figure 4A and Figure 5A)
Original name and description: Bufo carens—A. Smith (1848). Illustrations of the Zoology of South Africa; consisting chiefly of figures and descriptions of the objects of natural history collected during an expedition into the interior of South Africa, in the years 1834, 1835, and 1836; fitted out by “The Cape of Good Hope Association for exploring Central Africa”. Reptilia. Part 27: Plate 68 (1).
Type locality: “The interior of Southern Africa”.
External morphology: The Red Toad is a large species [
4,
107,
110,
111]. Moderate sexual size dimorphism, SVL of males 54–88 mm, SVL of females 55–92 mm. Nuptial pads on the first three fingers and a subgular vocal sac present in males. Coloration in life: Brick-red to light brown dorsum with a pair of dark spots in the posterior part, the venter whitishly speckled with gray spots. Parotid glands are absent, distinguishing the species from members of
Sclerophrys. Broad glandular dorsolateral ridges running from above the tympanum to the hind limb. Dorsal skin relatively smooth for a toad and less less warty than in
Sclerophrys. Tympanum conspicuously large and round, its diameter equaling eye diameter. Tarsal fold present.
Advertisement call: Call description is based on calls recorded in South Africa [
107]. The call sounds like a loud, long whoop [
6,
107]. Males emit pulsed calls in series (intercall interval 1.256–3.416 ms depending on temperature or social interactions with conspecifics) and often form choruses. A single call has a duration of 690–1702 ms (average: 1118 ms) and consists of 47–74 pulses (average: 58) (
Figure 5A). The pulse rate varies between 44 and 71 pulses/s (average: 59 pulses/s). Pulse duration is 6–10 ms (average: 8 ms), interpulse interval 6–9 ms (average: 7 ms). Dominant frequency at 209–344 Hz (average: 267 Hz).
Distribution: Widely distributed throughout southeastern and eastern Africa from South Africa and Botswana to Kenya, Rwanda, and Uganda at elevations from sea level to 1600 m. Altitudinal range and province records in Rwanda: So far known from a single locality south of Akagera National Park at 1232 m; Eastern Province.
Habitat preference and ecology: Terrestrial. Natural habitats include grassland and wooded savanna. Tolerant to human impact with occurrences on livestock ranches, around human settlements, and on agricultural land. Deep ponds are used for breeeding [
107].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Proposed national conservation status: Vulnerable, because its occurrence is limited to a restricted, non-protected area in the extreme East of the country that is used for livestock breeding and agriculture.
3.1.10. Sclerophrys berghei (Laurent, 1950) comb. nov. (Figure 4B and Figure 5B)
Original name and description: Bufo berghei—Laurent, R. F. (1950). Revue de Zoologie et de Botanique Africaines, 44: 1–18 [p. 1].
Type locality: “Makamba, Terr. de Bururi, Urundi [=Burundi]”.
Taxonomic remark: The species was hitherto considered a synonym of
Sclerophrys funerea (Bocage, 1866) [
112]. Examination of the type series of
Bufo berghei and other specimens in the RMCA revealed significant morphological differences to
S. funerea. Furthermore, the two species differ markedly in the sequences of molecular mitochondrial and nuclear markers [
63]. Therefore, we resurrect the name (as
Sclerophrys berghei comb. nov.) for the populations from Rwanda and Burundi. The phylogenetic relationship of the population from Nyungwe to the topotypic population from Burundi is currently under investigation.
External morphology: A small species of
Sclerophrys, van den Berghe’s Toad exhibits a pronounced sexual size dimorphism, SVL of males 38.3–48.5 mm, SVL of females 47.6–54.9 mm. Nuptial pads (c1usters of black spinules) on the first two fingers of males. Parotid glands about four times as long as wide. Dorsal skin warty with spines. Coloration in life: Dorsum brown with pairs of dark spots [
28].
Advertisement call: Males emit calls consisting of 2–3 pulsed notes in series of up to 9 calls (intercall interval 901–2913 ms). A single call has a duration of 344–702 ms (average: 503 ms) and consists of 15–21 pulses (average: 19) (
Figure 5B). The duration of a single note is 40–51 ms (average: 48 ms) and the number of pulses per note is 5–7. The pulse rate varies between 30 and 44 pulses/s (average: 38 pulses/s). Pulse duration is 4–6 ms (average: 5 ms), interpulse interval 0–1 ms. Dominant frequency at 1033–1119 Hz (average: 1067 Hz). The call has not been described before.
Distribution: As the species was considered a synonym of S. funerea, the distributional range is currently unclear. So far known with certainty from several localities in Burundi and Rwanda. Populations currently assigned to S. funerea from eastern DR Congo and in Uganda possibly also belong to S. berghei. Altitudinal range and province records in Rwanda: 1750–1900 m; Southern and Western Provinces.
Habitat preference and ecology: Terrestrial. On the ground in leaf litter in the interior of rainforests. Breeding probably takes place in streams.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Proposed national conservation status: Data Deficient, because of the lack of information on the distributional range and population size. The species occurs in Nyungwe National Park.
3.1.11. Sclerophrys gutturalis (Power, 1927) (Figure 4C and Figure 5C)
Original name and description: Bufo regularis gutturalis—Power, J. H. 1927. Transactions of the Royal Society of South Africa, 14: 405–422 [p. 416].
Type locality: “Lobatsi” and “Kuruman”, South Africa.
Taxonomic note: Until recently, the population from Rwanda has been referred to as
Sclerophrys (
Bufo)
regularis [
11,
20,
23,
48,
76,
77,
78].
External morphology: A large species of
Sclerophrys, the Guttural Toad exhibits a pronounced sexual size dimorphism, SVL of males 62–91 mm, SVL of females 70–130 mm [
4,
6,
113]. Reproductive males have a yellow, dark-speckled throat, a single subgular vocal sac, enlarged thenar tubercles and black nuptial pads on the outer faces of the first, second and third fingers; the latter is often less obvious. Large, prominent, straight-lined or slightly kidney-shaped parotid glands, with a relatively smooth appearance. The tympanum shows a shallow circular depression. Its diameter ranges between 50% and 70% of that of the eye. Coloration in life: Dorsum dark-olive brown with dark brown patches, the venter cream to whitish. A characteristic light cross between four conspicuous dark brown blotches on the head. Lateral faces of thigh often with red markings, more pronounced in juveniles.
Advertisement call: The call is a lengthy rattling sound. Males emit pulsed calls in series (intercall interval 631–1281 ms) and often form choruses. A single call has a duration of 525–1011 ms (average: 719 ms) and consists of 8–17 pulses (average: 12) (
Figure 5C). The pulse rate varies between 13 and 20 pulses/s (average: 17 pulses/s). Pulse duration is 13–37 ms (average: 24 ms), interpulse interval 24–58 ms (average: 40 ms). Dominant frequency at 301–510 Hz (average: 388 Hz) [
4,
76].
Distribution: A wide geographical range from Uganda and Kenya, through much of eastern and southern Africa except the southwestern parts at elevations from sea level to 2348 m [
6,
79,
113]. Altitudinal range and province records in Rwanda: 960–2348 m; all 5 provinces.
Habitat preference and ecology: Terrestrial. In savanna, grassland, shrubland, dry forests, and the edges of rainforests. Tolerant to human disturbance, present in cultivated wetlands and human settlements. Breeds in shallow permanent or temporary ponds and puddles, vegetated lake shores, slow-moving streams and ditches, also in rice fields and artifical fishponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Proposed national conservation status: Least Concern, because of its occurrence in high numbers in both a protected area (Akagera National Park) and in agricultural wetlands (marais) throughout the country.
3.1.12. Sclerophrys kisoloensis (Loveridge, 1932) (Figure 4D and Figure 5D)
Original name and description: Bufo regularis kisoloensis—A. Loveridge (1932). Occasional Papers of the Boston Society of Natural History, 8: 43–54 [p. 52].
Type locality: “Kisolo, Kigezi district, southwestern Uganda”.
External morphology: The Kisolo Toad is a medium-sized bufonid [
4,
6,
31,
110,
114,
115]. Pronounced sexual size dimorphism, SVL of males 50–71 mm, SVL of females 53–87 mm. Reproductive males are bright yellow during and a short period after amplexus. Unlike
S. gutturalis, breeding males have a shagreen back with flattened warts lacking spines, and the gular pigmentation is never black. Coloration in life: Dorsum olive-brown with dark brown markings, often with a light thin mid-dorsal stripe, the venter lighter. Parotid glands are two to three times longer than wide, separated from the eyelid. Tympanum diameter is smaller than eye diameter.
Advertisement call: Males emit pulsed calls in series (intercall interval 825–1713 ms) and often form choruses. A single call has a duration of 965–1251 ms (average: 1091 ms) and consists of 72–95 pulses (average: 78) (
Figure 5D). The pulse rate varies between 73 and 87 pulses/s (average: 76 pulses/s). Pulse duration is 3–4 ms (average: 3 ms), interpulse interval 9–10 ms (average: 9 ms). Dominant frequency at 411–496 Hz (average: 452 Hz).
Distribution: The species has a fragmented range from central Malawi to Uganda and western and central Kenya in altitudes from 1500 m to 3031 m [
6,
79,
113]. Altitudinal range and province records in Rwanda: 1467–3031 m; Western, Northern, and Southern Provinces.
Habitat preference and ecology: Terrestrial. In wetlands within montane forests. Occurs also in agricultural wetlands near forested areas and near Lake Kivu. Breeds in natural ponds and puddles, slow-moving streams, and ditches, and also in rice fields and artificial ponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Proposed national conservation status: Least Concern, because of its occurrence in protected areas (Volcanoes, Gishwati-Mukura, and Nyungwe National Parks including Cyamudongo Forest) as well as anthropogenically disturbed sites.
This family is represented in Rwanda by a single species. The large, water-associated African Groove-crowned Frog cannot be confused with any other species. Eye with an oval vertical pupil.
3.1.13. Hoplobatrachus occipitalis (Günther, 1858) (Figure 2H and Figure 3F)
Original name and description: Rana occipitalis—A. Günther (1858). Archiv für Naturgeschichte, 24: 319–328 [p. 320].
Type locality: “West-Afrika”.
External morphology: The African Groove-crowned Frog is a robust, very large anuran species. Pronounced sexual size dimorphism: SVL of males 52–110 mm, body mass 24–84 g, SVL of females 65–160 mm, body mass 20–235 g [
4,
6,
116,
117,
118]. Males have paired lateral vocal sacs, that are dark grey, light blue, or cream, and considerably enlarged thenar tubercles. Transverse pale groove behind the eyes. The skin is tuberculate with many enlarged tubercles and short dermal ridges, of which some are arranged to form interrupted dorsolateral glandular folds; slippery due to numerous glands. Feet fully webbed. Large tympanum (70–100% of eye diameter) bordered by a conspicuous, strongly curved supratympanic fold. Coloration in life: Dorsum olive, drab brown or gray with darker markings and a pale occipital band behind the eyes, venter white with large irregular dark spots.
Advertisement call: Call description is based on calls recorded in Mauritania [
119]. The pulsed advertisement call is a low pitched, deep roar. Males usually emit call series (intercall interval 1438–1630 ms) and often form choruses. A single call has a duration of 223–238 ms (average: 232 ms) and consists of 19–25 pulses (average: 22) (
Figure 3F). The pulse rate varies between 83 and 109 pulses/s (average: 97 pulses/s). Pulse duration is 13–37 ms (average: 24 ms), interpulse interval less than 1 ms. Dominant frequency at 316–532 Hz (average: 481 Hz). For additional call descriptions see [
6,
120,
121,
122].
Distribution: Most parts of sub-Saharan Africa from Mauretania and southern Mali through West Africa eastwards to Ethiopia, southwards to Zambia and Angola and Gabon, except for the central Congo Basin. Altitudinal range and province records in Rwanda: Known only from the Bugarama area at 930–980 m; Western Province.
Habitat preference and ecology: Semi-aquatic, always closely associated with water bodies. In savanna, farmland, and disturbed edges of forests. Breeds on riverbanks, in rock pools, and in savanna ponds. In Rwanda only in agricultural wetlands.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Proposed national conservation status: Least Concern, because it occurs in high numbers in anthropogenically altered sites.
The Hyperoliidae are the most diverse frog family in Rwanda and comprise 17 species in five genera, i.e., Afrixalus (3 species), Callixalus (1 species), Hylambates (1 species), Hyperolius (11 species), and Kassina (1 species). Morphological or bioacoustic distinction of species is difficult in some genera, but usually feasible in combination. All taxa are well differentiated with respect to partial 16S sequences. The advertisement call (unknown for Callixalus pictus) allows for a reliable distinction of reproductive males in the field.
Key to the species of Afrixalus of Rwanda based on morphological characters:
- 1
Dorsal pattern consisting of longitudinal stripes ....................................2
- 1′
Dorsal pattern consisting of irregularly shaped blotches and spots ....................................................................................................... A. phantasma
- 2
Two pairs of sharply delimited longitudinal dark brown dorsal bands, central one extending from level of anterior corners of eyes to end of body, running over bases of upper eyelids, light brown area between the bands without dark spots or speckles ........................................................................... A. quadrivittatus
- 2′
Usually a single pair of narrow longitudinal dark paravertebral stripes (inconspicuous second pair of lateral stripes rarely present), from level of anterior corners of eyes to end of body, not running over bases of upper eyelids, dorsum and limbs densely covered with small dark spots ............................ A. orophilus
Key to the species of Hyperolius of Rwanda based on morphological characters:
- 1
Ventral skin without white pigmentation, translucent with a green or bluish hue, internal organs well visible through skin ............................................... 2
- 1′
Ventral skin or peritoneum with whitish pigmentation, internal organs not or only partly visible ................................................................................... 4
- 2
Large, SVL of males > 27 mm, dorsum coarsely granular, green with heavy brown and black speckling .... 3
- 2′
Smaller, SVL of adult males < 24 mm, dorsum smooth, uniformly bright green, rarely with few brown dots ................... H. jackie
- 3
Area between tip of snout and anterior corners of eyes brighter than the rest of the dorsum ... H. frontalis
- 3′
Area not brighter than the rest of the dorsum ................ H. discodactylus
- 4
Body elongated, head small and narrow, snout long and acuminate ............... H. rwandae
- 4′
Body not markedly elongated, head wide, snout rounded or subacuminate ......................... 5
- 5
Colouration very variable but usually dorsum darker than flanks, separated from each other by narrow line of bright yellow to yellowish green pigmentation from tip of snout along edge of eyelid to groin ................................ H. castaneus
- 5′
Narrow yellow or green line between dorsum and flanks absent ................................ 6
- 6
Light, straight-lined band running from tip of snout on both sides above nostril, along lateral edge of eyelid, above tympanic area along body side the groin, flank region ventral to band without heavy white pigmentation .... H. cinnamomeoventris (male)
- 6′
Light lateral band absent or, if present, not running in straight line, through different body areas or excluding the eyelid, or, if straight-lined, flank area ventral to band with dense white pigmentation ............ 7
- 7
Dorsum uniformly bright to dark green, rarely brownish or greyish; well-defined, although sometimes interrupted black stripe from nostrils to eye, through tympanic area onto flanks (if absent, dorsum bright green), preaxial side of thigh with sharply delimited bright red blotch .... 8
- 7′
Dorsal colouration different, greyish brown if uniform, always without a black strip from nostril to flank, thigh without sharply delimited red blotch but leg can be red overall or thigh with diffuse red pigmentation ................... 9
- 8
Green colouration on dorsum, flanks, and dorsal faces of limbs sharply delimited by a black edge from largely unpigmented ventral colouration that extends around arm insertion, disrupting the colouration of arm and flank, SVL up to 23 mm ................................. H. cinnamomeoventris (female)
- 8′
Dorsum green, brown, or greyish, lower flanks and venter white, no disruption of colouration of arm and flank, SVL of adult males (with gular gland) > 23.5 mm, of adult females (no gular gland) > 25 mm .................................... H. kivuensis
- 9
Webbing reaching to distal subarticular tubercle on third and fifth toes, colouration variable but flanks with conspicuously dense white pigmentation forming well defined stripes or spots or diffusely delimited blotches ............. H. lateralis
- 9′
Webbing reaching to disk on third and fifth toes, colouration highly variable but usually without conspicuous dense white pigmentation on flanks ............. H. glandicolor, H. parallelus and H. viridiflavus (very variable species, closely resembling each other, rendering their identification based on external characters in many cases impossible. They are unequivocally distinguished by genetics.)
Key to the hyperoliid species of Rwanda based on advertisement call features:
- 1
Call consisting of two or more clearly audible elements ......................... 2
- 1′
Call consisting of a single short vocalisation ........................................ 6
- 2
Call consisting of 1–3 very short (<12 ms) notes (clicks)................................................ Hyperolius cinnamomeoventris (part.)
- 2′
Call consisting of a series of usually >3 longer (>30 ms) notes ................. 3
- 3
Call consisting of a series of 2–13 pulsed notes, repeated at 4.8–6.3 notes/s, each consisting of 10–18 pulses and lasting 54–107 ms ............... Hyperolius jackie
- 3′
Note repetition rate < 8/s, repeated notes briefer .................................. 4
- 4
Call consisting of an initiating long note, composed of 6 pulses, immediately followed by a long series of 17–138 briefer 3-pulse notes, repeated at approximately 9 notes/s, each lasting <40 ms ........................ Afrixalus quadrivittatus
- 4′
Call a shorter trill, consisting of <16 notes ........................................... 5
- 5
Trill consisting of 4–6 notes, repeated at 7–12 notes/s .. Afrixalus phantasma
- 5′
Trill consisting of 9–15 notes, repeated at 14–25 notes/s .... Afrixalus orophilus
- 6
Dominant frequency markedly upward-modulated .............................. 7
- 6′
Dominant frequency not modulated ................................................... 8
- 7
Frequency modulation moderate, frequency not rising to 1000 Hz, call duration < 100 ms, note fully tonal ................................................. Hylambates verrucosus
- 7′
Frequency modulation strongly expressed, frequency rising to above 1000 Hz, call duration > 120 ms, note initially distinctly pulsed ...... Kassina senegalensis
- 8
Call consisting of 1–3 very short (<12 ms) notes (clicks) .................................................... Hyperolius cinnamomeoventris (part.)
- 8′
Note duration > 12 ms .................................................................. 9
- 9
Call duration > 150 ms, interpulse interval increasing towards the end of call ..................................................................... Hyperolius discodactylus
- 9′
Call duration briefer .................................................................... 10
- 10
Calls not pulsed or indistinctly pulsed in the first half only ................. 11
- 10′
Calls distinctly pulsed throughout .................................................. 13
- 11
Maximum amplitude at the beginning of the call ................................ 12
- 11′
Maximum amplitude in the middle of the call .......... Hyperolius glandicolor
- 12
Call indistinctly pulsed ........................................ Hyperolius parallelus
- 12′
Call unpulsed ................................................... Hyperolius viridiflavus
- 13
First pulse of call separated by a long interval from the successive pulses, resulting in a cracking sound ...................................... Hyperolius kivuensis
- 13′
Call different ............................................................................... 14
- 14
Buzzing call, consisting of >10 pulses, repeated at >250 pulses/s at the beginning, followed by 1–3 isolated pulses ........................... Hyperolius rwandae
- 14′
Notes consisting of <10 pulses ..........................................................15
- 15
Maximum amplitude at second pulse ................................................. 16
- 15′
Maximum amplitude later ...................................... Hyperolius castaneus
- 16
Pulses separated by distinct interpulse intervals (>2 ms) ........................................................................... Hyperolius frontalis
- 16′
Interpulse intervals indistinct (<1 ms) ..................... Hyperolius lateralis
3.1.14. Afrixalus orophilus (Laurent, 1947) (Figure 6A and Figure 7A)
Original name and description: Megalixalus orophilus—R. Laurent (1947). Bulletin du Musée royal d’Histoire naturelle de Belgique, 23 (7): 1–8 [p. 1].
Figure 6.
Rwandan species of Hyperoliidae (A) Afrixalus orophilus, Rugezi Wetland (photo: J.M.D.), (B) Afrixalus phantasma, Nyungwe NP (photo: J.M.D.), (C) Afrixalus quadrivittatus, Huye (photo: J.M.D.), (D) Callixalus pictus, preserved holotype, RMCA 105145, from Rutsiro, Western Province (photo: J.M.D.), (E) Hylambates verrucosus, Kahuzi-Biega NP, DR Congo (photo: Bonny Dumbo), (F) Hyperolius castaneus, Nyungwe NP (photo: J.M.D.), (G) Hyperolius cinnamomeoventris, Huye (photo: J.M.D.), (H) Hyperolius discodactylus, Nyungwe NP (photo: J.M.D.).
Figure 6.
Rwandan species of Hyperoliidae (A) Afrixalus orophilus, Rugezi Wetland (photo: J.M.D.), (B) Afrixalus phantasma, Nyungwe NP (photo: J.M.D.), (C) Afrixalus quadrivittatus, Huye (photo: J.M.D.), (D) Callixalus pictus, preserved holotype, RMCA 105145, from Rutsiro, Western Province (photo: J.M.D.), (E) Hylambates verrucosus, Kahuzi-Biega NP, DR Congo (photo: Bonny Dumbo), (F) Hyperolius castaneus, Nyungwe NP (photo: J.M.D.), (G) Hyperolius cinnamomeoventris, Huye (photo: J.M.D.), (H) Hyperolius discodactylus, Nyungwe NP (photo: J.M.D.).
Figure 7.
Advertisement calls of Hyperoliidae in Rwanda, I. (A) Afrixalus orophilus, (B) Afrixalus phantasma, (C) Afrixalus quadrivittatus, (D) Hylambates verrucosus, (E) Kassina senegalenis, (F) Hyperolius castaneus, (G) Hyperolius cinnamomeoventris, (H) Hyperolius discodactylus. Calls are shown as representative spectrograms and oscillograms.
Figure 7.
Advertisement calls of Hyperoliidae in Rwanda, I. (A) Afrixalus orophilus, (B) Afrixalus phantasma, (C) Afrixalus quadrivittatus, (D) Hylambates verrucosus, (E) Kassina senegalenis, (F) Hyperolius castaneus, (G) Hyperolius cinnamomeoventris, (H) Hyperolius discodactylus. Calls are shown as representative spectrograms and oscillograms.
Type locality: “Lac Magera (2,000 m.) [Parc National Albert]” (=Lake Magera, 1°25′28″ S 29°3′23″ E, North Kivu Province, DR Congo).
External morphology: The Montane Spiny Reed Frog is a small species with hardly expressed sexual size dimorphism: males 18.3–22.6 mm, females 19.3–24.9 mm [
4,
27,
44,
86]. Eyes large and protruding. Two conspicuous narrow dark dorsal bands, with little contrast to ground color, converging on the head.
Advertisement call: A fast trill, typically consisting of 9–15 notes; total call duration 462–841 ms; dominant frequency slightly increasing from 2900–3180 Hz at the beginning of the call to 3090–3280 Hz at the end (
Figure 7A) [
87].
Distribution: The species occurs in an altitudinal range from 950 to 2800 m in the DR Congo, Burundi, Rwanda, and southwestern Uganda [
27,
44]. In Rwanda, it is known from Nyungwe National Park and wetlands in the Northern Province [
44,
87]. Altitudinal range and province records in Rwanda: 1865–2400 m; Western and Northern Provinces.
Habitat preference and ecology: Arboreal. In montane swamps, often found in reeds, papyrus, and bamboo. Breeds in small pools of standing water and in seasonally flooded meadows [
44,
87].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed national conservation status: Endangered, because the species has lost most of its former habitat and its distribution is currently limited to unprotected wetland that is under threat of conversion into farmland.
3.1.15. Afrixalus phantasma Dehling, Greenbaum, Kusamba & Portik in Greenbaum et al., 2022 (Figure 6B and Figure 7B)
Original name and description: Afrixalus phantasma—J. M. Dehling, E. Greenbaum, C. Kusamba & D. M. Portik in E. Greenbaum, D. M. Portik, K. E. Allen, E. R. Vaughan, G. Badjedjea, M. F. Barej, M. Behangana, N. Conkey, B. Dumbo, L. N. Gonwouo, M. Hirschfeld, D. F. Hughes, F. Igunzi, C. Kusamba, W. Lukwago, F. M. Masudi, J. Penner, J. M. Reyes, M.-O. Rödel, C. E. Roelke, S. Romero & J. M. Dehling (2022). Zootaxa, 5174: 201–232 [p. 213].
Type locality: “Gishwati Forest (01.823745° S, 29.360373° E, 2084 m), now part of Gishwati-Mukura National Park, Western Province, Rwanda”.
External morphology: The Ghost Spiny Reed Frog is a small species with a short body and large protruding eyes [
86]. Moderate sexual size dimorphism: SVL of males 20.1–24.6 mm, SVL of females 22.6–26.4 mm. Coloration in life: At night, dorsal side of head, trunk, forelimbs, crus and tarsus light yellowish brown with more or less regular dark brown speckling; indistinct darker brown transverse stripe between upper eyelids; large dark brown, irregularly shaped spot in scapula region, weak brown stripe along canthus, continuing behind eye on anterior half of trunk on both sides, ventral side of head, trunk, and limbs largely unpigmented and translucent. At daytime, basic dorsal coloration brighter, very light brown to bright cream-colored; dark brown dorsal speckles, spots and stripes in more pronounced contrast to basic coloration, more distinctly visible.
Advertisement call: Five to six, rarely four pulse groups (notes), repeated at a rate of 7.1–11.4/s. Each note consisting of 10–11 pulses, that are repeated at a rate of 190–292/s, resulting in a note length of 36–94 ms (
Figure 7B). Amplitude modulation prominent within individual notes. Energy maximum at 3020–3810 Hz without marked frequency modulation. Prominent harmonics at about 6000–7000 Hz and 9000–11,000 Hz [
86].
Distribution: Western Rwanda (Nyungwe and Gishwati Forest) and eastern DR Congo (in and near Kahuzi-Biega National Park, Itombwe Plateau and Kabobo Plateau; very likely also in southwestern Uganda and northwestern Burundi [
86]. Altitudinal range and province records in Rwanda: 1962–2084 m; Western Province.
Habitat preference and ecology: Occurs in swamps in forest openings, or at forest edges. Males call from leaves above standing bodies of water with thick lower vegetation cover.
Conservation status and population trend: IUCN Red List: Not evaluated [
7] but proposed as Vulnerable [
86]. Proposed national conservation status: Near Threatened, because of the limited geographic distribution in the country at few localities in Gishwati-Mukura and Nyungwe National Parks.
3.1.16. Afrixalus quadrivittatus (Werner, 1908) (Figure 6C and Figure 7C)
Original name and description: Megalixalus leptosomus subsp. quadrivittata—F. Werner (1908 “1907”). Sitzungsberichte der mathematisch-naturwissenschaftlichen Klasse der Kaiserlichen Akademie der Wissenschaften, 116: 1823–1926 [p. 1900].
Type locality: “am Nil bei Khor Atta [angloägyptischer Sudan]” (= “by the Nile near Khor Attar [Anglo-Egyptian Sudan]”, 9.33° N, 31.41° E, 400 m, nowadays Jonglei State, Republic of South Sudan [
99].
Taxonomic remark: This taxon is probably a complex of cryptic species [
123,
124].
External morphology: The Four-lined Spiny Reed Frog is a small species. Sexual size dimorphism weakly expressed: SVL of males 23–27 mm, SVL of females 25–28 mm [
2,
6,
38]. Body long, limbs relatively short, eyes large. Reproductive males possess small spines on dorsum, head, and legs, venter smooth without spines. Coloration in life: dorsum brown with three pale longitudinal stripes converging at head, venter creamy white.
Advertisement call: The pulsed advertisement call is composed of a long introductory note (buzz), followed by >20 short notes (clicks comprised of 2–3 pulses) [
2,
6,
76]. Males usually emit single calls. A call has a duration of 859–7598 ms (average: 2855 ms) and consists of 17–138 notes (average: 68) (
Figure 7C). The pulse rate varies between 16 and 30 pulses/s (average: 24 pulses/s). Pulse duration is 3–8 ms (average: 6 ms). Interpulse interval is 4–11 ms (average: 7 ms). Dominant frequency 3078–3511 Hz (average: 3251 Hz). Calls have been described in detail elsewhere [
76,
123].
Distribution: The nominal species to which the Rwandan specimens belong is found in eastern DR Congo, southern Ethiopia, Kenya, South Sudan, Rwanda, Tanzania Uganda, and northern Zambia [
6]. It is unclear whether populations reported from West and Central Africa are conspecific with the nominal taxon [
124,
125]. Altitudinal range and province records in Rwanda: 962–1807 m; in all five provinces.
Habitat preference and ecology: Arboreal. In moist and dry savanna, agricultural areas, and degraded former forest. Breeds in pools of standing water. Eggs are deposited above the water surface on leaves that are subsequently folded. The tadpole has been depicted by Ref. [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. It is generally an abundant and adaptable species that is tolerant to human habitat disturbance. In Rwanda, it is a common and abundant species of the agricultural wetlands (marais) as well as natural savannas. Proposed national conservation status: Least Concern, because of its abundance in protected natural areas (Akagera National Park) as well as anthropogenically disturbed sites.
3.1.17. Callixalus pictus Laurent, 1950 (Figure 6D)
Original name and description: Callixalus pictus—R. Laurent (1950). Revue de Zoologie et de Botanique Africaines, 44: 1–18 [p. 6].
Type locality: “Lutsiro (Alt. 2600 m.), Territoire de Kisenyi” (=Rutsiro, Western Province, Rwanda).
External morphology: The Painted Beauty Frog is large, stout hyperoliid [
2,
6,
28]. Moderate sexual size dimorphism: SVL of males up to 37 mm, SVL of females up to 43 mm. Dorsum finely tuberculate with scattered larger tubercles, digital discs enlarged, webbing reduced. No vocal sac and no vocal sac openings. Dorsum chocolate brown to almost black with numerous small orange or golden spots. Hidden parts of limbs yellow to orange, venter pink, mauve or bluish, whitish in juveniles. Iris brown.
Advertisement call: Unknown. The species lacks a vocal sac and a tympanum and is therefore presumed to be mute.
Distribution: Itombwe and Kabobo highlands of eastern DR Congo, Gishwati-Mukura and Nyunge National Parks of western Rwanda at 2100–2600 m. Historical altitudinal range and province records in Rwanda: 2600 m; Western Province.
Habitat preference and ecology: Montane bamboo forest, in broken bamboo stalks at 2–4 m above the ground, or, where bamboo is scarce, between bark and moss, on tree trunks. Breeding habitat is unknown, but the species is presumed to breed in phytotelmata or other small bodies of standing water [
36].
Conservation status and population trend: IUCN Red List: Vulnerable [
7]. Population trend unknown. Proposed national conservation status: Critically Endangered, due to the loss of almost the complete former habitat in Rwanda during the past 50 years and the lack of recent records from the country. The species might even be extinct.
3.1.18. Hylambates verrucosus Boulenger, 1912 (Figure 6E and Figure 7D)
Original name and description: Hylambates verrucosus—G. A. Boulenger (1912). The Annals and Magazine of Natural History including Zoology, Botany, and Geology, Eighth Series, 10: 140–142 [p. 141].
Type locality: “Mabira Forest, Chagwe, Uganda”.
External morphology: The Warty Stripe-legged Frog is a large tree frog [
2,
4,
6,
48]. Sexual size dimorphism apparently absent, males 46–52 mm, females up to 51 mm [
2]. Reproductive males have numerous white asperities on the dorsal skin. Coloration in life: Dorsum uniformly black or dark grey with darker markings. Femur and corresponding part of flanks, barred with black and yellow or orange. Venter dark brown with few small spots.
Advertisement call: The tonal call is brief rising whistle, lasting 70–150 ms [
2,
4,
6]. Dominant frequency is steeply upward modulated from 400 to 900 Hz, like the call of
Kassina senegalensis (
Figure 7D). A spectrogram of the call has been shown in Ref. [
2].
Distribution: The species occurs in parts of the DR Congo, Rwanda, and Uganda at elevations of 600–2000 m. Altitudinal range and province records in Rwanda: Recorded from only a single locality at 1936 m in Nyungwe National Park; Western Province.
Habitat preference and ecology: Aboreal. Occurs in lowland and montane forest, usually associated with edge habitats to open areas. Tolerant to human disturbance. Breeds in temporary ponds. Eggs are placed singly below water.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed national conservation status: Vulnerable, due to the very limited geographic range in the country.
3.1.19. Hyperolius castaneus Ahl, 1931 (Figure 6F and Figure 7F)
Original name and description: Hyperolius castaneus—E. Ahl (1931). Mitteilungen aus dem Zoologischen Museum in Berlin, 17 (1): 1–132 [p. 31].
Type locality: “Vulkangebiet nord-östlich des Kivu-See’s” (=“volcano area northeast of Lake Kivu”; i.e., the Virunga Mountains, probably in Rwanda).
External morphology: The Chestnut Reed Frog is a medium-sized species of
Hyperolius [
2,
4,
6,
57,
84,
85]. Pronounced sexual size dimorphism: SVL of males 20.3–27.2 mm (average: 23.8 mm), SVL of females 24.1–36 mm (average: 28.9 mm). Reproductive males differ from females by a yellow throat and a vocal sac. Coloration in life: Dorsum brown to green, sometimes with irregular darker spots forming variable patterns, venter and feet yellow to reddish. Metamorphs are uniformly green, sometimes with yellow to white lateral lines and small black spots.
Advertisement call: The pulsed call is brief and high-pitched, emitted singly or in series (intercall interval 726–3894 ms depending on temperature or social interactions with conspecifics) [
2,
57,
58]. The call has a duration of 26–73 ms (average: 45 ms) and consists of 6–11 pulses (average: 8) (
Figure 7F). Pulse rate varies between 125 and 280 pulses/s (average: 193 pulses/s). Pulse duration is 2–4 ms (average: 2 ms). Interpulse interval is 2–7 ms (average: 4 ms). Dominant frequency at 2670–3402 Hz (average: 2991 Hz). Calls have been described in detail elsewhere [
57,
58].
Distribution: An Albertine Rift endemic of montane forests at elevations of 1600–2879 m in eastern DR Congo, western Rwanda, and western Uganda [
2,
6,
57,
58]. In Rwanda, this species occurs in Volcanoes, Gishwati-Mukura, and Nyungwe National Parks and their vicinity with degraded rests of montane forest. Altitudinal range and province records in Rwanda: 1813–3031 m; Northern, Western, and Southern Provinces.
Habitat preference and ecology: Arboreal. In pristine and degraded montane forests. Breeding takes place in small ponds in wetlands within or at the edge of forests. Clutches of 20–57 eggs are deposited on moss pads (
Polytrichum commune, Isotachis aubertii) or grass tussocks (
Andropogon shirensis) 2–5 cm above the water level [
83]. Egg masses are occasionally infested by frogflies. Seven days after oviposition, tadpoles (Gosner stage 24–25) move into the water and begin their free-swimming tadpole stage. The tadpole has been described in detail elsewhere [
83,
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. It is an abundant species that is widespread in montane forests. In Rwanda, it is one of the most common and abundant species of montane wetlands and occurs abundantly in three protected areas. Proposed national conservation status: Least Concern.
3.1.20. Hyperolius cinnamomeoventris Bocage, 1866 (Figure 6G and Figure 7G)
Original name and description: Hyperolius cinnamomeo-ventris—J. V. B. du Bocage (1866). Jornal de sciencias mathematicas, physicas e naturaes, 1: 57–78 [p. 77].
Type locality: “Duque de Bragança” (=Calandula, Malanje Province, Angola).
Taxonomic remark: This taxon is probably a complex of at least three cryptic species [
74,
126,
127].
External morphology: The Cinnamon-bellied Reed Frog is a small species of
Hyperolius [
2,
4,
6,
57,
84,
85]. Moderate sexual size dimorphism: SVL of males 18.6–22.8 mm (average: 21.1 mm), SVL of females 19–23 mm. Reproductive males differ from females by a small gular sac flap and in coloration. Coloration in life: Dorsum of males dark brown to light brown with white to yellow dorsolateral bands, dorsum of females uniformly green, venter whitish to yellow, thighs of breeding individuals reddish.
Advertisement call: The call has a duration of 550–2003 ms (average: 664 ms) and typically consists of 2–3 short notes (clicks), each consisting of 2–4 pulses [
2,
4,
6,
76,
126,
128] (
Figure 7G). Pulse rate varies between 125 and 280 pulses/s (average: 193 pulses/s). Pulse duration is 1–5 ms (average: 3 ms). Interpulse interval is usually 0 ms, interval between notes up to 593 ms. Dominant frequency at 3083–3992 Hz (average: 3245 Hz). Calls of Rwandan specimens have been described in detail elsewhere [
76].
Distribution: The Cinnamon-bellied-Reed-Frog-species complex occurs in Central and eastern Africa, from Cameroon to Kenya and south to Angola, Zambia, and Rwanda [
2,
6,
74,
76]. Altitudinal range and province records in Rwanda: 1287–1785 m; Southern and Eastern Provinces. Erroneously reported from Volcanoes National Park based on misidentified
H. castaneus [
56,
57] and
H. glandicolor [
129,
130].
Habitat preference and ecology: Arboreal. In savanna at larger water bodies, papyrus wetland, and other open habitats. The species breeds in dense vegetation on lake shores, small ponds or ditches in wetlands including farmland. In Rwanda, it often co-occurs with
H. lateralis in natural or recovering reed areas of wetland. The tadpole has been depicted elsewhere [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed national conservation status: Least Concern, because the species is common in a protected area (Akagera National Park) and also occurs scatteredly in farmland.
3.1.21. Hyperolius discodactylus Ahl, 1931 (Figure 6H and Figure 7H)
Original name and description: Hyperolius discodactylus—E. Ahl (1931). Mitteilungen aus dem Zoologischen Museum in Berlin, 17 (1): 1–132 [p. 89].
Type locality: “Rugege-Wald” (=“Rugege Forest”, in the northeastern part of Nyungwe Forest, Rwanda).
External morphology: The Disc-fingered Reed Frog is a medium to large species of Hyperolius [
2,
4,
6,
38,
62,
84,
85]. Moderate sexual size dimorphism: SVL of males 29.0–34.1 mm (average: 31.8 mm), SVL of females up to 40 mm. Reproductive males differ from females by a bright green throat and in coloration. Coloration in life: dorsum green with dark speckles, rarely brownish green, throat and venter yellow-orange, underside of hands and feet pink.
Advertisement call: The call is a brief, low-pitched buzz, sometimes emitted in short series of up to six calls [
2,
4,
61]. The call has a duration of 167–362 ms (average: 254 ms) and includes 19–50 pulses (average: 37) (
Figure 7H). Relative amplitude rises from the beginning to the middle of the call and declines towards the end. Pulse repetition is highest at the beginning (142–250 pulses/s) and declines towards the end (58–125 pulses/s). Pulse duration is 7–16 ms (average: 12 ms), interpulse intervals vary from 0 ms at the beginning of the call to 19 ms at the end. There is a modulation of the dominant frequency from between 1950 and 2350 Hz at the beginning of the call to a maximum of 2350–2750 Hz at about three-fourths the call length (
Figure 7H). The call has been described in detail elsewhere [
62].
Distribution: The geographical range of this species is fragmented because it is restricted to montane forest in eastern DR Congo, Burundi, Rwanda, and Uganda in the altitudinal range of 1600–2700 m [
2,
62]. The range includes Volcanoes, Gishwati-Mukura, and Nyungwe National Parks in Rwanda. Altitudinal range and province records in Rwanda: 1813–2379 m; Southern, Western, and Northern provinces.
Habitat preference and ecology: Arboreal. In montane forests associated with rivers, streams, and swamps. Breeds in streams, in sections with slow current. The tadpole has been described elsewhere [
85]. Males call from trees near streams, 1–5 m above ground [
62]. The species is capable of parachuting [
81]. Sexual maturity is attained within the first year following metamorphosis, longevity is two years [
84].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Locally abundant, but population trend unknown. Proposed category for the National Red List: Least Concern, due to its frequent occurrence in protected areas.
Figure 8.
Rwandan species of Hyperoliidae (A) Hyperolius frontalis, Nyungwe NP (photo: J.M.D.), (B) Hyperolius glandicolor, Volcanoes NP (photo: J.M.D.), (C) Hyperolius jackie, Nyungwe NP (photo: JMD), (D) Hyperolius kivuensis, Huye (photo: J.M.D.), (E) Hyperolius lateralis, Huye (photo: J.M.D.), (F) Hyperolius parallelus, Ruzizi (photo: J.M.D.), (G) Hyperolius rwandae, Huye (photo: J.M.D.), (H) Hyperolius viridiflavus, Kigali (photo: J.M.D.).
Figure 8.
Rwandan species of Hyperoliidae (A) Hyperolius frontalis, Nyungwe NP (photo: J.M.D.), (B) Hyperolius glandicolor, Volcanoes NP (photo: J.M.D.), (C) Hyperolius jackie, Nyungwe NP (photo: JMD), (D) Hyperolius kivuensis, Huye (photo: J.M.D.), (E) Hyperolius lateralis, Huye (photo: J.M.D.), (F) Hyperolius parallelus, Ruzizi (photo: J.M.D.), (G) Hyperolius rwandae, Huye (photo: J.M.D.), (H) Hyperolius viridiflavus, Kigali (photo: J.M.D.).
Figure 9.
Advertisement calls of Hyperoliidae in Rwanda, II. (A) Hyperolius frontalis, (B) Hyperolius glandicolor, (C) Hyperolius jackie, (D) Hyperolius kivuensis, (E) Hyperolius lateralis, (F) Hyperolius parallelus, (G) Hyperolius rwandae, (H) Hyperolius viridiflavus. Calls are shown as representative spectrograms and oscillograms.
Figure 9.
Advertisement calls of Hyperoliidae in Rwanda, II. (A) Hyperolius frontalis, (B) Hyperolius glandicolor, (C) Hyperolius jackie, (D) Hyperolius kivuensis, (E) Hyperolius lateralis, (F) Hyperolius parallelus, (G) Hyperolius rwandae, (H) Hyperolius viridiflavus. Calls are shown as representative spectrograms and oscillograms.
3.1.22. Hyperolius frontalis Laurent, 1950 (Figure 8A and Figure 9A)
Original name and description: Hyperolius frontalis—R. Laurent (1950). Revue de Zoologie et de Botanique Africaines, 44: 1–18 [p. 11].
Type locality: “Bushoho (Alt. 1100 m.) à 10 km. au nord-ouest de Bunyakiri, Territoire de Kalehe, Kivu” (=“Bushoho, 1100 m, 10 km northwest of Bunyakiri, Kalehe Territory, Kivu [nowadays South Kivu Province]”, DR Congo).
External morphology: The White-Snouted Reed Frog is a medium-sized species of
Hyperolius [
2,
4,
28,
85]. Moderate sexual size dimorphism: SVL of males 27.6–31.0 mm, SVL of females up to 35 mm. Reproductive males differ from females by a green throat (as in
H. discodactylus). Coloration in life: dorsum green (speckled with darker markings) with an irregular, yellow or golden triangle on the snout, the pale marking often continues posteriorly as short line or a series of small dots.
Advertisement call: The call is a brief buzz, sometimes in short series of 2–3 calls (intercall interval about 10 s depending on temperature or social interactions with conspecifics) [
2,
4,
6]. A call has a duration of 64–69 ms (average: 65 ms) and consists of 8 pulses (
Figure 9A). The pulse rate varies between 116 and 125 pulses/s. Pulse duration is 4–7 ms (average: 6 ms). Interpulse interval is 3–4 ms (average: 3 ms). Dominant frequency 2996–3080 Hz (average: 3030 Hz).
Distribution: The species occurs from northwest of Mount Rwenzori in the eastern DR Congo, south to west of Lake Kivu, in Bwindi Impenetrable Forest in south-western Uganda, and in Nyungwe National Park in Rwanda, in an altitudinal range of 700–2000 m [
2,
4,
6,
28]. Recorded only from the southern part of Nyungwe National Park in Rwanda. Altitudinal range and province records in Rwanda: 2283 m; Western Province.
Habitat preference and ecology: Arboreal. In upper lowland rainforest and montane forest associated with streams and swamps. Breeds in small ponds and stream sections with slow current. Males call from dense vegetation overhanging ponds. The opaque white eggs (diameter: 5 mm) are attached to vegetation up to 3 m above water [
2]. Clutch size is ca. 24. The tadpole has been described elsewhere [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Locally abundant, but population trend unknown. Proposed category for the National Red List: Vulnerable, because of the very restricted range in the country, at a single known locality in a protected area.
3.1.23. Hyperolius glandicolor Peters, 1878 (Figure 8B and Figure 9B)
Original name and description: Hyperolius glandicolor—W. Peters (1878). Monatsberichte der Königlich Preussischen Akademie der Wissenschaften zu Berlin. Aus dem Jahre 1878: 194–209 [p. 208].
Type locality: “Taita”, Kenya.
External morphology: The Montane Common Reed Frog is a medium-to-large species of
Hyperolius [
2,
4,
85]. Pronounced sexual size dimorphism: SVL of males 20.5–33.2 mm (average: 25.2 mm), SVL of females 26–35 mm (average: 31.2 mm). Coloration in life: not as variable as in the other species of the viridiflavus group in Rwanda; dorsum of males yellow and dark brown, dorsum of females dark grey, venter and limbs red.
Advertisement call: The call is a brief click, often emitted in series (intercall interval between 1 and 4 s depending on temperature or social interactions with conspecifics) [
4]. The tonal call structure is common in advertisement calls of species in the
H. viridiflavus group. The call has a duration of 39–41 ms (average: 39 ms). Dominant frequency 2344–2388 Hz (average: 2358 Hz) (
Figure 9B).
Distribution: The species occurs in Rwanda, Uganda, northern Tanzania and southern Kenya [
85,
131]. Altitudinal range and province records in Rwanda: 2055–2879 m; Western and Northern Provinces.
Habitat preference and ecology: Arboreal. In swamps in montane forest, on lake shores, also in cultivated land near irrigation channels and ponds. Breeds in ponds and other bodies of stagnant water. Males call from emergent vegetation above the water surface. Maturity is reached in males within the first year of life, in females within the second. Longevity is about three years [
84].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Usually abundant, but population trend unknown. Proposed category for the National Red List: Least Concern, because the species occurs in high numbers both in montane swamps in protected areas (Volcanoes National Park) and in farmland.
3.1.24. Hyperolius jackie Dehling, 2012 (Figure 8C and Figure 9C)
Original name and description: Hyperolius jackie—J. M. Dehling (2012). Zootaxa, 3391: 52–64 [p. 54].
Type locality: “a natural pond at Karamba (S 2°28′44.28″, E 29°06′44.50″, 1940 m a.s.l.), Nyungwe National Park, Rwanda”.
External morphology: The Nyungwe Reed Frog is a small species of
Hyperolius [
80,
85]. Sexual size dimorphism absent: SVL of males 18.8–23.2 mm (average: 21.1 mm), SVL of females unknown. Reproductive males have nuptial pads and a green throat. Coloration in life: dorsum green with dark brown spots, venter transparent. Bones are blue, visible through skin. Tympanum visible, size about half the area of the eye.
Advertisement call: Males emit pulsed calls in series (intercall interval 109–132 ms depending on temperature or social interactions with conspecifics) [
80]. A single call has a duration of 54–107 ms (average: 75 ms) and consists of 10–18 pulses (average: 14) (
Figure 9C). Pulse rate on average 187 pulses/s. Pulse duration is 2–4 ms (average: 3 ms). Interpulse interval is 1–3 ms (average: 2 ms). Dominant frequency 2650–3150 Hz (average: 2956 Hz).
Distribution: Known only from the Karamba swamp and a neighbouring stream in the southwestern part of Nyungwe National Park, southwestern Rwanda. Altitudinal range and province records in Rwanda: 1881–1936 m; Western Province.
Habitat preference and ecology: Arboreal. In small swamps and near small streams in montane forest. Breeds in small ponds and probably also in stream sections with low current. Tadpole morphology and ecology are described in Ref. [
73].
Conservation status and population trend: IUCN Red List: Data Deficient [
7]. Population trend unknown. Proposed category for the National Red List: Endangered, because the type locality, and thus one of only two localities from where the species was known, was destroyed during road construction between 2014 and 2017.
3.1.25. Hyperolius kivuensis Ahl, 1931 (Figure 8D and Figure 9D)
Original name and description: Hyperolius kivuensis—E. Ahl (1931). Mitteilungen aus dem Zoologischen Museum in Berlin, 17 (1): 1–132 [p. 26].
Type locality: “Kivu-See” (=Lake Kivu).
External morphology: The Kivu Reed Frog is a large species of
Hyperolius [
2,
4,
6,
85]. Pronounced sexual size dimorphism, males 23.5–37.1 mm (average: 29.7 mm), females 25.1–40.5mm (average: 33.1 mm) [
94]. Males with gular vocal sac, in reproductive individuals colored brightly yellow. Snout long and pointed. Coloration in life: dorsum uniformly light to dark brown or green with a distinct dark lateral band, light canthal, and dorsolateral stripe often present in the brown specimens, rarely in green ones. Hidden parts of limbs reddish to yellow. Venter unpigmented in most populations. Dorsum usually without asperities.
Advertisement call: The call is brief harsh chirp [
2,
4,
6]. A single call has a duration of 50–107 ms (average: 75 ms) and includes 4–7 pulses (average: 5). Pulse duration is 1–5 ms (average: 3 ms). Interpulse interval between the first and second pulse (15–41 ms) always longer than among the following pulses (3–9 ms). Dominant frequency 2177–3124 Hz (average: 2649 Hz) (
Figure 9D). The call has been described in detail and a spectrogram of the call has been depicted elsewhere [
2,
76].
Distribution: The Kivu Reed Frog is distributed from Angola to Mozambique and northwards through Tanzania, DR Congo, Rwanda, Uganda to southwestern Ethiopia and eastern South Sudan at elevations up to 2300 m. Altitudinal range and province records in Rwanda: 1287–2143 m; all five provinces.
Habitat preference and ecology: Arboreal. In natural grassland and cultivated wetlands (marais). The species breeds in ponds and on lake shores with dense vegetation. Maturity is reached at 10–14 months, longevity in field up to four years, in captivity up to six years [
84]. Tadpole morphology and ecology are described in Refs. [
85,
132].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. One of the most common species in the wetlands of Rwanda, tolerant to human habitat alteration. Proposed category for the National Red List: Least Concern, because it occurs abundantly in natural savanna in a protected area (Akagera National Park) and in farmland throughout the country.
3.1.26. Hyperolius lateralis Laurent, 1940 (Figure 8E and Figure 9E)
Original name and description: Hyperolius lateralis—R. Laurent (1940). Revue de Zoologie et de Botanique Africaines, 34: 1–7 [p. 1].
Type locality: “N’Gesho (près des Lacs Mokoto)” (=“N’Gesho, near the Mokoto Lakes”), DR Congo.
External morphology: The Side-striped Reed Frog is a small species of
Hyperolius [
2,
4,
6,
85]. Moderate sexual size dimorphism, males 18.6–25.0 mm (average: 21.6 mm), females 21–30 mm (average: 25.3 mm). Reproductive males have a yellow throat. Coloration in life: variable patterns in males, dorsum brown to green with a dark line from nostril to eye. A dorsolateral pale white stripe is common. Venter and feet with a reddish tinge. Females dark brown or green with a broad yellow stripe, which has black margins.
Advertisement call: The call is brief harsh creak [
2,
4,
6,
76]. Males emit pulsed calls often in long series (intercall interval 0.6–1.7 s depending on temperature or social interactions with conspecifics) or singly. A single call has a duration of 13–55 ms (average: 32 ms) and contains 4–8 pulses (average: 6) [
76]. Average pulse rate is 214 pulses/s. Pulse duration is 1–4 ms (average: 3 ms). Interpulse interval is 0–3 ms (average: 1 ms). Dominant frequency 3046–3854 Hz (average: 3500 Hz) (
Figure 9E). The call has been described in detail and a spectrogram of the call has been depicted elsewhere [
2,
76,
126].
Distribution: The species is a locally common species in Burundi, DR Congo, Kenya, Rwanda, Tanzania, and Uganda in at elevations of 650–2100 m. Recent records in Rwanda are from a few scattered localities only. Altitudinal range and province records in Rwanda: 1324–2055 m; Northern, Eastern, and Southern Provinces.
Habitat preference and ecology: Arboreal. In natural or near-natural wetlands. The species breeds in small ponds and in waterfilled depressions in seasonally flooded meadows. Males reach maturity before completing the first year of life, longevity in nature is up to two years [
84]. Tadpole morphology and ecology are described in Ref. [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Locally common species in natural or moderately disturbed wetlands in Rwanda. Population trend unknown. Proposed category for the National Red List: Vulnerable, because the species has lost most of its former area of occupancy in the country and the few remaining areas where the species occurs are threatened with conversion into farmland.
3.1.27. Hyperolius parallelus Günther, 1858 (Figure 8F and Figure 9F)
Original name and description: Hyperolius parallelus—A. Günther (1858). Archiv für Naturgeschichte, 24: 319–328 [p. 326].
Type locality: “Süd-Afrika” (=“South Africa”). Later stated as “South Africa” and “Angola” [
133] and restricted to “Cape of Good Hope” and “Ambris” (=Ambriz), “Angola” [
134]. Ref. [
131] further restricted the type locality to Ambriz, Angola and assigned (by implication) the types from Cape of Good Hope to
Hyperolius marmoratus Rapp, 1842.
External morphology: The Angolan Reed Frog is a medium to large species of
Hyperolius [
85]. No marked sexual size dimorphism, males 30.0–38.4 mm (average: 35.5 mm), females slightly larger. Coloration in life variable: Dorsum usually black, dark brown or dark reddish brown with dark-edged light (white to light brown) broad stripes or large spots in the middle and on flanks. Limbs lighter colored.
Advertisement call: The call is a short metallic click. It is indistinctly pulsed, primarily in the first half. Males emit calls in series (intercall interval 0.6–1.1 s depending on temperature or social interactions with conspecifics). A single call has a duration of 60–88 ms (average: 71 ms). Dominant frequency 2534–2584 Hz (average: 2573 Hz) (
Figure 9F).
Distribution: The species is distributed from northern Namibia, through Angola, across to southern Lake Tanganyika, Burundi, Rwanda, and northern DR Congo [
131]. Only in the southwestern part of Rwanda. Altitudinal range and province records in Rwanda: 962–1839 m; Western Province.
Habitat preference and ecology: Arboreal. In savanna and cultivated wetlands; tolerant to human disturbance. Breeding habitat: ponds and ditches.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because it occurs abundantly in cultivated wetlands in southwestern Rwanda.
3.1.28. Hyperolius rwandae Dehling, Sinsch, Rödel & Channing in Channing et al., 2013 (Figure 8G and Figure 9G)
Original name and description: Hyperolius rwandae—J. M. Dehling, U. Sinsch, M.-O. Rödel & A. Channing in A. Channing, A. Hillers, S. Lötters, M.-O. Rödel, S. Schick, W. Conradie, D. Rödder, V. Mercurio, P. Wagner, J. M. Dehling, L. H. du Preez, J. Kielgast & M. Burger (2013). Zootaxa, 3620: 301–350 [p. 337].
Type locality: “Pond in farmland on the eastern outskirts of Butare, Huye District, South Province, Rwanda (2°37′10.79″ S, 29°45′08.45″ E)”.
External morphology: The Rwanda Reed Frog is a small species of
Hyperolius [
6,
82,
85]. Sexual size dimorphism absent, males 18.4–22.0 mm (average: 19.5 mm), females 18.2–23.5 mm (average: 19.2 mm) [
82,
84]. Coloration in life: Dorsum yellowish green; yellowish white, moderately broad dorsolateral stripe running along each side of the body from lateral edge of upper eyelid to groin; very small dark brown to black dots and larger brown to reddish brown specks on dorsum; ventral side and parts of dorsal side of thigh and upper arm largely unpigmented, appearing bluish-green. Iris reddish-brown during the night, yellowish-brown during the day.
Advertisement call: The call is a high-pitched chirp immediately followed by 2–4 single pulses [
6,
76,
82]. Males emit the calls in series (intercall interval 1.0–1.3 s depending on temperature or social interactions with conspecifics) or singly. A single call (long note pluse single pulses) has a duration of 62–287 ms (average: 139 ms) and consists of 13–23 pulses (average: 18) (
Figure 9G). The average pulse rate is 130 pulses/s. Pulse duration is 2–4 ms (average: 3 ms). Interpulse interval within the introductory long note ca. 1 ms, between the following single pulses 20–25 ms (average: 23 ms). Dominant frequency 4005–5272 Hz (average: 4621 Hz). The call has been described in detail elsewhere [
76,
82].
Distribution: The species is known from Rwanda, Burundi, and eastern DR Congo [
82,
85,
135]. Altitudinal range and province records in Rwanda: 1287–1807 m; all five provinces.
Habitat preference and ecology: Arboreal. Occurs in savanna, grasslands, and cultivated wetlands. It is tolerant to human impact. Breeding takes place in ponds and densely vegetated lake shores. Males call from vegetation in 5–200 cm height. Eggs are small (diameter: 1.0 mm) and placed on vegetation above the water. Maturity is reached in the second year of life at 18.2 mm; longevity in nature up to 3 years in females and 5 years in males [
84]. Size at metamorphosis is 9.0 mm. Tadpole morphology is described in [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend seems stable. Proposed category for the National Red List: Least Concern, because the species commonly occurs throughout the country in farmland and a protected area (Akagera National Park).
3.1.29. Hyperolius viridiflavus (Duméril & Bibron, 1841) (Figure 8H and Figure 9H)
Original name and description: Eucnemis viridi-flavus—A.-M.-C. Duméril & G. Bibron (1841). Erpétologie générale ou histoire naturelle complète des reptiles. Tome huitième, 784 pp. [p. 528].
Type locality: “Abyssinie” (=Ethiopia), restricted to “Adoua” (=Adwa, Ethiopia) [
136].
External morphology: The Common Reed Frog is a medium to large
Hyperolius [
6,
76,
84,
85,
99]. Moderate sexual size dimorphism, males 22.1–36.3 mm (average: 28.6 mm), females 28.4–43.0 mm (average: 30.4 mm) [
84,
94]. Reproductive males have a yellow gular disc. Coloration in life: many variations of dorsal patterns within the same population, dorsum can be uniformly greyish to brown, green with small black dots, or dark brown with yellow or whitish blotches or with longitudinal bands, or yellow with red or gray spots. The color of metamorphs is brownish to green with paired light dorsolateral lines (SVL: 13–15 mm).
Advertisement call: The call is loud metallic click [
2,
4,
6,
76]. The tonal call is emitted in long series (intercall interval 0.5–0.7 s depending on temperature or social interactions with conspecifics) or singly [
76]. A single call has a duration of 24–82 ms (average: 47 ms). Dominant frequency 2220–2917 Hz (average: 2640 Hz) (
Figure 9H). The call has been described elsewhere in detail [
76].
Distribution: It is one of the most abundant species in Burundi, DR Congo, Ethiopia, Kenya, Rwanda, Sudan, Tanzania, and Uganda at elevations of up to 2400 m. Altitudinal range and province records in Rwanda: 1287–2143 m; all five provinces. Erroneously reported from up to 3000 m in Volcanoes National Park based on misidentified
H. glandicolor [
56,
129,
130].
Habitat preference and ecology: Arboreal. Occurs in all types of savanna, grassland, bush land, and at the forest edge; tolerant to human disturbance and abundant in cultivated land. The species breeds in ponds, irrigation channels, densely vegetated lake shores areas, and in flooded meadows. Males call from vegetation at heights of 0–200 cm and compete for preferred calling sites by physical combat including kicking and grasping [
137]. Maturity is reached during the second year of life, longevity in nature up to 3 years [
84]. Tadpole morphology and ecology are described in Ref. [
85].
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because it occurs very commonly throughout the country in both farmland and a protected area (Akagera National Park).
3.1.30. Kassina senegalensis (Duméril & Bibron, 1841) (Figure 7E and Figure 10A)
Original name and description: Cystignathus Senegalensis—A.-M.-C. Duméril & G. Bibron (1841). Erpétologie générale ou histoire naturelle complète des reptiles. Tome huitième, 784 pp. [p. 418].
Type locality: “Sénégal [...] dans les étangs des environs de Galam” (=“in the ponds of Galam”, “Senegal”).
External morphology: The Senegal Running Frog is medium-sized hyperolid [
2,
4,
6]. Sexual size dimorphism absent, males and females 25–52 mm SVL. Reproductive males have a dark vocal sac, covered almost completely with the large square gular gland. Coloration in life: Dorsum yellowish brown to gray with a dark vertebral band and a pair of sometimes interrupted dark paravertebral bands, venter greyish white. Fingertips slightly swollen.
Advertisement call: The tonal call is fast rising whistle [
2,
4,
6], emitted in long series (intercall interval 2–5 s depending on temperature or social interactions with conspecifics) or singly [
76]. A single call has a duration of 58–159 ms (average: 118 ms); dominant frequency is upward-modulated (753–2174 Hz) (
Figure 7E). The call has been described elsewhere in detail [
76,
138].
Distribution: The species is distributed from Senegal through most of West Africa except the wettest coastal areas eastwards to Eritrea and southwards to southeastern South Africa, central Namibia and southwestern DR Congo at elevations from near sea level to 2500 m. It is absent from the wetter regions of Central Africa and the drier regions of the Horn of Africa. Altitudinal range and province records in Rwanda: 1287–2143 m; all five provinces.
Habitat preference and ecology: Terrestrial. The species occurs in humid and dry savannas, shrubland, forest edge, degraded forest, and cultivated wetlands. It breeds in temporary or permanent ponds. Males call at the margins of ponds, often well hidden. Males often form large choruses and respond to each other when one male begins calling.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because the species occurs in high numbers throughout the country in both farmland and natural wetlands including a protected area (Akagera National Park).
The family is represented in Rwanda by a single genus with a single species.
3.1.31. Phrynomantis bifasciatus (Smith, 1847) (Figure 10B and Figure 11H)
Original name and description: Brachymerus bifasciatus—A. Smith (1847). Illustrations of the Zoology of South Africa; consisting chiefly of figures and descriptions of the objects of natural history collected during an expedition into the interior of South Africa, in the years 1834, 1835, and 1836; fitted out by “The Cape of Good Hope Association for exploring Central Africa”. Reptilia. Part 26: Plate 63.
Type locality: “country to the east and north-east of the Cape Colony”, “on the banks of the Limpopo River, close to the tropic of Capricorn”, Limpopo Province, South Africa.
Taxonomic remark: The genetic variation among populations referred to as
Phrynomantis bifasciatus requires a re-evaluation in future [
139].
External morphology: The Banded Rubber Frog is a small to medium-sized frog species [
4,
6,
48,
140]. Sexual size dimorphism present, males up to 53 mm SVL, females up to 65 mm SVL, specimens from Rwanda smaller (up to 42 mm; [
48]). Reproductive males have a black throat, whereas that of females is greyish. Skin smooth, fingers, and toes with small, truncated discs. Coloration in life: Dorsum shows two orange to red broad bands from snout over eyes to the leg on a black background, orange to red patch above the vent. Venter grey with white small spots and blotches. Unmistakable due to its unique dorsal color pattern.
Advertisement call: The pulsed call is a long melodious trill, which can be heard from over a kilometer [
4,
6,
141,
142]. The maximum sound pressure level is 108 dB [
143]. Average call duration is 1862 ms (range: 1–7 s), intercall interval ranges from 1597 to 5004 ms. The number of pulses per call ranges from 41 to 44 pulses, pulse duration averages 22 ms (range: 20–26 ms), and the interpulse interval averages 20 ms (range: 19–23 ms). The pulse rate averages 22 pulses/s. The power spectrum includes two harmonics with almost the same intensity, the lower band ranges between 900 Hz and 1200 Hz, the upper band ranges between 1800 and 2400 Hz. A spectrogram of the call has been depicted elsewhere [
107].
Distribution: The species occurs from northern Angola to southern Kenya and southern Somalia and to northern South Africa at elevations from sea level to 1450 m. In Rwanda, known from two localities in the eastern lowlands. Altitudinal range and province records in Rwanda: 1290–1292 m; Eastern Province.
Habitat preference and ecology: Terrestrial, also arboreal. Inhabits grassland, wooded savanna, and agricultural areas such as banana plantations, tolerant to human disturbance. Breeding takes place in temporary pans and pools and in flooded grassland.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Vulnerable, because its occurrence is limited to a restricted, non-protected area in the extreme East of the country that is used for livestock breeding and agriculture.
Figure 10.
Rwandan species of Hyperoliidae (A) Kassina senegalensis, Huye (photo: J.M.D.); Microhylidae (B) Phrynomantis bifasciatus, Eastern Province (photo: Eberhard Fischer); and Phrynobatrachidae (C) Phrynobatrachus acutirostris, Nyungwe NP (photo: Eberhard Fischer), (D) Phrynobatrachus auritus, Libreville, Gabon (photo: J.M.D.), (E) Phrynobatrachus bequaerti, Rugezi Wetland (photo: J.M.D.), (F) Phrynobatrachus graueri, Nyungwe NP (photo: J.M.D.), (G) Phrynobatrachus kakamikro, Huye (photo: J.M.D.), (H) Phrynobatrachus natalensis, Huye (photo: J.M.D.).
Figure 10.
Rwandan species of Hyperoliidae (A) Kassina senegalensis, Huye (photo: J.M.D.); Microhylidae (B) Phrynomantis bifasciatus, Eastern Province (photo: Eberhard Fischer); and Phrynobatrachidae (C) Phrynobatrachus acutirostris, Nyungwe NP (photo: Eberhard Fischer), (D) Phrynobatrachus auritus, Libreville, Gabon (photo: J.M.D.), (E) Phrynobatrachus bequaerti, Rugezi Wetland (photo: J.M.D.), (F) Phrynobatrachus graueri, Nyungwe NP (photo: J.M.D.), (G) Phrynobatrachus kakamikro, Huye (photo: J.M.D.), (H) Phrynobatrachus natalensis, Huye (photo: J.M.D.).
Figure 11.
Advertisement calls of Rwandan Phrynobatrachidae (A) Phrynobatrachus acutirostris, (B) Phrynobatrachus bequaerti, (C) Phrynobatrachus kakamikro, (D) Phrynobatrachus natalensis, (E) Phrynobatrachus parvulus, (F) Phrynobatrachus scheffleri, (G) Phrynobatrachus versicolor; and Microhylidae (H) Phrynomantis bifasciatus. Calls are shown as representative spectrograms and oscillograms.
Figure 11.
Advertisement calls of Rwandan Phrynobatrachidae (A) Phrynobatrachus acutirostris, (B) Phrynobatrachus bequaerti, (C) Phrynobatrachus kakamikro, (D) Phrynobatrachus natalensis, (E) Phrynobatrachus parvulus, (F) Phrynobatrachus scheffleri, (G) Phrynobatrachus versicolor; and Microhylidae (H) Phrynomantis bifasciatus. Calls are shown as representative spectrograms and oscillograms.
Nine species of the genus Phrynobatrachus have been recorded in Rwanda so far. Morphological distinction of most species is difficult because they resemble each other in general appearance and differ mainly in adult size. All taxa are well differentiated with respect to partial 16S sequences. The advertisement call (unknown for P. auritus and P. graueri) allows for a reliable distinction of reproductive males in the field.
Key to the species of Phrynobatrachus of Rwanda based on morphological characters:
- 1
Distinct ridge from snout to eye, continued as anterior part of supratympanic fold then as dorsolateral ridge to end of trunk present ......... P. acutirostris
- 1′
Continuous ridge from snout to end of trunk absent ........ 2
- 2
Continuous, well defined, narrow concave dermal scapular ridge present ................. 3
- 2′
Dermal ridge absent or elongated tubercles present in scapular region ..... 6
- 3
Posterior part of abdomen yellow, ventral side of leg reddish orange ............. P. graueri
- 3′
Colouration of abdomen and ventral side of legs different .............4
- 4
SVL of adult males < 23 mm, adult females < 26 mm, tympanum concealed ...... P. bequaerti
- 4′
SVL of adult males and females larger, tympanum visible ................... 5
- 5
Supratympanic fold straight-lined from eye to arm insertion, lateral fold absent .... P. auritus
- 5′
Supratympanic fold rounded to angular, lateral dermal fold usually present ... P. versicolor
- 6
Tympanum covered by skin but visible, no elongated tubercles or dermal folds in scapular region, SVL of adults > 26 mm ............................. P. natalensis
- 6′
Tympanum invisible, pair of elongated large tubercles in scapular region, SVL < 25 mm ..... 7
- 7
White line from eye to arm insertion, breeding males with blackish throat and long gland on thigh ................................. P. parvulus
- 7′
Colouration of side of head and on throat different, no gland on thigh in males .... 8
- 8
Two pairs of elongated scapular tubercles forming an interrupted “W”, posterior ones almost meeting in the centre, interrupted row of enlarged tubercles on flank ............................ P. scheffleri
- 8′
Two pairs of elongated scapular tubercles forming an interrupted “X”, posterior ones separated from each other by half their length, enlarged tubercles on flank not in a row ... P. kakamikro
Key to the species of Phrynobatrachus of Rwanda based on advertisement call features:
- 1
Call consists of a single pulse group, sometimes with an additional terminal pulse ............................................................................................. 2
- 1′
Call consists of more than one pulse group ............................................ 6
- 2
Call consists of equally spaced pulses ............................................... 3
- 2′
Call with interpulse intervals of variable duration ............................ 4
- 3
Dominant frequency about 1500 Hz ................ P. natalensis
- 3′
Dominant frequency > 3500 Hz .......................... P. scheffleri
- 4
Call consists of a long pulse group followed by a well-separated single pulse ................................................................. P. kakamikro
- 4′
Call structure complex with more than 2 distinct elements ...................... 5
- 5
Call with 3 distinct elements .............................. 6
- 5′
Call with less than 3 distinct elements .............................. 7
- 6
Call duration < 500 ms, final element consisting of a single loud pulse ............................................... P. bequaerti
- 6′
Call duration > 1 s, final element consisting of 3–5 widely spaced loud pulses ...................................... P. acutirostris
- 7
Call consisting of 3–5 pulse groups, each with at least 4 pulses ................................................................. P. versicolor
- 7′
Call consisting of >10 pulse groups with a terminal loud single click ................................................................. P. parvulus
3.1.32. Phrynobatrachus acutirostris Nieden, 1912 (Figure 10C and Figure 11A)
Original name and description: Phrynobatrachus acutirostris—F. Nieden (1912 “1911”). In Schubotz H. (Ed.): Wissenschaftliche Ergebnisse der Deutschen Zentral-Afrika-Expedition 1907–1908 unter Führung Adolf Friedrichs, Herzogs zu Mecklenburg. Band IV, Zoologie II: 165–195 [p. 173].
Type locality: “Rugegewald” (=“Rugege Forest”, in the northeastern part of Nyungwe Forest, Rwanda).
External morphology: The Sharp-snouted Puddle Frog is a large species of
Phrynobatrachus with SVL of males 43.8–49.2 mm and SVL of females up to 51 mm [
6,
23,
46]. Back with characteristic inverted-u-shaped scapular ridge, lateral face separated from dorsal face by glandular ridge. Coloration in life: Dorsum uniformly brown, sometimes with dark spots, flanks darker brown. Venter brown with darker mottling.
Advertisement call: The complete advertisement call consists of three parts: (a) an initial series of pulses that are repeated with increasing rate and decreasing amplitude and fade into (b) a series of 10 or more rapidly repeated notes with constant amplitude that are finally condensed to a short pulsed note with increasing amplitude that ends with (c) a distinct last pulse of much higher amplitude and is followed by two to four separate, more or less equally spaced pulses with further increasing amplitude, the loudest part of the call (
Figure 11A). Call duration of the complete call averages 1677 ms (range: 1432–1781 ms). Dominant frequency is at 750–1000 Hz and slightly decreases from the first pulse to the pulsed note before it increases again towards the last pulse. Amplitude decreases from the first note to the penultimate pulse of the condensed note, and then significantly increases from the last pulse of the note to the end of the final pulse series (
Figure 11A). Most commonly, part c is emitted separately, especially when two or more males call near each other. The rapid series and condensed note without the louder final pulse (part b) is sometimes also emitted separately, as are, rarely, the first 3–5 pulses of the initial pulse series. This suggests that the three parts of the call may serve different purposes.
Distribution: The species is an Albertine-Rift endemic recorded from Rwanda and eastern DR Congo at elevations between 1750 and 2380 m. It probably also occurs in northwestern Burundi. Altitudinal range and province records in Rwanda: 2238–2503 m; Southern Province. Erroneously reported from Lac Ngezi in Volcanoes National Park due to a typesetting error by Ref. [
57].
Habitat preference and ecology: Terrestrial. Inhabits wetlands in pristine montane rainforest along streams, which are used for breeding. Tadpoles are found in stream sections with low current. Reproductive males call hidden below vegetation at the shore of streams. Calling activity is limited to daytime.
Conservation status and population trend: IUCN Red List: Near Threatened [
7]. Population trend decreasing. Proposed category for the National Red List: Vulnerable, due to the species’ very small geographical range.
3.1.33. Phrynobatrachus auritus Boulenger, 1900 (Figure 10D)
Original name and description: Phrynobatrachus auritus—G. A. Boulenger (1900). Proceedings of the Zoological Society of London, 1900: 433–456 [p. 440].
Type locality: “Benito River [north of the Gaboon River between 20 and 30 miles inland from the coast]”, Gabon.
External morphology: The Golden Puddle Frog is a medium-sized species of Phrynobatrachus, SVL of males up to 33 mm, that of females up to 40 mm [
6,
144]. The dorsum is smooth with a conspicuous, large X-shaped dermal ridge extending from behind the eyes to about the middle of the trunk. The canthus rostralis and the supratympanic fold are well developed. Legs long, tips of fingers and toes not enlarged.
Advertisement call: The call has been described as a “low croaking” [
6]. A quantitative call analysis is not available.
Distribution: The species is distributed from eastern Nigeria and western DR Congo to Uganda and eastern DR Congo from sea level to about 1600 m. The only record from Rwanda is based on a single specimen (RMCA 1848,
Figure 10D) collected at “Kissenyi” (=Gisenyi/Rubavu, Rwanda) in January 1926 by H. Schouteden (as
P. plicatus) [
22,
23,
145]. Laurent [
38] doubted the occurrence of this lowland-forest species at Gisenyi which he thought was improbable. No other specimen has ever been collected or observed at Gisenyi or elsewhere in Rwanda, and the presence of the species needs to be confirmed. Altitudinal range and province records in Rwanda: approximately 1500 m; Western Province.
Habitat preference and ecology: Terrestrial. Inhabits primary and secondary lowland to submontane rainforest. The species breeds in ponds, puddles, and stream sections with low current.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Data Deficient.
3.1.34. Phrynobatrachus bequaerti (Barbour & Loveridge, 1929) (Figure 10E and Figure 11B)
Original name and description: Arthroleptis bequaerti—T. Barbour & A. Loveridge (1929). Proceedings of the New England Zoological Club, 11: 25–26 [p. 25].
Type locality: “a swamp on Mt. Vissoke, Belgian Congo [=Mount Bisoke, DR Congo], 8000–9000 feet altitude”.
External morphology: Bequaert’s Puddle Frog is a very small species. Sexual size dimorphism present, SVL of females 20.0–25.8 mm, SVL of males 18.8–22.8 mm [
19]. Coloration in life: Dorsum uniformly brown, less than one-third of specimens with narrow vertebral stripe or broader vertebral band [
23]. Venter white, mottled and vermiculated with dusky brown on throat and chest.
Advertisement call: The high-pitched, pulsed call is composed of up to three parts: (a) 2–4 equally spaced pulses of low intensity, slightly increasing on the last pulse; (b) a long buzzing note consisting of 22–25 pulses with further increasing amplitude that are repeated at an increasing rate; and (c) a series of 1–4 widely separated single-pulse klick notes, the loudest part of the call (
Figure 11B). The first klick note is added after a brief interval of approximately 40 ms, the following ones after intervals of 230–250 ms. Call duration of a complete call with one klick note averages 268 ms (range: 240–277 ms). The three-part call (n = 6) was initially emitted in a spontaneous call series including 33 calls. Later, element (a) was skipped until the end of the series. The two-part call (b + c) is similar to the call of
P. kakamikro. Element duration was 13–28 ms (a), 104–190 ms (b), and 2–3 ms (c), respectively. Dominant frequency averages 3942 Hz (3936–4073 Hz). The call has not been described before.
Distribution: The species is endemic to the Albertine Rift and occurs in the DR Congo, Rwanda, and Uganda at elevations of 600–3475 m [
23,
38]. Altitudinal range and province records in Rwanda: 2055–3475 m; Northern Province.
Habitat preference and ecology: Terrestrial. Inhabits wetlands at higher altitudes, including clearings in montane forest and swamps in Dendrosenecio-Lobelia paramo. Breeds in shallow puddles and ponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend decreasing. Proposed category for the National Red List: Least Concern, because it occurs in high numbers in protected areas (Volcanoes National Park, Rugezi Wetland) and in farmland.
3.1.35. Phrynobatrachus graueri (Nieden, 1911) (Figure 10F)
Original name and description: Arthroleptis graueri—F. Nieden (1911 “1910”). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, 1910: 436–441 [p. 441].
Type locality: “Rugegewald” (=“Rugege Forest”, in the northeastern part of Nyungwe Forest, Rwanda).
External morphology: Grauer’s Puddle Frog is a small species; SVL of males up to 23 mm, SVL of females up to 28 mm [115, Dehling unpublished). Dorsum smooth with conspicuous dermal ridge on each side from eye to scapula region. Canthus rostralis and supratympanic fold well developed. Feet large with little webbing. Coloration in life: Dorsum grey to light brown with dark speckling, sometimes with a broad, yellowish vertebral stripe. Venter white with brown marbling anteriorly, orange in the posterior part. Legs dark orange to red.
Advertisement call: The call has been described as “tink-tink” or clicking [
4]. It is similar to the call of
P. bequaerti but at a lower frequency. A quantitative call analysis is not available.
Distribution: The species is endemic to the Albertine Rift, occurring in eastern DR Congo, Rwanda, Uganda, and western Kenya at elevations of 1200–3031 m [
23,
79]. Altitudinal range and province records in Rwanda: 2379–3031 m; Western and Northern Provinces.
Habitat preference and ecology: Terrestrial. The species inhabits swamps in montane forest and breeds in shallow ponds and puddles.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Near Threatened, because of the limited geographic distribution in the two protected areas (Volcanoes National Park, Nyungwe National Park).
3.1.36. Phrynobatrachus kakamikro Schick, Zimkus, Channing, Köhler & Lötters, 2010 (Figure 10G and Figure 11C)
Original name and description: Phrynobatrachus kakamikro—S. Schick, B. M. Zimkus, A. Channing, J. Köhler & S. Lötters (2010). Salamandra, 46: 24–36 [p. 29].
Type locality: “a temporary puddle at the border of the Kakamega Forest (00°21′ N, 34°51′ E; 1650 m a.s.l.), Western Province, Kenya”.
External morphology: The Kakamega Puddle Frog is among the smallest species of
Phrynobatrachus. Sexual size dimorphism present, SVL of males up to 16.2 mm, SVL of females up to 19.0 mm [
146]. Fingertips expanded, not forming discs, tympanum not visible. Coloration in life: Dorsum light brown, greyish brown to olive-brown with darker markings, sometimes with a yellowish vertebral stripe. Legs with darker cross bars. Venter whitish, throat of males greyish, chest often with darker mottling.
Advertisement call: The call is a buzz followed by a single click. The pulsed call has a duration of 400–620 ms (average: 540 ms) and consists of 15–28 pulses (average: 22.3). Intercall interval varies between 754 and 1301 ms (depending on temperature or social interactions with conspecifics). Pulse duration is 3–10 ms (average: 7 ms), interpulse interval is 5–12 ms (average: 9 ms). Pulse rate averages 62 pulses/s. Dominant frequent averages 3809 Hz (3488–4306 Hz). The call has been described elsewhere in detail [
76,
146].
Distribution: The species has been reported from western Kenya and Rwanda at elevations of 962–1807 m. It is expected to occur in southeastern Uganda and northwestern Tanzania. Altitudinal range and province records in Rwanda: 962–1807 m; all five provinces.
Habitat preference and ecology: Terrestrial. Inhabitats wetlands and the shore area of lakes. Tolerant to human habitat alteration, it is one of the most common species of cultivated wetlands and flooded grassland. The species breeds in temporary ponds, puddles, ditches, and densely vegetated lake shores.
Conservation status and population trend: IUCN Red List: Data Deficient [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because it occurs abundantly in both protected natural habitats (Akagera National Park) and cultivated areas throughout the country.
3.1.37. Phrynobatrachus natalensis (Smith, 1849) (Figure 10H and Figure 11D)
Original name and description: Stenorhynchus natalensis—A. Smith (1849). Illustrations of the Zoology of South Africa; consisting chiefly of figures and descriptions of the objects of natural history collected during an expedition into the interior of South Africa, in the years 1834, 1835, and 1836; fitted out by “The Cape of Good Hope Association for exploring Central Africa”. Reptilia. Appendix: 1–28 [p. 24].
Type locality: “The country around Port Natal” (=Durban, KwaZulu-Natal, South Africa).
Taxonomic remark: The taxon is a complex of several species [
70]. Two distinct species of the complex occur in Rwanda.
External morphology: The Natal Puddle Frog is medium-sized to large species of
Phrynobatrachus. Sexual size dimorphism weakly expressed; SVL of males up to 36 mm; SVL of females up to 38 mm [
4,
6]. Reproductive males with dark grey throat, throat of females whitish with darker spots. Fingertips slightly swollen but not expanded to discs. Dorsal skin with several large warts. Coloration in life variable. Dorsum light brown, greyish brown, light or dark grey, crème or even greenish with few small darker markings, sometimes with a yellowish vertebral stripe or band. Legs with darker cross bars. Venter whitish to light grey.
Advertisement call: The call is a slow loud snore. It is very loud and audible at distances of several hundred metres. The pulsed call has a duration of 501–821 ms (average: 632 ms) and consists of 34–58 pulses (average: 42) (
Figure 11D). Pulse duration is 6–10 ms (average: 8 ms), interpulse interval is 4–9 ms (average: 7 ms). Pulse rate averages 67 pulses/s (range: 56–80). The dominant frequent averages 1490 Hz (range: 1292–1840 Hz). The call has been described elsewhere in detail [
76].
Distribution: The nominal species is widely distributed in sub-Saharan Africa from Senegal to Eritrea and to northern Namibia and eastern South Africa in the south, from sea level to about 2000 m; absent from the wetter parts of West and Central Africa and the driest regions at the horn of Africa and in the southwestern deserts. Altitudinal range and province records in Rwanda: 1287–1785 m; all five provinces.
Habitat preference and ecology: Terrestrial. Inhabits arid and humid savanna, cultivated wetlands and agricultural areas. The species breeds in temporary ponds, puddles, and ditches. Adults are active during both day and night, but calling is largely restricted to night time and after heavy rain also during the day.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because the species occurs abundantly in farmland throughout the country as well in a protected area (Akagera National Park).
3.1.38. Phrynobatrachus sp. A
Taxonomic remark: One of two species of the
natalensis group in Rwanda (see
Section 3.1.37), a medium-sized species of
Phrynobatrachus. It differs in morphological characters, advertisement call, and 16S rRNA sequence from the other
Phrynobatrachus species from Rwanda. The taxonomic status of the species will be addressed elsewhere.
Distribution in Rwanda: So far, only known from the Western Province.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Population trend and local abundance unknown. Proposed national conservation status: Data Deficient.
Figure 12.
Rwandan species of Phrynobatrachidae (A) Phrynobatrachus parvulus, Saruhembe, Eastern Province (photo: J.M.D.), (B) Phrynobatrachus scheffleri, Akagera NP (photo: J.M.D.), (C) Phrynobatrachus versicolor, Nyungwe NP (photo: J.M.D.); and Pipidae (D) Xenopus muelleri, Bugarama (photo: J.M.D.), (E) Xenopus victorianus, Huye (photo: J.M.D.), (F) Xenopus wittei, Nyungwe NP (photo: J.M.D.).
Figure 12.
Rwandan species of Phrynobatrachidae (A) Phrynobatrachus parvulus, Saruhembe, Eastern Province (photo: J.M.D.), (B) Phrynobatrachus scheffleri, Akagera NP (photo: J.M.D.), (C) Phrynobatrachus versicolor, Nyungwe NP (photo: J.M.D.); and Pipidae (D) Xenopus muelleri, Bugarama (photo: J.M.D.), (E) Xenopus victorianus, Huye (photo: J.M.D.), (F) Xenopus wittei, Nyungwe NP (photo: J.M.D.).
3.1.39. Phrynobatrachus parvulus (Boulenger, 1905) (Figure 11E and Figure 12A)
Original name and description: Arthroleptis parvulus—G. A. Boulenger (1905). The Annals and Magazine of Natural History including Zoology, Botany, and Geology, Seventh Series, 16: 105–115 [p. 109].
Type locality: “Bange Ngola” [“Jinga country, Loanda province”] (nowadays Bange Angola, 8°26′17″ S, 16°40′11″ E, Malanje Province, Angola.
External morphology: The Little Puddle Frog is a small species of
Phrynobatrachus. Sexual size dimorphism present, males up to 16 mm SVL, rarely reaching 20 mm, females up to 25 mm SVL [
4,
6,
147]. Reproductive males with black throat and a flattened gland on the upper leg. Fingertips not expanded; tympanum usually not visible. Coloration in life: Dorsum light brown, greenish brown or dark grey, often with a yellowish vertebral stripe. Usually, a white stripe is present along the side of the head from the level of the eyes to the back edge of the jaw. Back of the thigh has a lighter band, legs with dark crossbars. Venter whitish, sometimes marked with black pattern, female throat speckled dark brown.
Advertisement call: The call is a long, buzzy, cricket-like trill, which is modified within a call series [
4,
6]. Call series start with a long call (duration up to 1064 ms) including >10 notes (duration: 32–49 ms), each one a group of 3–4 pulses with inter-pulse-group intervals of 14–28 ms. The call ends with a single loud pulse (duration: 27 ms) (
Figure 11E). In the following calls, the number of notes is successively reduced from two to zero. The end of a call series consists of up to four spaced single pulses. Dominant frequency of the pulse groups averages 4287 Hz, that of the terminal single pulses 4416 Hz. A spectrogram of a call from South Africa has been depicted elsewhere [
107].
Distribution: The species occurs from Angola to Rwanda and Burundi, and to southern Tanzania, Malawi, and northeastern Zimbabwe at elevations of 900–2400 m. Altitudinal range and province records in Rwanda: 1292–1839 m; Western and Eastern Provinces. Erroneously reported from higher elevations in Volcanoes National Park based on misidentified
P. graueri by [
129,
130].
Habitat preference and ecology: Terrestrial. Inhabits humid savannas and grasslands, montane grasslands, sometimes at the edge of forests, cultivated wetlands, and agricultural areas. Tolerant to human disturbance. Breeding habitat: grassy pools, puddles, marshes.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because it occurs adundantly in near-natural and cultivated wetlands.
3.1.40. Phrynobatrachus scheffleri (Nieden, 1911) (Figure 11F and Figure 12B)
Original name and description: Arthroleptis scheffleri—F. Nieden (1911 “1910”). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin 1910: 436–441 [p. 438].
Type locality: “bei Kibwesi in Britisch-Ostafrika” (=“near Kibwezi”, Makueni County, Kenya); “Nairobi”, Nairobi County, Kenya; “Zanzibar”, Tanzania; “Mpapua” (=Mpwapwa, Dodoma Region, Tanzania).
External morphology: Scheffler’s Puddle Frog is a tiny species of
Phrynobatrachus. Sexual size dimorphism absent, SVL of males averages 12.9 mm (range: 10.0–16.4 mm), SVL of males 15.1 mm (range: 13.0–19.1 mm) [
6,
146]. Dorsum with green rounded warts, two distinct pairs of large scapular warts. Toes webbed only at the bases. Coloration in life: Dorsum light to reddish brown with some darker markings. Top of the head in front of the eyes lighter, separated from dorsal colouration behind the eyes by a dark interorbital triangle. Legs with dark crossbars. Venter whitish with dark markings. Reproductive males with yellow or grey throat.
Advertisement call: The call is a long trill. The call has a duration of 310–2514 ms and consists of 17–108 pulses (
Figure 11F). Pulse duration is 3–5 ms (average: 4 ms), interpulse interval is 39–160 ms (average: 44 ms). Pulse rate averages 42 pulses/s. The dominantfrequency ranges between 4100 Hz and 5006 Hz). The call of a Kenyan specimen has been described elsewhere in detail [
146].
Distribution: The species occurs in Kenya, Rwanda, Tanzania, and Uganda at elevations from sea level to 1800 m. Altitudinal range and province records in Rwanda: a single locality at 1290 m; Eastern Province.
Habitat preference and ecology: Terrestrial. In herbaceous vegetation, leaf litter, rocks, or mud in moist grasslands and forest clearings. Tolerant to human disturbance. In Rwanda, only known from wooded savanna. The species breeds in the shallow shore regions of lakes, rivers, streams and pools; in Rwanda in water-filled depressions in grassland.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient. The species occurs in Akagera National Park.
3.1.41. Phrynobatrachus versicolor Ahl, 1924 (Figure 11G and Figure 12C)
Original name and description: Phrynobatrachus versicolor—E. Ahl (1924). Zoologischer Anzeiger, 61: 99–103 [p. 100].
Type locality: “Rugegewald, 2500 m” (=“Rugege Forest”, in the northeastern part of Nyungwe Forest, Rwanda).
External morphology: The Variable Puddle Frog is a medium-sized species of
Phrynobatrachus. Sexual size dimorphism present, males up to 28.4 mm SVL, females up to 34 mm SVL [
4,
6]. Dorsum smooth with few scattered tubercles. A prominent longitudinal dorsolateral dermal ridge on both sides of the trunk. Fingertips expanded to small discs, tympanum clearly visible. Breeding males with numerous pointed tubercles on back and legs. Coloration in life: Dorsum brown to greenish, sometimes with a vertebral stripe. Some individuals have a thin yellowish vertebral stripe. Legs with dark cross bars. Venter whitish with black marbling.
Advertisement call: The call is rapid series of 3–5 trills [
4,
6]. The pulsed call is usually emitted singly and has a duration of 549 ms (range: 479–705 ms). The first note is a pulse group consisting of 8–14 (average: 11) pulses and is considerably longer (range: 119–223 ms) than the following notes (
Figure 11G). These notes consist of 3–5 (average: 4) pulses and last 48–83 (average: 64) ms. The inter-pulse-group intervals are 40–50 ms. The power spectrum shows two harmonics with almost the same intensity: the lower band ranges between 750 Hz and 950 Hz, the upper one between 1550 and 1850 Hz.
Distribution: The species is endemic to the Albertine Rift, occurring in Burundi, DR Congo, Rwanda, and Uganda at elevations of 1500–2379 m. Altitudinal range and province records in Rwanda: 1961–2379 m, in Volcanoes, Gishwati-Mukura, and Nyungwe National Parks; Northern, Western, and Southern Provinces.
Habitat preference and ecology: Semi-aquatic. Inhabits swamps and wetlands in montane forests. The species breeds in swamp pools. Males call from vegetation submerged in water.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend decreasing, locally abundant. Proposed category for the National Red List: Near Threatened, because of the limited geographic distribution in the country.
3.1.42. Phrynobatrachus sp. B
Taxonomic remark: A medium-sized species of Phrynobatrachus. It differs in morphological characters, advertisement call, and 16S rRNA sequence from the other Phrynobatrachus species from Rwanda. The taxonomic status of the species will be addressed elsewhere (Dehling et al., in preparation).
Distribution in Rwanda: So far, only known from the Western Province.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Population trend and local abundance unknown. Proposed national conservation status: Data Deficient.
Four species of the genus Xenopus have been recorded from Rwanda. The presence of one species (X. vestitus) requires confirmation. External morphology, especially the dorsal color pattern and the ventral coloration in life, allows for a reliable distinction. The advertisement call is emitted underwater, and calls differ markedly between the species. Moreover, the species are genetically well differentiated.
Key to the species of Xenopus of Rwanda based on morphological characters:
- 1
Dorsum yellowish light brown with distinct large olive blotches ..................... X. muelleri
- 1′
Dorsum more or less uniformly coloured, without distinct large blotches ............ 20
- 2
Throat and lateral sides of venter grey to brown, contrasting with whitish central venter .... X. vestitus
- 2′
Throat and lateral sides of venter not differing in colour with rest of venter ................ 3
- 3
Venter yellow with small black spots ............................ X. victorianus
- 3′
Venter yellow with small faint black spots, throat and chest whitish .......... X. wittei
Key to the species of Xenopus of Rwanda based on advertisement call features:
- 1
Call a single long note, consisting of >20 pulses increasing in amplitude ...... 2
- 1′
Call containing several distinct short notes, each usually consisting of <15 pulses of constant amplitude .............................................................. 3
- 2
Dominant frequency at about 2000 Hz; note lasting 0.2 ms to 1.2 s, consisting of 26–114 pulses that are repeated at a rate of 76–116 pulses/s .................................................................................X. vestitus
- 2′
Dominant frequency at 1300–1500 Hz; note lasting 0.29 ms to more than 3 s, consisting of 9 to more than 100 pulses that are repeated at a rate of 30–40 pulses/s ....................................................................... X. wittei
- 3
Notes consisting of 2–6 pulses, notes repeated at 9/s ...... X. victorianus
- 3′
Notes consisting of (4)6–12 pulses, repeated at a rate of 1.5–3.4/s ................................................................................ X. muelleri
Figure 13.
Advertisement calls of Pipidae in Rwanda. (
A)
Xenopus muelleri, (
B)
Xenopus victorianus, and (
C)
Xenopus wittei (call recordings by Ref. [
68]). Calls are shown as representative spectrograms and oscillograms.
Figure 13.
Advertisement calls of Pipidae in Rwanda. (
A)
Xenopus muelleri, (
B)
Xenopus victorianus, and (
C)
Xenopus wittei (call recordings by Ref. [
68]). Calls are shown as representative spectrograms and oscillograms.
3.1.43. Xenopus muelleri (Peters, 1844) (Figure 12D and Figure 13A)
Original name and description: Dactylethra Muelleri—[W. C. H.] Peters (1844). Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin. Aus dem Jahre 1844: 31–37 [p. 37].
Type locality: “Mozambique”. Later restricted to “Cabaceira, Boror, Sena, Tette” by Ref. [
148], and further restricted to “Tete, Zambesi River, Mozambique” by Ref. [
149].
External morphology: Müller’s Clawed Frog is a large species of
Xenopus. Pronounced sexual size dimorphism with SVL of males SVL up to 61 mm, that of females up to 90 mm [
4,
6]. Three claws on the foot. Head relatively small; snout rounded. Eyes moderately large. Subocular tubercle long, two-thirds of eye diameter. Coloration in life: Dorsum yellowish light brown with several large olive blotches and many smaller olive or dark brown spots. Venter yellowish brown with few black spots.
Advertisement call: Males produce series of notes with 6–12 (rarely 4) pulses that are repeated at a rate of 1.5–3.4/s; other vocalisations such as a series of rapidly repeated double-pulsed notes have been reported [
68,
150,
151].
Distribution: The species is distributed from eastern DR Congo and Rwanda to southern Kenya, south to northeastern Namibia and eastern South Africa at elevations from sea level to 2000 m. In Rwanda, only known from the Bugarama region in the extreme Southwest of the country. Altitudinal range and province records in Rwanda: 962 m; Western Province.
Habitat preference and lifestyle: Aquatic. Inhabits many kinds of stagnant water bodies, from small puddles to the shore area of lakes in open habitats including savanna and farmland. Tolerant to human disturbance.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because although its distribution in Rwanda is very restricted it ocurs in high numbers in near natural habitats and farmland.
3.1.44. Xenopus vestitus Laurent, 1972
Original name and description: Xenopus vestitus—R. F. Laurent (1972). Exploration du Parc National des Virunga. Deuxième série, 22: 1–125 [p. 9].
Type locality: “riv. Musugereza, près de Rutshuru, +/−1.250 m, Kivu, Congo” (=“Musugereza River, near Rutshuru, ±1250 m, Kivu, Congo” [nowadays North Kivu Province, DR Congo]).
External morphology: The Jacketed Clawed Frog is a relatively small species of
Xenopus. SVL of males to 47 mm, that of females to 55 mm [
152,
153]. Three claws on the foot. Head wedge-shaped or pointed, snout protruding. Eyes relatively small. Subocular tubercle short, one-third of eye diameter. Coloration in life: Dorsum, throat, and lateral sides of venter grey to brown with a marbling of light silver-golden to bronze chromatophores [
38,
154]. Chest, central part of venter, and ventral side of legs whitish.
Advertisement call: A long note lasting 0.2–1.2 s and consisting of 26–114 pulses of steadily increasing amplitude repeated at 76–116 pulses/s. The dominant frequency is at about 2000 Hz [
66,
150].
Distribution: The species is endemic to the Albertine Rift and distributed from eastern DR Congo through southwestern Uganda to northeastern Rwanda at elevations of 925–2100 m. In Rwanda, the species has been reported from two localities; a single specimen collected at Gabiro by Burgeon in 1932, together with two
X. victorianus [
152], and two males recorded at Shama [
150]. Altitudinal range and province records in Rwanda: 1377–1772 m; Eastern Province.
Habitat preference and lifestyle: Aquatic. Inhabits montane forests and grassland. The species breeds in ponds and other kinds of stagnant water bodies. Tolerant to human disturbance, it is also found in cultivated areas.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient.
3.1.45. Xenopus victorianus Ahl, 1924 (Figure 12E and Figure 13B)
Original name and description: Xenopus victorianus—E. Ahl (1924). Zoologischer Anzeiger, 60: 269–273 [p. 270].
Type locality: “Bussissi, am Victoria-See” (=“Bussissi, at Lake Victoria”) [nowadays Busisi, 2°43′ S, 32°51′ E], Tanzania.
External morphology: The Lake Victoria Clawed Frog is large species of
Xenopus. Pronounced sexual size dimorphism; SVL of males up to 60 mm, SVL of females up to 78 mm [
4,
6,
154]. Three claws on the foot. Head large and rounded. Eyes relatively large. Subocular tubercle short, one-third of eye diameter. Coloration in life: Dorsum yellowish brown to grey with many small spots and few irregular larger spots. Venter yellow with small black spots.
Advertisement call: Males produce calls of the click and burst type, which are emitted underwater [
68]. The call is a series of short notes, each lasting 20–50 ms and consisting of 2–6 pulses. The notes are rapidly repeated at a rate of 9/s [
4].
Distribution: The species is distributed from southern South Sudan and eastern DR Congo, throughout Uganda, Rwanda and Burundi, to eastern Kenya and northern and eastern Tanzania at elevations from sea leavel to about 2700 m. It is the most common Clawed Frog in Rwanda. The species occurs in Akagera National Park [
155]. Altitudinal range and province records in Rwanda: 1287–1839 m; all five provinces.
Habitat preference and ecology: Aquatic. Inhabits open habitats such as savanna, natural and cultivated wetlands and farmland. The species breeds in stagnant waterbodies from puddles to the shore area of lakes, and also in sections of streams with no or low current. Tolerant to human disturbance, very frequently found in eutrophic fishponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend increasing. Proposed category for the National Red List: Least Concern, because the species occurs abundantly throughout the country in cultivated areas as well as in natural wetlands including a protected area (Akagera National Park).
3.1.46. Xenopus wittei Tinsley, Kobel & Fischberg, 1979 (Figure 12F and Figure 13C)
Original name and description: Xenopus wittei—R. C. Tinsley, H. R. Kobel & M. Fischberg (1979). Journal of Zoology 188: 69–102 [p. 73].
Type locality: “Chelima Forest (waterhole alongside Kabale—Rutenga road), Kigezi District, S.W. Uganda; alt. 2200 m. 1°04′ S, 29°55′ E”.
External morphology: de Witte’s Clawed Frog is medium-sized species of
Xenopus. Moderate sexual size dimorphism, SVL of males up to 50 mm, SVL of females up to 61 mm [
4,
153,
154].
Three claws on the foot. Head relatively wide and snout rounded. Eyes large. Subocular tubercle short, one-third of eye diameter. Coloration in life: Dorsum uniformly greyish to yellowish brown. Venter yellow with small faint black spots, throat and chest whitish.
Advertisement call: Males produce calls of the trill type, which are emitted underwater [
68]. The call consists of a single note that lasts from 0.29 s to more than 3 s and consists of 9 to more than 100 pulses that are repeated at a rate of 30–40/s and initially slowly increase in amplitude but are repeatedly and regularly downward- and again upward-modulated in the second half of longer calls; the dominant frequency is at 1300–1500 Hz [
66,
150].
Distribution: The species is endemic to the Albertine Rift and occurs in eastern DR Congo, southwestern Uganda, Rwanda, and Burundi at elevations 1200–2379 m Altitudinal range and province records in Rwanda: 1936–2379 m; Northern, Western, and Southern Provinces.
Habitat preference and ecology: Aquatic. Inhabits montane swamps and lakes within forest, grasslands, and cultivated wetlands. Tolerant to human disturbance. The species breeds in bodies of standing water from large puddles to ponds and lakes.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because the species occurs in high numbers in both protected areas (Volcanoes, Gishwati-Mukura, and Nyungwe National Parks) and cultivated wetlands and farmland.
Nine species of the genus
Ptychadena have been recorded so far in Rwanda. The identity and taxonomic status of two species recorded recently during the preparation of the manuscript will be addressed elsewhere (Dehling, in preparation). We only treat in detail herein the seven other species. These can be distinguished based on external morphological characters as well as characteristics of their advertisement calls [
87]. All taxa are well differentiated in the sequence of the mitochondrial 16S gene.
Key to the species of Ptychadena of Rwanda based on morphological characters:
- 1
Median dorsal ridge reaching onto snout ........................ P. uzungwensis
- 1′
Median dorsal ridge not reaching beyond level of eyes ..................... 2
- 2
Webbing on Toe IV reaching to distal subarticular tubercle or beyond ..... 3
- 2′
Webbing on Toe IV reaching to penultimate subarticular tubercle or slightly beyond .......... 4
- 3
SVL of adult males > 49 mm, of adult females 58.0 mm, internarial distance less than distance from nostril to tip of snout, postaxial side of thigh with yellow mottling .......... P. oxyrhynchus
- 3′
SVL of adult males < 44 mm, of adult females < 52 mm, internarial distance equal to distance from nostril to tip of snout, postaxial side of thigh with reticulated longitudinal yellow bands ... P. anchietae
- 4
Postaxial side of thighs with yellow spots, inner metatarsal tubercle large, more than half the length of metatarsus of first toe ..................... P. porosissima
- 4′
Postaxial side of thighs with bright longitudinal bands, inner metatarsal tubercle small, less than half the length of metatarsus of first toe .................... 5
- 5
Ventral side of head white, sometimes with grey mottling, webbing reaching beyond distal subarticular tubercle on first toe and on postaxial side of fourth toe, vocal sac aperture in males superior ...................................................... P. nilotica
- 5′
Ventral side of head yellow, webbing not reaching beyond distal subarticular tubercle on first toe and on postaxial side of fourth toe, vocal sac aperture in males inferior .................................... 6
- 6
SVL of adult males > 37.7 mm, of adult females 48.0–57.7 mm, postaxial side of thighs with irregularly shaped bright bands .............................. P. chrysogaster
- 6′
SVL of adult males < 37.6 mm, of adult females 43.6–49.0 mm, postaxial side of thighs with sharply defined stripes .......................................... P. guibei
Key to the species of Ptychadena of Rwanda based on advertisement call features:
- 1
Call with three notes ................................................................... 2
- 1′
Call with a single note ................................................................ 3
- 2
Amplitude of notes increases from the first to the third, dominant frequency > 4000 Hz .................................................................. P. guibei
- 2′
Amplitude of notes remains at the same level, dominant frequency < 2500 Hz .................................................................... P. chrysogaster
- 3
Pulse repetition rate < 25 ...................................... P. oxyrhynchus
- 3′
Pulse repetition rate > 36 .............................................................. 4
- 4
Fundamental frequency < 800 Hz ................................. P. nilotica
- 4′
Fundamental frequency > 1100 Hz ................................................. 5
- 5
Call without marked frequency modulation, repeated at high rate (>150/m) in long series, pulse repetition rate > 95/s .................... P. uzungwensis
- 5′
Call with marked dominant-frequency modulation, cricket-like, repeated at lower rate (<120/m), pulse repetition rate < 85/s ................................. 6
- 6
Pulses widely separated, interpulse interval 13.2 [7–20] ms, fundamental frequency > 1800 Hz, dominant frequency usually <3600 [2971–3724] Hz ...........................................................................P. anchietae
- 6′
Pulses not widely separated, interpulse interval 5.5 [3–8] ms, fundamental frequency < 1700 Hz, dominant frequency usually >3700 [3617–4306] Hz ...................................................................... P. porosissima
Figure 14.
Rwandan species of Ptychadenidae (A) Ptychadena anchietae, Huye (photo: J.M.D.), (B) Ptychadena chrysogaster, Rugezi Wetland (photo: J.M.D.), (C) Ptychadena guibei, Bugarama (photo: Mapendo Mindje), (D) Ptychadena nilotica, Mugesera Wetland (photo: J.M.D.), (E) Ptychadena oxyrhynchus, Akagera NP (photo: J.M.D.), (F) Ptychadena porosissima, Huye (photo: J.M.D.), (G) Ptychadena uzungwensis, Huye (photo: J.M.D.); and Pyxicephalidae, (H) Cacosternum plimptoni, Akagera NP (photo: J.M.D.).
Figure 14.
Rwandan species of Ptychadenidae (A) Ptychadena anchietae, Huye (photo: J.M.D.), (B) Ptychadena chrysogaster, Rugezi Wetland (photo: J.M.D.), (C) Ptychadena guibei, Bugarama (photo: Mapendo Mindje), (D) Ptychadena nilotica, Mugesera Wetland (photo: J.M.D.), (E) Ptychadena oxyrhynchus, Akagera NP (photo: J.M.D.), (F) Ptychadena porosissima, Huye (photo: J.M.D.), (G) Ptychadena uzungwensis, Huye (photo: J.M.D.); and Pyxicephalidae, (H) Cacosternum plimptoni, Akagera NP (photo: J.M.D.).
Figure 15.
Advertisement calls of Ptychadenidae in Rwanda. (A) Ptychadena anchietae, (B) Ptychadena chrysogaster, (C) Ptychadena guibei, (D) Ptychadena nilotica, (E) Ptychadena oxyrhynchus, (F) Ptychadena porosissima, (G) Ptychadena uzungwensis. Calls are shown as representative spectrograms and oscillograms.
Figure 15.
Advertisement calls of Ptychadenidae in Rwanda. (A) Ptychadena anchietae, (B) Ptychadena chrysogaster, (C) Ptychadena guibei, (D) Ptychadena nilotica, (E) Ptychadena oxyrhynchus, (F) Ptychadena porosissima, (G) Ptychadena uzungwensis. Calls are shown as representative spectrograms and oscillograms.
3.1.47. Ptychadena anchietae (Bocage, 1867) (Figure 14A and Figure 15A)
Original name and description: Rana anchietae—J. V. B. du Bocage (1867). Proceedings of the scientific meetings of the Zoological Society of London for the year 1867: 843–846 [p. 843].
Type locality: “Benguella”, (=Benguela, Benguela Province, Angola).
External morphology: Anchieta’s Ridged Frog is a medium-sized species of
Ptychadena. Pronounced sexual size dimorphism, SVL 38.0–42.4 mm in adult males, 46.7–51.3 mm in adult females [
4,
88,
89,
90]. Vocal sac aperture in males inferior; internarial distance equal to distance from nostril to tip of snout; inner metatarsal tubercle about half the length of metatarsus I; toe webbing I0.5/2II0.5/2III0.5,1/2IV2-/0.5V; reproductive males lack spines on venter. Coloration in life: Ventral side of head white, of trunk yellow; dark brown stripe on preaxial side of tibia absent; light tibial line absent; light medial dorsal band absent; light, prominent dorsolateral fold usually absent; postaxial side of femur with irregularly delimited, reticulated, longitudinal bands, alternately yellow and dark brown coloured.
Advertisement call: The call is a high-pitched short trill emitted in short series [
4,
88]. The pulsed call has a duration of 110–278 ms (average: 203 ms) and consists of 7–14 pulses (average: 11) (
Figure 15A). Intercall interval varies between 274 and 3072 ms (depending on temperature or social interactions with conspecifics). Pulse duration is 8–13 ms (average: 11 ms), interpulse interval is 3–8 ms (average: 6 ms). Pulse rate averages 54 pulses/s. The power spectrum shows two harmonics with almost the same intensity, the lower band ranges between 1480 and 1691 Hz, the upper one between 2971 and 3724 Hz. The call has been described in detail and spectrograms of the call have been depicted elsewhere [
88,
90,
107].
Distribution: The species is distributed from Angola to eastern DR Congo, South Sudana and Eritrea, south to Namibia and South Africa at elevations from near sea level to up to 2143 m. Altitudinal range and province records in Rwanda: 1287–2143 m; all five provinces.
Habitat preference and ecology: Terrestrial. Inhabits savanna, grassland, natural and cultivated wetlands, clearings in forests, and farmland. The species breeds in temporary ponds and puddles in savanna and in ditches, puddles, ruts, and rice fields in farmland.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because of its occurrence in high numbers in both a protected area (Akagera National Park) and in agricultural wetlands (marais) throughout the country.
3.1.48. Ptychadena chrysogaster Laurent, 1954 (Figure 14B and Figure 15B)
Original name and description: Ptychadena chrysogaster—R. Laurent (1954). Annales du Musée Royal du Congo Belge. Sciences Zoologiques, 34: 1–34 [p. 18].
Type locality: “Lac Karago, 2.250 m., Terr. de Kisenyi, Ruanda” (=“Lake Karago, 2250 m, Gisenyi Territory, Rwanda”).
External morphology: The Yellow-bellied Ridged Frog is a medium-sized species of
Ptychadena. Pronounced sexual size dimorphism, SVL of males 37.8–49.7 mm (average: 43.3 mm), SVL of males 48.0–57.7 mm (average: 53.7 mm) [
4,
6,
89,
90]. Vocal sac aperture in males is inferior; inner metatarsal tubercle less than half the length of metatarsus I; toe webbing I2/2.5II1.5,1.75/3III2,2-/3.25,3+IV3/1.5,2V; reproductive males have smooth skin covered with minute spines. Coloration in life: Ventral side of head and trunk yellow; dark brown stripe on preaxial side of tibia, continuous or almost continuous; light tibial line usually present, rarely absent; light medial dorsal band usually present, rarely absent; light, prominent dorsolateral fold present; postaxial side of femur with irregularly delimited, reticulated, longitudinal dark bands on greenish-yellow background.
Advertisement call: The call consists of three pulsed notes. The complete call has a duration of 164–189 ms (average: 177 ms). The intercall interval varies between 796 and 2558 ms (depending on temperature or social interactions with conspecifics). The note structure is uniform, with a duration of 30–45 ms (average: 37 ms), 3–6 pulses per note (average: 5), and an internote interval of 27–34 ms (average: 29 ms) (
Figure 15B). Pulse duration is 4–5 ms (average: 5 ms), interpulse interval is 0–3 ms (average: 2 ms). Pulse rate averages 82 pulses/s. The fundamental frequency is at 1136–1195 Hz and the dominant frequency at 2368–2411 Hz [
90].
Distribution: The species is endemic to the Albertine Rift and is distributed from eastern DR Congo and southwestern Uganda to western Rwanda and Burundi at elevations of 1200–2600 m. Altitudinal range and province records in Rwanda: 1650–2600 m; formerly widely distributed in Northern, Western, Eastern, and Southern Provinces, but the species has lost much of its former range in Rwanda [
89]. Recently recorded from scattered localities in Northern, Western, and Southern Provinces.
Habitat preference and ecology: Terrestrial. Inhabits montane wetlands at the edge of forest. The species does not survive in intensively cultivated areas. Breeding occurs in ponds, puddles, and seasonally flooded meadows.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Endangered, because of a massive population decline during the last 60 years [
89]. The species has lost most of its former range in Rwanda and is now restricted to few scattered and isolated localities that are threatened with habitat alteration. Recorded from Volcanoes National Park and from the edges of Gishwati-Mukura and Nyungwe National Parks.
3.1.49. Ptychadena guibei Laurent, 1954 (Figure 14C and Figure 15C)
Original name and description: Ptychadena chrysogaster guibei—R. Laurent (1954). Annales du Musée Royal du Congo Belge. Sciences Zoologiques, 34: 1–34 [p. 23].
Type locality: “Muita, Luembe E, Angola” (=Muíta, 7.48° S, 21.27° E, 800 m [
35]; Lunda Norte Province, Angola).
External morphology: Guibe’s Ridged Frog is a medium-sized species of
Ptychadena. Pronounced sexual size dimorphism, SVL 35.5–37.5 mm in adult males, 43.6–49.0 mm in adult females [
90,
107,
156]. Vocal sac aperture in males inferior; inner metatarsal tubercle less than half the length of metatarsus I; toe webbing I2/2.5II1.5,1.75/2.75,3,3.25III2-,2/3IV3/1.5,1.75V; reproductive males have smooth skin covered with very small spines. Coloration in life: Ventral side of head and trunk yellow; series of dark brown blotches on preaxial side of tibia; light tibial line usually present, rarely absent; light medial dorsal band present; light, prominent dorsolateral fold present; postaxial side of femur with three sharply delimited longitudinal, bright greenish yellow stripes on dark brown to blackish background.
Advertisement call: The call consists of three pulsed notes. The complete call has a duration of 52–128 (average: 124 ms). The intercall interval varies between 399 and 1027 ms (depending on temperature or social interactions with conspecifics). Note duration increases from first (average 22.6 ms) to third note (35.6 ms), alongside an increase in number of contained pulses (4–5 in the first note, 10–12 in the third note) (
Figure 15C). Internote interval of 11–13 ms. Pulse duration is 2–5 ms (average: 3 ms). Pulse repetition rate within a note averages 300 pulses/s. The fundamental frequency is at 1828–2296 Hz and the dominant frequency is at 4284–4970 Hz [
90].
Distribution: The species is distributed from Angola to northern Zambia and northern Mozambique at elevations from near sea level to about 1400 m. It was recently recorded from Rwanda where it is known from a single locality [
90]. Altitudinal range and province records in Rwanda: 962 m; Western Province.
Habitat preference and lifestyle: Terrestrial. Inhabits savanna and other grassland, also in cultivated wetlands and farmland. The species breeds in stagnant water bodies including puddles, ditches, rice fields, and ponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient.
3.1.50. Ptychadena nilotica (Seetzen, 1855) (Figure 14D and Figure 15D)
Original name and description: Rana nilotica—U. J. Seetzen (1855). Reisen durch Syrien, Palästina, Phönicien, die Transjordan-Länder, Arabia Petraea und Unter-Aegypten. Dritter Band [p. 490].
Type locality: “Kahira” (=Cairo, Egypt).
External morphology: The Nile Ridged Frog is a large species of
Ptychadena. Pronounced sexual size dimorphism, SVL 37.2–45.2 mm adult in males, 45.6–53.1 mm in adult females [
88,
89,
90]. Vocal sac aperture in males superior; inner metatarsal tubercle less than half the length of metatarsus I; toe webbing I1.5,1.75/2,2.25II1.5/2.75,3III1.75,2/3IV2.75/1,1.5V; reproductive males lack ventral spines. Coloration in life: Ventral side of head white, mottled with grey; abdomen yellow; dark brown stripe on preaxial side of tibia absent in most specimens, few with dark mottling, not forming continuous stripe; light tibial line present or absent; light medial dorsal band present or absent; light, prominent dorsolateral fold present; postaxial side of femur with relatively sharply delimited longitudinal bands, alternately yellow and black coloured.
Advertisement call: The call consists of a single pulsed note, emitted in series. It has a duration of 72–281 ms (average: 163 ms) and consists of 8–26 pulses (average: 16) (
Figure 15D). Intercall interval varies between 95 and 2504 ms (depending on temperature or social interactions with conspecifics). Pulse duration is 3–7 ms (average: 5 ms), interpulse interval is 2–6 ms (average: 4 ms). Pulse rate averages 96 pulses/s. The fundamental frequency is at 599–754 Hz, and the dominant frequency at 1722–2712 Hz. The call has been described in detail and a spectrogram of the call has been presented elsewhere [
76,
88,
90].
Distribution: The species is distriubuted from the mouth of the Nile River at Cairo along the Nile to Rwanda and Burundi and to eastern DR Congo, Kenya, Tanzania, and western Zambia at elevations from near sea level up to 2143 m. Altitudinal range and province records in Rwanda: 962–2143 m; all five provinces.
Habitat preference and ecology: Terrestrial. Inhabits natural wetlands along the Nile River including the lake system of the Akagera. Tolerant to human disturbance and one of the most common species in cultivated wetlands, rice fields, and grassland with secondary tall herbaceous vegetation near permanent water bodies. The species breeds in still and slow-flowing water bodies, e.g., puddles, ditches, ponds, rice fields, and densely vegetated shore areas of lakes.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because of its occurrence in high numbers in both a protected area (Akagera National Park) and in agricultural wetlands (marais) throughout the country.
3.1.51. Ptychadena oxyrhynchus (Smith, 1849) (Figure 14E and Figure 15E)
Original name and description: Rana oxyrhynchus—A. Smith (1849). Illustrations of the Zoology of South Africa; consisting chiefly of figures and descriptions of the objects of natural history collected during an expedition into the interior of South Africa, in the years 1834, 1835, and 1836; fitted out by “The Cape of Good Hope Association for exploring Central Africa”. Reptilia. [Part 28: Plate 77, 2–2c].
Type locality: “Kaffir Land and the country about Port Natal” (=Eastern Cape Province and region of Durban, South Africa).
External morphology: The Sharp-nosed Ridged Frog is one of the largest species of
Ptychadena. Pronounced sexual size dimorphism, SVL 49.2–52.7 mm in adult males, 58.0 mm in adult females [
4,
6,
90,
107]. Vocal sac aperture in males inferior; inner metatarsal tubercle about half the length of metatarsus I; toe webbing I0.5,1/1.75,2-II0.5/2+,2.25III0.5/2.25IV2-,2/0,0.5V; reproductive males lack ventral spines. Coloration in life: Ventral side of head white, ventral side of trunk yellow; dark brown stripe on preaxial side of tibia absent; light tibial line absent; light medial dorsal band absent; light prominent dorsolateral fold absent, but postpalpebral fold white and prominent in its posterior fourth; postaxial side of femur with irregularly reticulated mottling.
Advertisement call: The call is a moderately low-pitched trill, consists of a single pulse group [
4,
90,
107]. It is usually emitted in series comprising 10 to 19 calls, separated by intervals of 602–1801 ms. Series start with a call consisting of 5–6 pulses and having a slightly lower amplitude than the following calls. At the end of a series, calls contain up to 10 pulses. Pulse repetition rate is 17–23/s. Pulse amplitudes within a call increase from the first pulse to reach its maximum in the central pulses and decreases again at the last two pulses (
Figure 15D). Correspondingly, the dominant frequency increases from the first pulse to the central pulses and slightly decreases at the last pulses. The fundamental frequency is the dominant frequency, at 1722–2712 Hz.
Distribution: The species is widely distributed in sub-Saharan Africa, from Senegal to Kenya, south to southern Republic of the Congo and eastern South Africa at elevations from near sea level to about 2000 m. Altitudinal range and province records in Rwanda: 1326–ca. 1700 m; Eastern Province.
Habitat preference and ecology: Terrestrial. Inhabits dry and moist savanna and other grasslands. The species breeds in small permantent and temporary ponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because of its frequent occurrence in a protected area (Akagera National Park).
3.1.52. Ptychadena porosissima (Steindachner, 1867) (Figure 14F and Figure 15F)
Original name and description: Rana porosissima—F. Steindachner (1867). Reise der österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter den Befehlen des Commodore B. von Wüllerstorf-Urbair. Zoologischer Theil. Erster Band. Amphibien. 70 pp, 5 plates [p. 18, Plate 9,
Figure 9,
Figure 10,
Figure 11,
Figure 12 and
Figure 13].
Type locality: “Angola”.
External morphology: The Striped Ridged Frog is a medium-sized species of
Ptychadena. Pronounced sexual size dimorphism, SVL 37.3–44.5 mm in adult males, 39.0–52.1 mm in adult females [
4,
6,
88,
89,
90]. Vocal sac aperture in males inferior; inner metatarsal tubercle more than half the length of metatarsus I; toe webbing I1.75,2/2.25II1.5/3III1.75/3,3.25IV3/1,1.5V; reproductive males with many large ventral spines. Coloration in life: Ventral side of head and trunk yellow; dark brown stripe on preaxial side of tibia absent; light tibial line present; light medial dorsal band present or absent; light, prominent dorsolateral fold present; postaxial side of femur with yellow spots diffusely arranged in longitudinal rows on dark brown background.
Advertisement call: The call is a high-pitched trill emitted in series [
4,
88]. The pulsed call has a duration of 100–262 ms (average: 188 ms) and consists of 5–13 pulses (average: 10) (
Figure 15F). Intercall interval varies between 116 and 996 ms (depending on temperature or social interactions with conspecifics). Pulse duration is 3–12 ms (average: 6 ms), interpulse interval is 7–20 ms (average: 13 ms). Pulse rate averages 53 pulses/s. The fundamental frequency is at 1808–2067 Hz and contains almost as much energy as the dominant frequency at 3617–4306 Hz. The call has been described in detail and a spectrogram of the call has been presented elsewhere [
88,
90,
107].
Distribution: The species is distributed from Angola to Uganda and Kenya, and south to Zimbabwe and eastern South Africa at elevations of 1200–1900 m. Altitudinal range and province records in Rwanda: 1292–1807 m; Kigali, Eastern, and Southern Provinces.
Habitat preference and ecology: Terrestrial. Inhabits high-altitude swamps, moist grassland, and savanna. Tolerant to human disturbance, commonly found in cultivated wetlands. The species breeds in ponds, ditches, and rice fields.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Least Concern, because of its occurrence in high numbers in agricultural wetlands (marais) throughout the country.
3.1.53. Ptychadena uzungwensis (Loveridge, 1932) (Figure 14G and Figure 15G)
Original name and description: Rana mascareniensis uzungwensis—A. Loveridge (1932). Bulletin of the Museum of Comparative Zoology at Harvard College, 72: 375–387 [p. 384].
Type locality: “Dabaga, Uzungwe Mountains, Tanganyika Territory” (=Dabaga, Iringa Region, Tanzania).
External morphology: The Udzungwa Ridged Frog is a small species of
Ptychadena. Pronounced sexual size dimorphism, SVL 33.3–35.7 mm in adult males, 43.3 mm in adult females [
4,
6,
89,
90]. Vocal sac aperture in males semi-inferior; inner metatarsal tubercle about half the length of metatarsus I; toe webbing I2/2.25,2.5II1.5/3-III1.75,2-/3IV3/1+,1.25V; reproductive males have very small ventral spines. Coloration in life: Ventral side of head and abdomen yellow, breast yellowish white ventrally; dark brown stripe on preaxial side of tibia absent; light tibial line absent; light medial dorsal band present; light, prominent dorsolateral fold present; postaxial side of femur with two faint, longitudinal yellow bands, widened to irregularly delimited yellow blotches distally, on greyish dark brown background.
Advertisement call: The call is a moderately high-pitched trill, repeated in longer series that sound to the human ear like stuttered baaing [
4,
90]. A call series consists typically of up to 40 calls, repeated at a rate of 94–190/min. The single call lasts 154–585 (average: 310) ms and consists of 17–79 (average: 38) pulses, repeated at a rate of 97–162 pulses/s (
Figure 15G). A spectrogram of the call has been depicted elsewhere [
90,
107].
Distribution: The species is distributed from Angola to Rwanda and south to Mozambique and Zimbabwe at elevations of 800–2300 m. Altitudinal range and province records in Rwanda: 1643–2000 m; Southern Province.
Habitat preference and ecology: Terrestrial. Inhabits grassland, near pools or seepages, also in farmland. The species breeds in temporary pools and rice fields.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Vulnerable, because the species has lost about half of its former area of occupancy in the country within the past 50 years and cannot survive in intensively cultivated areas.
3.1.54. Ptychadena sp. A
Taxonomic remark: A medium-sized species of Ptychadena. It differs in morphological characters, advertisement call, and 16S rRNA sequence from the other Ptychadena species from Rwanda. The taxonomic status of the species will be addressed elsewhere (Dehling, in preparation).
Distribution in Rwanda: So far, only known from the Eastern Province.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Population trend and local abundance unknown. Proposed national conservation status: Data Deficient.
3.1.55. Ptychadena sp. B
Taxonomic remark: A small species of Ptychadena. It differs in morphological characters, advertisement call, and 16S rRNA sequence from the other Ptychadena species from Rwanda. The taxonomic status of the species will be addressed elsewhere (Dehling, in preparation).
Distribution in Rwanda: So far, only known from the Eastern Province.
Conservation status and population trend: IUCN Red List: Not evaluated [
7]. Population trend and local abundance unknown. Proposed national conservation status: Data Deficient.
Three species of the genus Amietia and one of the genus Cacosternum have been recorded in Rwanda. Morphological distinction of Amietia spp. is difficult, whereas all taxa are well differentiated with respect to partial 16S sequences. The advertisement call of A. ruwenzorica is unknown. The calls of the remaining species allow for a reliable distinction of reproductive males.
Key to the species of Amietia of Rwanda based on morphological characters:
- 1
Webbing well developed, leaving at most one phalanx free on the first and fifth toe, conspicuous narrow longitudinal dorsolateral dermal ridges from eye to sacrum, dorsal pattern consisting of blotches .................................................... 2
- 1′
Webbing poorly developed, leaving two phalanges free on first and fifth toe, low, rather inconspicuous, dorsolateral fold from eye to about half the trunk length, narrow longitudinal dark marks along the dorsolateral folds .... A. ruwenzorica
- 2
Rictal gland flat with few protruding warts, transverse C-shaped dermal scapular fold usually present .................. A. desaegeri
- 2′
Rictal gland protruding and smooth, no dermal scapular fold .... A. nutti
Key to the pyxicephalid species of Rwanda based on advertisement call features:
- 1
Call consisting of a series of evenly spaced single pulses followed by a long, pulsed note ...2
- 1′
Call consisting of 5–6 notes consisting of 2 to 4 pulses ........................................................................ Cacosternum plimptoni
- 2
Number of single pulses in series up to 5, repeated at 8–11/s ............................................................................... Amietia desaegeri
- 2′
Number of single pulses in series up to 10, repeated at 12–20/s ...................................................................................... Amietia nutti
Figure 16.
Rwandan species of Pyxicephalidae (A) Amietia desaegeri, Gishwati-Mukura NP (photo: J.M.D.), (B) Amietia nutti, Huye (photo: J.M.D.), (C) Amietia ruwenzorica, Volcanoes NP (photo: J.M.D.); Ranidae (D) Amnirana albolabris, South Kivu Province, DR Congo (photo: Bonny Dumbo), (E) Amnirana galamensis, Eastern Province (photo: Eberhard Fischer); and Herpelidae (F) Boulengerula fischeri, Cyamudongo Forest (photo: J.M.D.).
Figure 16.
Rwandan species of Pyxicephalidae (A) Amietia desaegeri, Gishwati-Mukura NP (photo: J.M.D.), (B) Amietia nutti, Huye (photo: J.M.D.), (C) Amietia ruwenzorica, Volcanoes NP (photo: J.M.D.); Ranidae (D) Amnirana albolabris, South Kivu Province, DR Congo (photo: Bonny Dumbo), (E) Amnirana galamensis, Eastern Province (photo: Eberhard Fischer); and Herpelidae (F) Boulengerula fischeri, Cyamudongo Forest (photo: J.M.D.).
Figure 17.
Advertisement calls of Rwandan Pyxicephalidae (
A)
Amietia desaegeri, (
B)
Amietia nutti, and (
C)
Cacosternum plimptoni; and Ranidae (
D)
Amnirana galamensis (call recording from South Africa [
107]). Calls are shown as representative spectrograms and oscillograms.
Figure 17.
Advertisement calls of Rwandan Pyxicephalidae (
A)
Amietia desaegeri, (
B)
Amietia nutti, and (
C)
Cacosternum plimptoni; and Ranidae (
D)
Amnirana galamensis (call recording from South Africa [
107]). Calls are shown as representative spectrograms and oscillograms.
3.1.56. Amietia desaegeri (Laurent, 1972) (Figure 16A and Figure 17A)
Original name and description: Rana desaegeri—R. F. Laurent (1972). Exploration du Parc National des Virunga. Deuxième série, 22: 1–125 [p. 91].
Type locality: “riv. Byangolo, secteur Munsenene, 1.300 m” (=“Byangolo River, Munsenene Sector”), Virunga National Park, North Kivu Province, DR Congo.
External morphology: de Saeger’s River Frog is a large anuran species [
6,
73]. Sexual size dimorphism absent, SVL of males up to 78 mm, females up to 76.5 mm [
73]. Dorsum smooth with conspicuous narrow longitudinal dorsolateral dermal ridges from posterior end of eyelid to sacrum. Transverse, c-shaped ridge between dorsolateral ridges at level of scapula. Legs long, feet large, toes extensively webbed. Coloration in life: Dorsum pale beige with brown infusions but no distinct markings. Pale band between the eyes. Venter immaculate, with diffuse darker mottling on throat.
Advertisement call: The call consists of two parts. The first part is a series of (1-)3–5 evenly spaced, single pulses, emitted at at rate of 11/s, with a dominant frequency at 1300 Hz (
Figure 17A). The pulse series is followed by a long note after an interval of 628–1398 ms. The long note lasts 49 to 402 ms and consists of 6–50 pulses, repeated at a rate of 125 pulses/s. The amplitude and the dominant frequency (1000–1300 Hz) increase from the first to the last pulse of the note. Both parts are frequently emitted without the other, suggesting that both serve different functions. The call has also been described elsewhere [
73].
Distribution: The species is endemic to the Albertine Rift and is distributed in eastern DR Congo and in western Rwanda at elevations of 1000–3500 m. Altitudinal range and province records in Rwanda: 1850–2082 m; Western Province.
Habitat preference and ecology: Terrestrial. Inhabits pristine submontane and montane forest, usually associated with streams. The species breeds in small streams.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Vulnerable, because the species has lost most of its former habitat in Gishwati Forest within the last 30 years. The remaining restricted habitat is threatened by degradation.
3.1.57. Amietia nutti (Boulenger, 1896) (Figure 16B and Figure 17B)
Original name and description: Rana Nutti—G. A. Boulenger (1896). The Annals and Magazine of Natural History including Zoology, Botany, and Geology, Sixth Series, 18: 467–468 [p. 467].
Type locality: “Lake Tanganyika”.
External morphology: Nutt’s River Frog is a large anuran species. Pronounced sexual size dimorphism, SVL of males up to 70.1 mm, SVL of females up to 87.8 mm [
6,
73]. Dorsum smooth with narrow longitudinal dorsolateral dermal ridges from posterior end of eyelid to sacrum, paralleled by one to four less distinct and interrupted lateral dermal ridges or rows of tubercles. Transverse dorsal ridge absent. Legs long, feet large, toes extensively webbed. Coloration in life: Dorsum pale brown with small darker brown blotches, yellowish vertebral stripe sometimes present. Venter immaculate, with diffuse darker markings on throat and breast.
Advertisement call: The call consists of two parts. The first part is a series of 7–12 (rarely as few as 3) evenly spaced, single pulses, emitted at at rate of 15/s (12–20), with a dominant frequency at 1120 Hz. The amplitude increases markedly from the first to the last pulse of the series (
Figure 17B). The pulse series is followed by a long note after a long interval of 869–2273 ms. The long note lasts 126 to 231 ms and consists of an initial series of up to 20 pulses that are repeated at a continuously increasing rate from 62/s between the first two pulses to about 200/s between the last two separable pulses, before the note becomes pulsatile at about half its duration and finally tonal in the last fourth (
Figure 17B). The amplitude remains at the same level. The dominant frequency decreases during the pulsed part of the note from 1000 Hz to 560 Hz remains at about 600 Hz until the end of the note. During the pulsatile section of the note, seven harmonics between 860 Hz and 2400 Hz are expressed, and in the final tonal section, three harmonics at 1100 Hz, 1800 Hz, and 2400 Hz are present and contain almost as much energy as the fundamental/dominant frequency, which makes this final part sound higher than the rest of the note. The long note is occasionally repeated once or twice without a preceding pulse series after intervals of about 3.6 and 5.0 s, respectively. For additional call descriptions, see Refs. [
73,
76].
Distribution: The species is distributed from Ethiopia to Burundi, eastern DR Congo, and Tanzania at elevations of 200–2879 m. Altitudinal range and province records in Rwanda: 1292–2879 m; all five provinces.
Habitat preference and ecology: Terrestrial. Inhabits pristine forests, grassland, cultivated wetlands. Tolerant to human disturbance. The species breeds in slow-moving streams, ditches, agricultural water channels, and ponds.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend stable. Proposed category for the National Red List: Least Concern, because the species occurs commonly in protectes areas (Volcanoes and Nyungwe National Parks) as well as in cultivated wetlands throughout the country.
3.1.58. Amietia ruwenzorica (Laurent, 1972) (Figure 16C)
Original name and description: Rana ruwenzorica—R. F. Laurent (1972). Exploration du Parc National des Virunga. Deuxième série, 22: 1–125 [p. 93].
Type locality: “Kikyo, secteur Munsenene, forêt de montagne [=Munsenene Sector, montane forest], 2080 m”, Virunga National Park, North Kivu Province, DR Congo.
External morphology: The Ruwenzori River Frog is a large anuran species. Pronounced sexual size dimorphism, SVL of males 44–71 mm, SVL of females 46–86 mm [
38]. Dorsum smooth with an inconspicuous low longitudinal dorsolateral fold from posterior end of the eyelid to about half the trunk length. Transverse dorsal ridge absent. Legs long, feet very large, toe webbing poorly developed. Coloration in life: Dorsum cream to light brown with irregular larger blotches on flanks and along dorsolateral fold. Sides of head behind eyes dark brown and with a large dark blotch anteroventrally from eye. Gular region immaculate to boldly marbled. Marbling may extend to the lower belly and to the ventral side of thighs.
Advertisement call: The call of the species is unknown.
Distribution: The species is endemic to the Albertine Rift and occurs in the Ruwenzori Mountains, Bwindi Impenetrable Forest in Uganda, Virunga Mountains of Rwanda and the Itombwe Plateau and Kabobo Mountains in DR Congo at elevations of 700–3031 m. Recorded from Volcanoes National Park in Rwanda. Altitudinal range and province records in Rwanda: 3031 m; Northern Province.
Habitat preference and ecology: Terrestrial. Inhabits swamps and open areas within montane forest, associated with small streams, which are used for breeding.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend decreasing. Proposed category for the National Red List: Data Deficient.
3.1.59. Cacosternum plimptoni Channing, Brun, Burger, Febvre & Moyer, 2005 (Figure 14H and Figure 17C)
Original name and description: Cacosternum plimptoni—A. Channing, C. Brun, M. Burger, S. Febvre & D. Moyer (2005). African Journal of Herpetology, 54: 139–148 [p. 140].
Type locality: “Musabi Plain (02°17′16″ S; 34°28′16″ E; 1297 m), in the Serengeti National Park, Tanzania”.
External morphology: Plimpton’s Dainty Frog is a very small anuran species. Sexual size little developed, SVL of males 16.4–19.2 mm, SVL of females up to 19.7 mm [
4,
6]. Head relatively narrow, body widest at level of abdomen. Legs short, feet relatively large, toe webbing absent. Coloration in life: Dorsum green to brown with a variable pattern of spots or stripes. Venter with few dark spots. Reproductive males have a yellow throat.
Advertisement call: The call is a rapid series of five or six high-pitched click notes. Each note consists of 2–4 pulses. Note repetition rate 12–13/s. Total call duration 348 (five notes) to 438 ms (six notes) (
Figure 17C). Intercall interval varies between 2.1 and 7.9 s (depending on temperature or social interactions with conspecifics). Dominant frequency is at 4600 Hz. The call of specimens from Tanzania has been described in detail by Ref. [
157].
Distribution: The species has been recorded in Tanzania, Kenya, Uganda, DR Congo, and Rwanda at elevations of 1200–2500 m. Altitudinal range and province records in Rwanda: A single locality at 1290 m in Akagera National Park; Eastern Province.
Habitat preference and ecology: Terrestrial. Inhabits grassland and dry savanna in highland areas. The species breeds in shallow ponds, flooded depressions, and puddles. Males call hidden in vegetation from the edge of the water body. Reproduction is in the rainy season from October to April.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient.
Two Amnirana species occur in the eastern part of Rwanda. Both are known from only a few localities in the Eastern Province.
Key to the species of Amnirana of Rwanda based on morphological characters:
- 1
Dorsolateral fold narrow, well defined, dorsal colour greenish brown, fingertips distinctly expanded .... A. albolabris
- 1′
Dorsolateral fold thick and glandular, dorsal colour dark brown and yellow, fingertips slightly swollen .... A. galamensis
3.1.60. Amnirana albolabris (Hallowell, 1856) (Figure 16D)
Original name and description: Rana albolabris—E. Hallowell (1856). Proceedings of the Academy of Natural Sciences of Philadelphia, 8: 146–153 [p. 153].
Type locality: “West Africa”; restricted to “Gabon” by Ref. [
158], further restricted to “north of the Ogooué River in Gabon” by Ref. [
159].
Taxonomic remark: This taxon is a complex of up to six species [
159]. The Rwandan population is not conspecific with the nominal taxon but belongs to a Central/East African lineage.
External morphology: Central and East African Forest White-lipped Frogs are medium-sized to large anuran species. Pronounced sexual size dimorphism, SVL of males up to 57 mm, SVL of females up to 74 mm [
4,
6]. Elongated body; snout moderately pointed. Legs lon, feet large, toes extensively webbed. Tympanum large. Longitudinal dorsolateral dermal folds from eye to sacrum. Tips of fingers and toes expanded to discs. Reproductive males with nuptial pads on thumb, elongated glands on the upper arm, minute spines on the back. External vocal sac absent. Coloration in life: Dorsum olive to dark brown with black markings. Venter whitish, occasionally with dark patterning.
Advertisement call: Unknown.
Distribution: The Central/East African lineage of
A. albolabris occurs in the DR Congo, Rwanda, Tanzania, Uganda, and Keyna at an altitudinal range of 600–1800 m [
38,
159,
160]. In Rwanda, the species is known only from the Akagera-River Basin [
48]. Altitudinal range and province records in Rwanda: 1290 m; Eastern Province.
Habitat preference and ecology: Terrestrial. Inhabits pristine and degraded forest, and gallery forest, associated with edge habitats to open areas. Tolerant to human disturbance. The species breeds in ponds, marshes, raphia swamps, and creeks.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient.
3.1.61. Amnirana galamensis (Duméril & Bibron, 1841) (Figure 16E and Figure 17D)
Original name and description: Rana Galamensis—A.-M.-C. Duméril & G. Bibron (1841). Erpétologie générale ou histoire naturelle complète des reptiles. Tome huitième, 784 pp. [p. 367].
Type locality: “Sénégal [...] dans les étangs de Galam” (= “in the ponds of Galam”, “Senegal”).
Taxonomic remark: This taxon is a complex of up to three species [
159]. The Rwandan population is not conspecific with the nominal taxon but belongs to a Central or East African lineage (possible available name:
Rana oubanghiensis Mocquard, 1896). Referred to as
Amnirana sp. ‘galamensis East’ in Ref. [
6].
External morphology: The East African Galam White-lipped Frog is a large anuran species. Sexual size dimorphism weekly expressed, SVL of males up to 78 mm, females up to 86 mm [
4,
6]. Body robust; snout rounded. Legs relatively short, toes moderately webbed. Toe and fingertips not expanded. Tympanum large. Thick, conspicuous dorsolateral dermal ridge extending on both sides from eye to sacrum. Numerous small, gland-like warts on flanks. Males have a pair of vocal sacs on in front of arm insertion and large glands on upper arm. Coloration in life: Dorsum dark brown with several paler yellow spots in the anal region and on the thighs. A pale yellow or orange stripe runs from the nostril across the eyelid along the dorsolateral ridge to the body end. Upper lip white. Venter whitish to light gray, lateral regions of belly and hind limbs mottled with dark.
Advertisement call: The pulsed call is low grunting [
6,
107]. The calls have a duration of 365–537 ms and 59–91 pulses. The power spectrum of the calls consists of three frequency bands (ca. 500, 2500, and 5000 Hz) with almost the same intensity. A spectrogram of the call has been depicted elsewhere [
107].
Distribution: The Central and East African lineages of
A. galamensis are distributed from Ethiopia and South Sudan through the Albertine Rift to Zambia and Mozambique, and along the African East Coast to Kenya and Somalia [
6,
159] at elevations from sea level to at least 1500 m. In Rwanda, the species is known from Lake Muhazi [
11] and the Akagera National Park and its vicinity [
48]. Altitudinal range and province records in Rwanda: 1232–1500; Eastern Province.
Habitat preference and ecology: Semi-aquatic, savanna, grasslands, where it lives in and around permanent lakes, small rivers, ponds, and swamps. Tolerant to human disturbance. Breeding habitat: lakes, ponds, and swamps.
Conservation status and population trend: IUCN Red List: Least Concern [
7]. Population trend unknown. Proposed category for the National Red List: Data Deficient.
A single caecilian species occurs in Rwanda. Limbless, its body very elongated and longitudinally segmented through body rings, and its eyes reduced, it cannot be confused with any other amphibian species in the country.
3.1.62. Boulengerula fischeri Nussbaum & Hinkel, 1994 (Figure 16F)
Original name and description: Boulengerula fischeri—R. A. Nussbaum & H. Hinkel (1994). Copeia 1994: 750–760 [p. 756].
Type locality: “Forêt de Cyamudongo near Forêt de Nyungwe, 1700–2000 m, Cyangugu Prefecture, Rwanda”.
External morphology: Fischer’s Caecilian is an extremely elongated caecilian with 183–204 body rings and a width at midbody of 3.0–4.5 mm [
6,
51,
52]. Sexual size dimorphism absent, body length of males 317–386 mm, body length of females 257–397 mm, [
52]. Coloration in life: Surface pink to light violet, head and neck and posterior end slightly paler. At the midbody a darker mid-dorsal stripe.
Distribution: The species is a local endemic of Rwanda, known exclusively from the Cyamudongo Forest (Western Province) and its immediate vicinity at elevations of 1743–1985 m [
52,
60]. Nothing is known about the reproduction.
Habitat preference and ecology: Fossorial. Inhabits pristine montane forest and adjacent banana plantations.
Conservation status and population trend: IUCN Red List: Vulnerable [
7]. Population trend unknown. Proposed category for the National Red List: Vulnerable.
3.2. Species without Validated Country Record That Have Been Presumed to Occur in Rwanda
Besides the species, for which there are validated country records (
Section 3.1.), several other species have been listed for Rwanda in regional or continental field guides [
3,
5,
6] and in the online data bases “Amphibian species of the world” [
8], “IUCN Red List” [
7], and AmphibiaWeb [
95]. These references might have had a reciprocal influence to list certain species.
Some species are listed for Rwanda because they have been recorded from localities in neighboring countries close to the border with Rwanda. Their presence in Rwanda is therefore only presumed or expected [
8] or accordingly stated as “uncertain” [
7] or extrapolated on a distribution map to cover all East Africa [
3]. There are no published reports or voucher specimens in museum collections from Rwanda, nor did our surveys yield any evidence that the geographic distribution of these species reaches Rwanda. The following species fall into this category:
Arthroleptis lameerei de Witte, 1921 [
8],
Arthroleptis pyrrhoscelis Laurent, 1952 [
7,
8],
Sclerophrys channingi (Barej, Schmitz, Menegon, Hillers, Hinkel, Böhme & Rödel, 2011) [
7,
8],
Hemisus guineensis Cope, 1865 [
7,
8],
Hemisus marmoratus (Peters, 1854) [
3,
7,
8],
Hyperolius chrysogaster Laurent, 1950 [
7],
Hyperolius diaphanus Laurent, 1972 [
7,
8],
Hyperolius kuligae Mertens, 1940 [
7],
Hyperolius langi Noble, 1924 [
7,
8],
Hyperolius marginatus Peters, 1854 [
7,
8],
Hyperolius ocellatus Günther, 1858 [
7,
8],
Hildebrandtia ornata (Peters, 1878) [
2].
In other cases, the taxonomy of species that occur in Rwanda had been changed because the Rwandan populations were assigned to different taxa or described as new species. Apart from the cases we report herein, i.e.,
Sclerophrys berghei, formerly
S. funerea (3.1.10), and
S. gutturalis, formerly
S. regularis (3.1.11) [
7,
8,
95], there are several instances, in which the old name alongside the new name is still listed. These cases include the following species:
Afrixalus fulvovittatus (Cope, 1861),
Afrixalus “quadrivittatus” [
124]: The term
Afrixalus “
quadrivittatus” listed in “Amphibian species of the world” is applied to a conglomeration of species not unequivocally assignable to either
Afrixalus fulvovittatus or
Afrixalus vittiger [
8,
124]. Ref. [
95] lists
Afrixalus fulvovittatus for Rwanda. However, only a single species of the
Afrixalus fulvovittatus/
A. quadrivittatus complex occurs throughout Rwanda and we agree with other published reports that the East African Four-lined Spiny Reed Frog populations, including those in Rwanda, pertain to the nominal species
Afrixalus quadrivittatus (3.1.16) [
6,
76,
109,
123,
161].
Afrixalus laevis (Ahl, 1930): The species is still listed for Rwanda [
7,
8,
95]. The populations from the Central African Albertine Rift that formerly had been assigned to
Afrixalus laevis were described as distinct species,
Afrixalus phantasma (3.1.15) and
A. lacustris Greenbaum, Dehling, Kusamba & Portik, 2022, and the geographical range of
A. laevis was restricted to Cameroon and Bioko Island, Equatorial Guinea [
86]. Only one of these species,
A. phantasma, has been recorded from Rwanda.
Hyperolius acuticeps Ahl, 1924: The East African species of the
Hyperolius-
nasutus group had been referred to as
H. acuticeps [
4,
126,
160,
162,
163,
164,
165], before the group was revised and the only species of the group that occurs in Rwanda was described as a new species,
H. rwandae (3.1.28) [
82].
Hyperolius acuticeps is still listed for Rwanda [
82,
95], although the taxon is now considered a synonym of
H. microps Günther, 1864 [
166], a species that is distributed in Mozambique, Malawi, and coastal Kenya and Tanzania [
6,
8].
Ptychadena “mascareniensis” [
8]: This taxon, listed in Ref. [
8], is regarded an aggregation of unnamed species and unallocated populations within the
Ptychadena mascareniensis complex, distributed from West Africa to northeastern DR Congo, Ethiopia, northern Namibia, and northern South Africa [
8,
71,
88,
167]. The species in Rwanda formerly treated as
P. mascareniensis has been identified as
P. nilotica (3.1.50) [
88]. Listing “
P. mascareniensis” alongside
P. nilotica in the species list for Rwanda implies that another unnamed species of the complex occurs in Rwanda, which according to our investigations is not the case.
Lastly, there are several species that have been claimed to occur in Rwanda but there are no published reports or voucher specimens in museum collections that would support the assumed occurrence in Rwanda.
Leptopelis christyi (Boulenger, 1912); type locality: “Mabira Forest, Chagwe, Uganda”. The species is regarded as occurring in Rwanda in regional and continental field guides [
5,
6] and is presumed to occur [
8] or considered “uncertain” [
7] in two data bases. The large species of
Leptopelis is known to occur in DR Congo, Tanzania, and Uganda [
123,
168]. The extension of the geographic range to Burundi and Rwanda [
5,
6] is not supported by published reports from these countries. Our surveys did not yield any evidence that its geographic distribution covers Rwanda.
Sclerophrys pusilla (Mertens, 1937); type locality: “Leteba-Camp”, Kruger National Park, Limpopo Province, South Africa. The species is regarded as occurring in Rwanda in a regional [
2] (as
Bufo maculatus) and a continental field guide [
6] as well as in a database [
95]; and presumed to occur [
8] or consistered “uncertain” [
7] in the two other data bases. The taxon is widely distributed in southern and eastern Africa [
169], but there is no published report on specimens collected in Rwanda. Our surveys did not yield any evidence that its geographic distribution reaches Rwanda.
Afrixalus osorioi (Ferreira, 1906); type locality: “Quilombo”, Angola. The species is stated as occurring in Rwanda in a regional field guide [
5] and its presence is considered possible in a data base [
8]. The species occurs from Gabon to northern Angola, through DR Congo to Uganda and Kenya [
6,
38,
44,
163]. There is no published report on specimens collected in Rwanda. Our surveys did not yield any evidence that its geographic distribution reaches Rwanda.
Hyperolius mariae Barbour & Loveridge, 1928; type locality: “near the mill dam at Derema, Usambara Mtns., Tanganyika Territory” (=Tanzania). Three species of the
Hyperolius viridiflavus/
H. marmoratus complex occur in Rwanda, i.e.,
H. glandicolor (3.1.23),
H. parallelus (3.1.27), and
H. viridiflavus (3.1.29) [
85]. A recent revision of the complex included our sequences from Rwanda and confirmed the presence, number, distinctness, and affiliation of the three species [
131]. In contrast to that,
Hyperolius glandicolor is missing from the list for Rwanda and
H. mariae is listed in its stead by Ref. [
8]. The reason is obviously a mix-up in the captions of Figures 8 and 9 in Ref. [
131]: Figure 8B shows the distribution of
H. glandicolor from northern Rwanda and southwestern Uganda to southern Kenya and northern Tanzania but is labelled as
H. mariae, and, vice versa, Figure 9C shows the distribution of
H. mariae in coastal Kenya and Tanzania including the islands off the coast but is labelled as
H. glandicolor. The correct localities are stated for each species in the respective accounts [
131].
Ptychadena grandisonae Laurent, 1954; type locality: “Muita, Luembe E., Angola” (=Muíta, 7.48° S, 21.27° E, 800 m [
35]; Lunda Norte Province, Angola). Two of the paratypes of the species were collected in “Bitare, 1650 m., Terr. de Kitega, Urundi” [
33], a town in central Burundi (3°15′ S, 29°54′ E), about 20 km north of Gitega [
89]. The species has been erroneously considered to occur in Rwanda, but not in Burundi, for almost 60 years [
89] and is still listed for Rwanda by all surveyed data bases (see Refs. [
7,
8,
95] and Ref. [
6]). There is no specimen of
P. grandisonae among the material Laurent collected in Rwanda. As we already pointed out [
89,
90], the misconception was very likely caused by Laurent himself in a monograph on the “Reptiles et Amphibiens de l’Angola” [
35]. Therein, Laurent cites one of the type localities of
P. grandisonae wrongly as “Bitare (Ruanda)” [
35] (p. 139), apparently confusing Bitare, Burundi, with Butare, Rwanda. Thus, the often-cited record of
P. grandisonae from Rwanda [
6,
147,
156,
170,
171] in fact refers to specimens from Burundi (RMCA 109036–37). So far, there is no evidence for the occurrence of
P. grandisonae in Rwanda.

























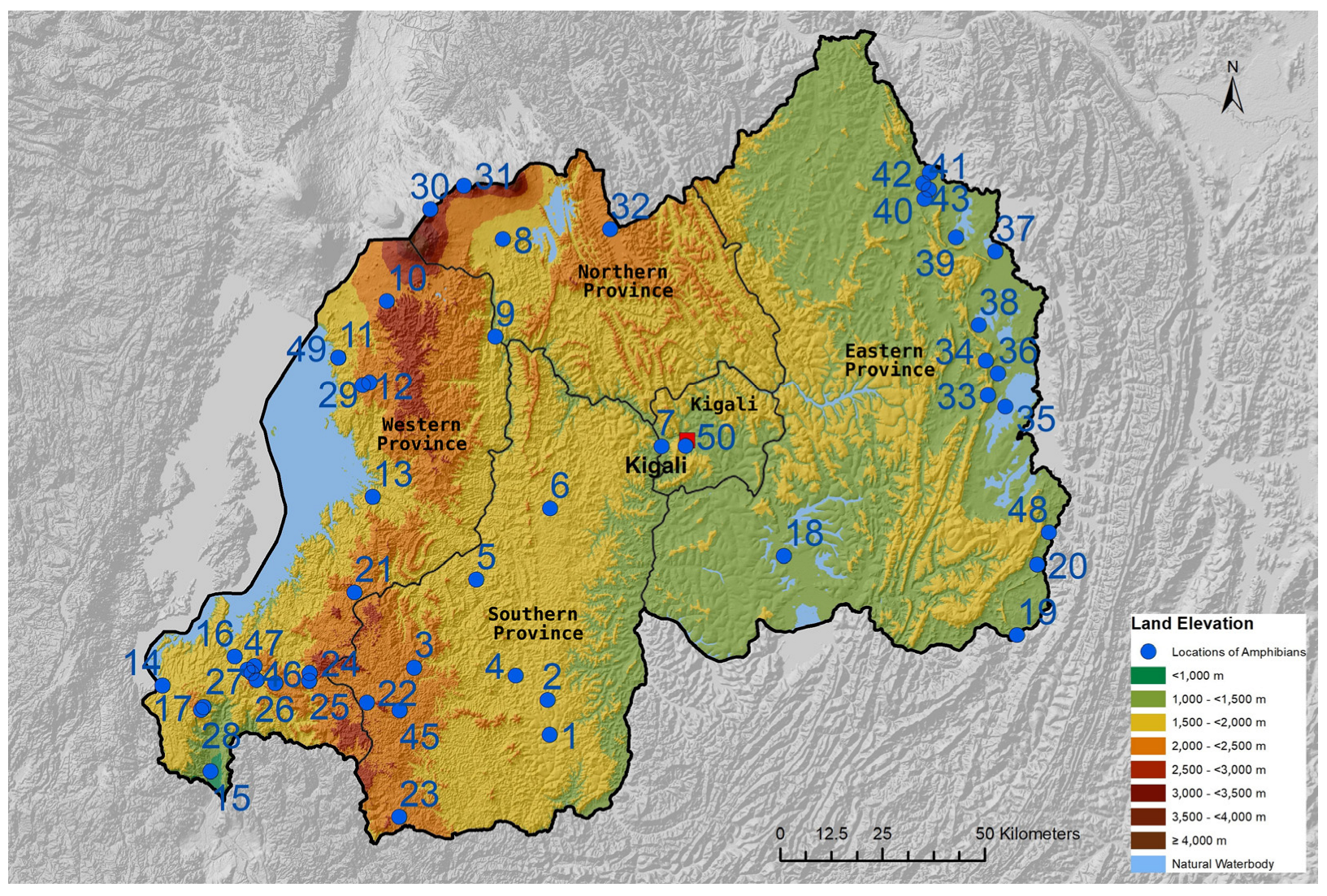{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}