
Description of a New Species of Parathalestris Brady & Robertson, 1873 (Copepoda: Harpacticoida: Thalestridae) from China, with a Key to Its Affiliated Species Group †


Abstract
:1. Introduction
2. Material and Methods
- Length-width ratio of female caudal ramus: (0) ≤ 1; (1) > 1.
- Bulbous seta on caudal ramus: (0) absent; (1) present.
- Armature formula of A2 exopod: (0) (1:1, 3); (1) (2:1, 2); (2) (2:1, 3).
- Inner edge of maxilliped basis: (0) straight; (1) concave.
- Length-width ratio of P1 endopod: (0) ≤ 5; (1) > 5.
- Rows of transversal spinules on female P5 baseoendopod: (0) absent; (1) present.
- Length of female P5: (0) extends to half-length but not reaching distal margin of the genital double-somite; (1) reaching distal margin of genital double-somite; (2) exceeding distal margin of genital double-somite, close to penultimate somite.
- Length of female P5 baseoendopod: (0) not exceeding distal margin of exopod; (1) exceeding distal margin of exopod.
- Shape of inner spine on male P1 basis: (0) normal; (1) curved.
- Armature formula of male P5: (0) 3/5; (1) 3/6; (2) 2/5.
- Median seta on male P5: (0) present, weak; (1) present, normal; (2) absent.
- Length of outermost seta on male P5 baseoendopod: (0) equal to median one; (1) half-length of median one; (2) less than half-length of median one.
3. Results
3.1. Systematics
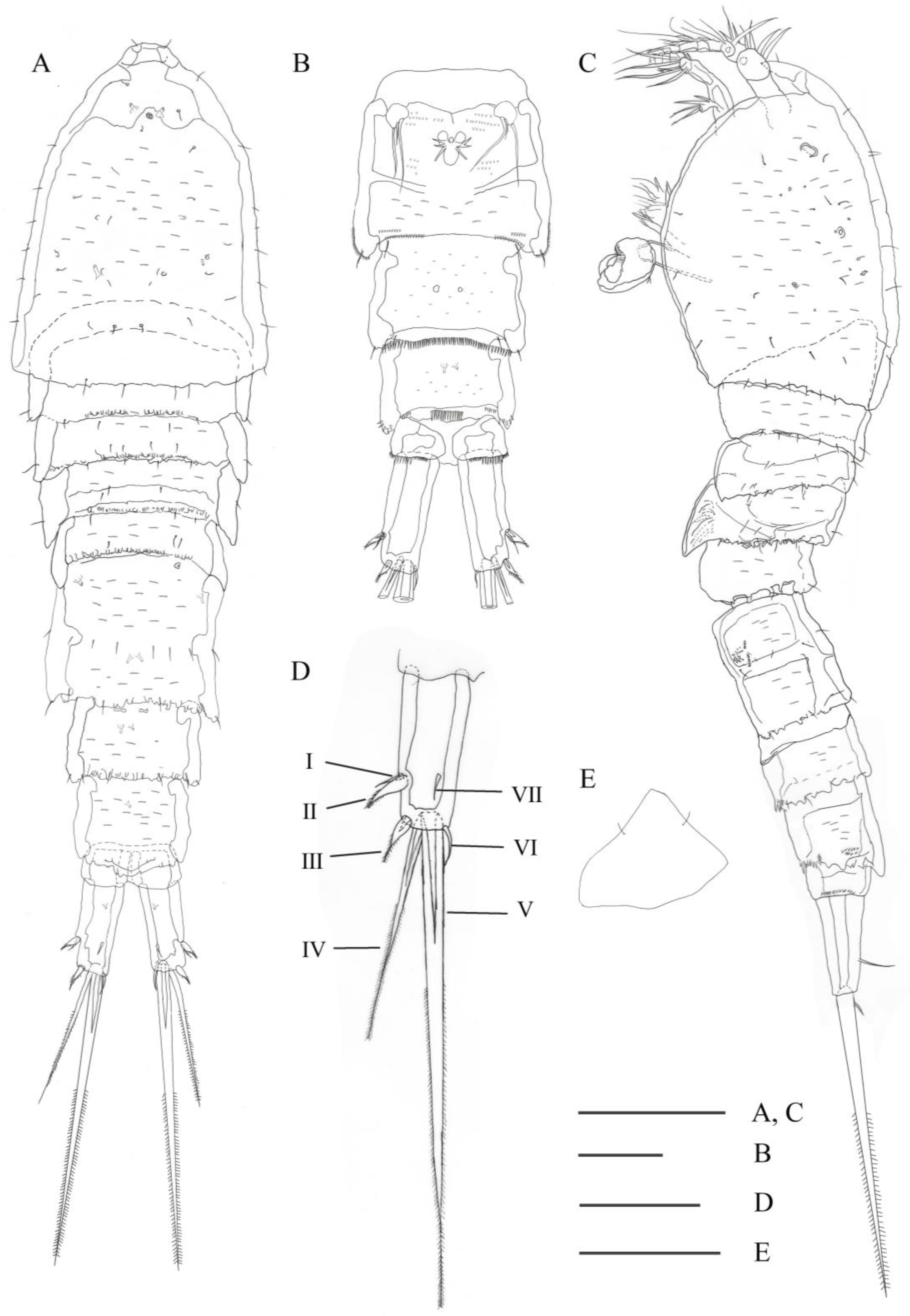
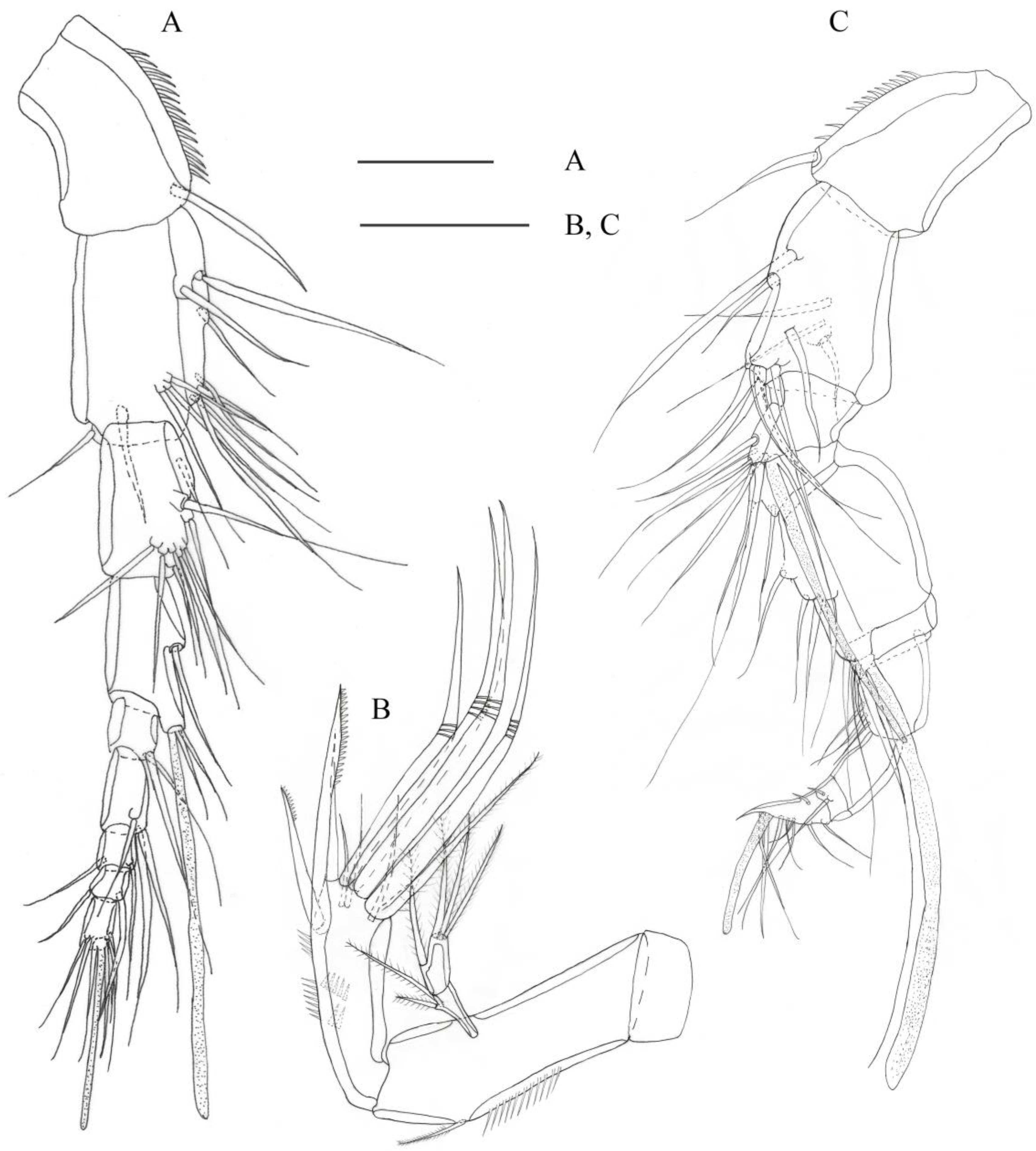
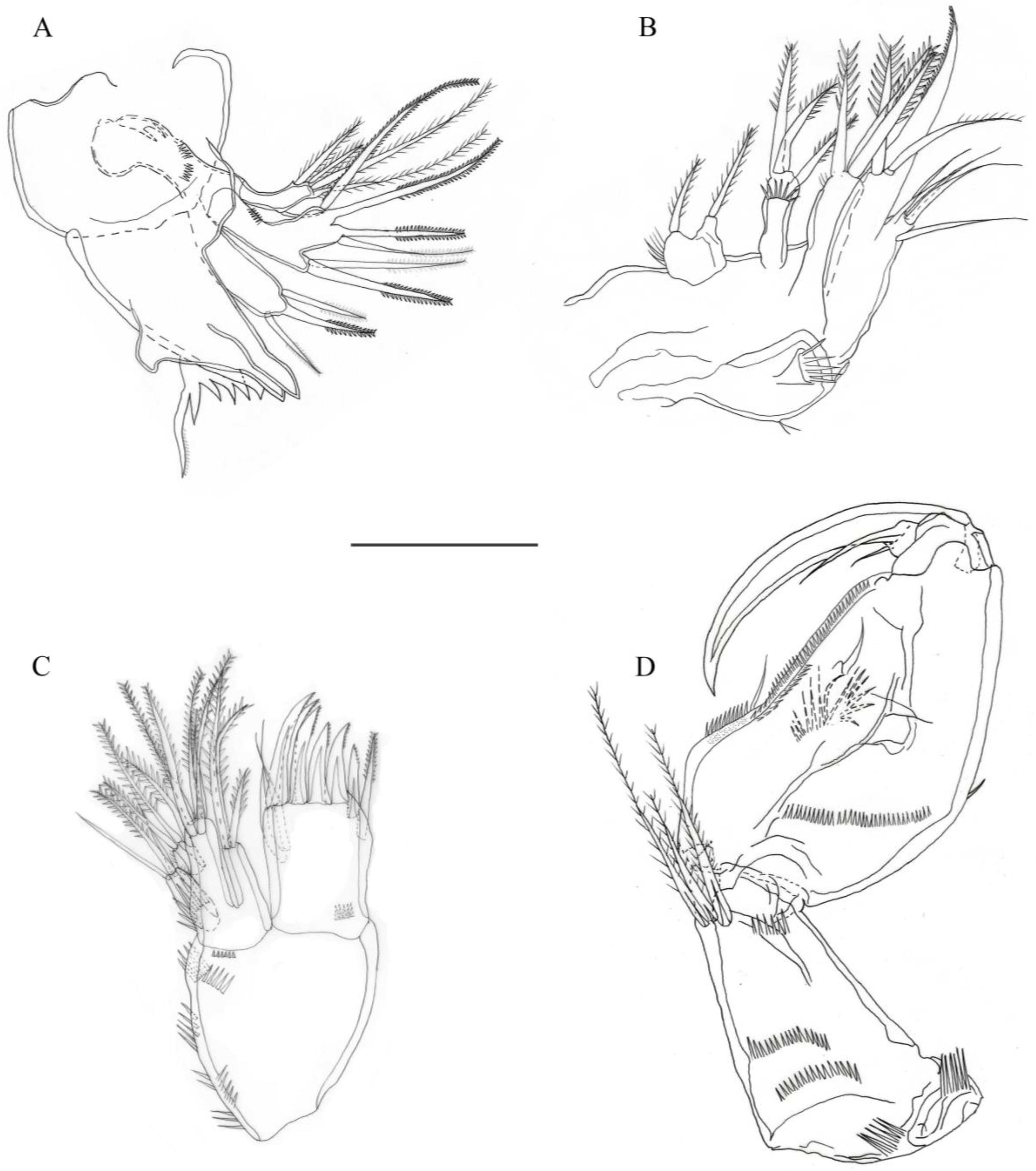
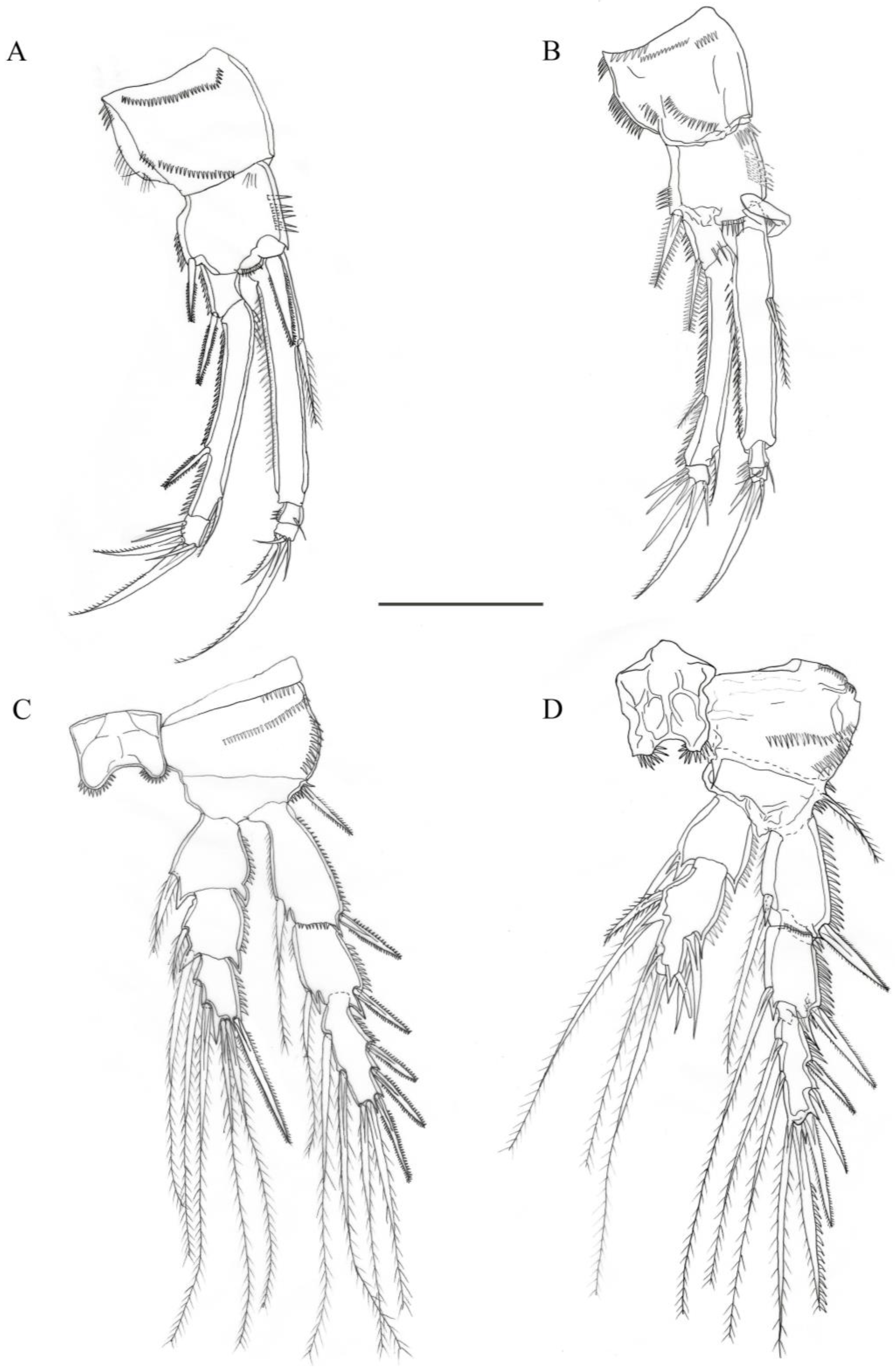
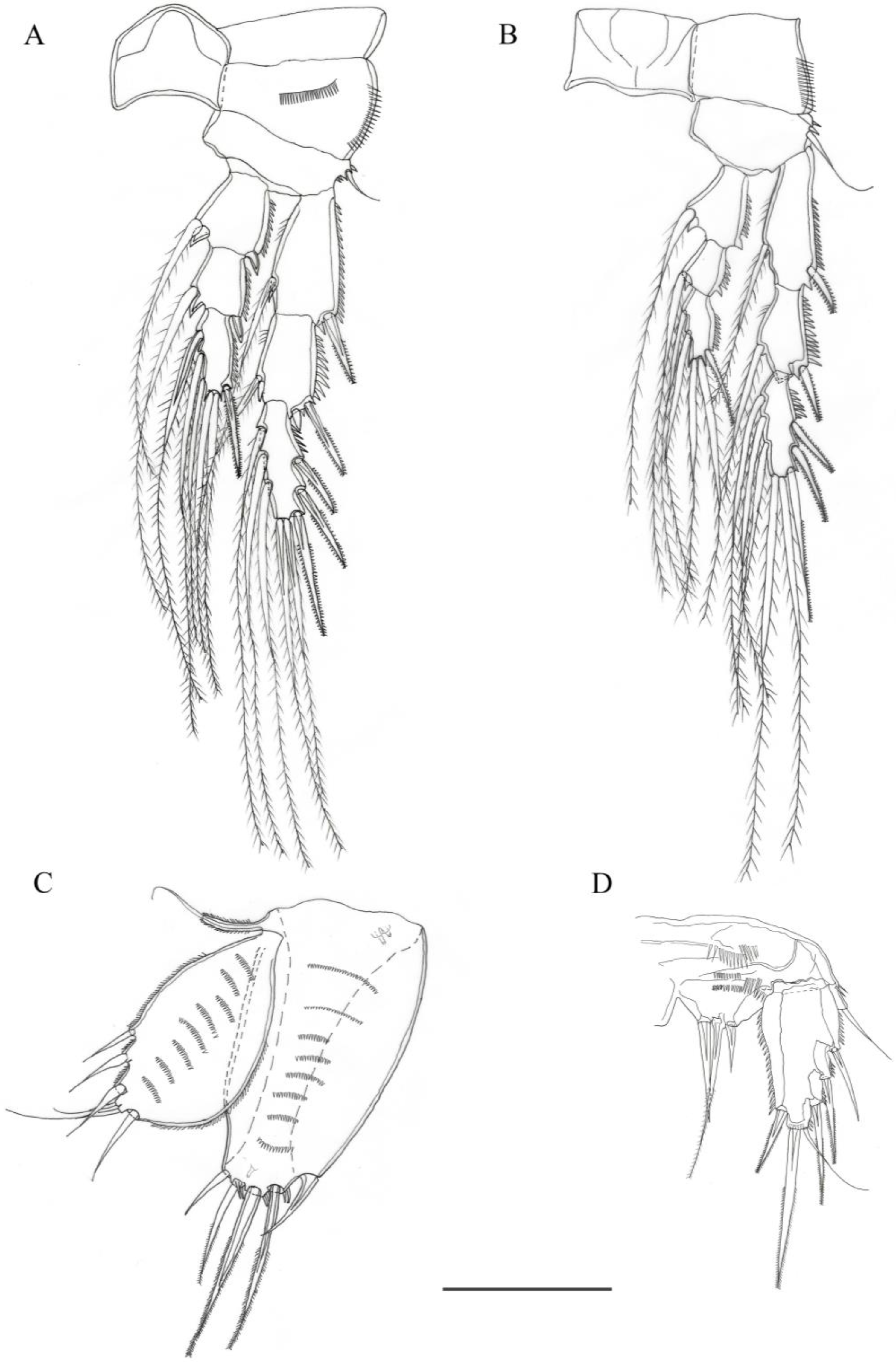
3.2. Description
3.2.1. Description of Female
3.2.2. Description of Male
3.2.3. Molecular Results
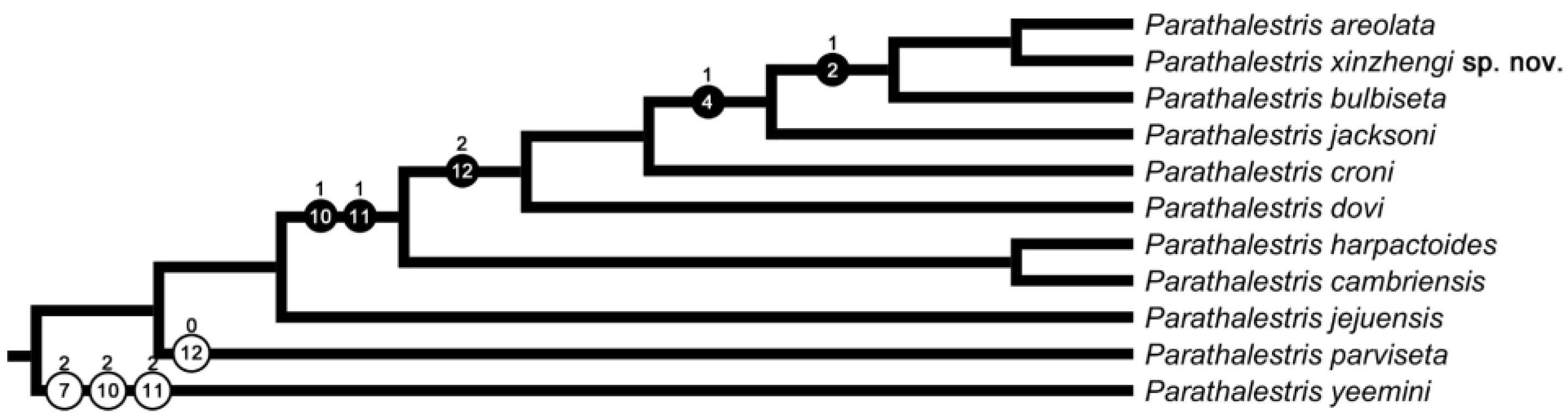
3.2.4. Morphological Analysis Results
4. Discussion
- Caudal ramus 4 times as long as wide …………… P. croni (Krøyer in Gaimard, 1842)Caudal ramus less than 4 times as long as wide …………………………………… 2
- Caudal ramus 2 times as long as wide ………………………….…………………… 3Caudal ramus as long as width or shorter than wide ………………….…………… 5
- Caudal ramus with bulbous seta ………………………………………………………… 4Caudal ramus without bulbous seta ………………………… P. jacksoni (Scott T., 1899)
- P1 enp-1 3.5 times as long as wide, female P5 exopod reaching beyond distal margin of baseoendopod ………………………………………… P. areolata Itô, 1972P1 enp-1 6.7 times as long as wide, female P5 exopod reaching close to distal margin of baseoendopod ………………………………………….……… P. xinzhengi sp. nov.
- Inner edge of basis of maxilliped concave, with an obtuse angle close to proximal margin, caudal ramus with bulbous seta …………………… P. bulbiseta Lang, 1965Inner edge of basis of maxilliped straight or convex, caudal ramus without bulbous seta ……………………………………………………………………………………… 6
- P1 enp-1 more than 5 times as long as wide ……………….………………………… 7P1 enp-1 less than or about 5 times as long as wide …………………………………… 9
- P1 enp-1 about 5 times as long as wide, female P5 beyond halfway but not beyond the end of the genital double-somite …………………………….………… 8P1 enp-1 about 6 times as long as wide, female P5 extends only to about halfway along the genital double-somite …………………………………… P. dovi Marcus, 1966
- Male P5 exopod rectangular, 2 times as long as wide; A2 exopod two-segmented, with setation formulae 2:1,2 ………….………………………… P. cambriensis Wells, 1964Male P5 exopod pyriform, 1.5 times as long as wide; A2 exopod two-segmented, with setation formulae 1:1,3 ………………………………… P. harpactoides (Claus, 1863)
- Exopod of male P5 baseoendopod with five setae ……………………………………… 10Exopod of male P5 baseoendopod with six setae ……………………………………… 11
- Inner margin of male P1 basis with curved spine; the middle seta of the male P5 baseoendopod minute ……………………………………… P. parviseta Chang & Song, 1997Inner margin of male P1 basis without spine; the innermost seta of the male P5 baseoendopod minute ……………………………… P. jejuensis Song & Hwang, 2010
- P1 enp-1 about 4 times as long as wide ……….……………………………………… 12P1 enp-1 about 5 times as long as wide ……………………… P. irelandica Roe, 1958
- Distal edge of male P2 Enp-2 with long, mucroniform projection and one stout, mucroniform spine on inner edge …………………………………… P. vinosa Pallares, 1975Distal edge of male P2 Enp-2 without project and one filiform seta on inner edge ……………………………………………………………………… P. similis Lang, 1936
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, C.Y.; Song, S.J. Marine harpacticoids of genus Parathalestris (Copepoda, Harpacticoida, Thalestridae) from Korea. Korean J. Syst. Zool. 1997, 13, 221–231. [Google Scholar]
- Song, S.J.; Hwang, U.W. Parathalestris jejuensis sp. nov. (Copepoda, Harpacticoida, Thalestridae) from macroalgal beds from Korea. Crustaceana 2010, 83, 1337–1348. [Google Scholar] [CrossRef]
- Wells, J.B.J. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef]
- Lang, K.A. Copepoda harpacticoidea from the Californian Pacific coast. K. Sven. Vetensk. Handl. 1965, 10, 1–566. [Google Scholar]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; The Ray Society: London, UK, 2004; p. 9. [Google Scholar]
- Ho, J.-S.; Hong, J.-S. Harpacticoid copepods (Thalestridae) infesting the cultivated Wakame (brown alga, Undaria pinnatifida) in Korea. J. Nat. Hist. 1988, 22, 1623–1637. [Google Scholar] [CrossRef]
- Huys, R. Harpacticoid copepods-their symbiotic associations and biogenic substrata: A review. Zootaxa 2016, 4174, 448–729. [Google Scholar] [CrossRef]
- Chullasorn, S.; Kangtia, P.; Song, S.J. A new species of the genus Parathalestris (Copepoda: Harpacticoida: Thalestridae) from dead staghorn corals on Tao Island, southern Thailand. Raffles Bull. Zool. 2016, 64, 343–350. [Google Scholar] [CrossRef]
- Itô, T. Descriptions and Records of Marine Harpacticoid Copepods from Hokkaido, III. Annot. Zool. Jpn. 1970, 43, 211–218. [Google Scholar]
- Itô, T. Descriptions and Records of Marine Harpacticoid Copepods from Hokkaido, IV (With 18 Text-figures and 2 Tables). J. Fac. Sci. Hokkaido Univ. Ser. 6 Zool. 1972, 18, 305–336. [Google Scholar]
- Chislenko, L.L. Copepoda Harpacticoida of the Karelia coast of the White Sea. Zool. Inst. 1967, 7, 48–196. [Google Scholar]
- Lian, G.; Sun, R.; Wang, Y.; Huang, J. Species Diversity of Marine Harpacticoid Copepods in China’s Seas and Adjacent Waters; Science Press: Beijing, China, 2022; pp. 1–206. [Google Scholar]
- Liu, Q.H. Study of the Meiofaunal Community Structure and Benthic Copepods in the Border of the Yellow Sea and East China Sea. Ph.D. Thesis, University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Science), Beijing, China, 2019. [Google Scholar]
- Wang, Y.; Zhang, M.; Shi, J.G.; Yu, W.W.; Shu, A.Y.; Sun, B.; Cheng, S.Q. Research progress of anti-fouling materials for fishing and its application in fisheries. Mar. Fish. 2021, 43, 247–256. [Google Scholar] [CrossRef]
- Lim, B.J.; Bang, H.W.; Moon, H.; Back, J. Integrative description of Diosaccus koreanus sp. nov. (Hexanauplia, Harpacticoida, Miraciidae) and integrative information on further Korean species. ZooKeys 2020, 927, 1–35. [Google Scholar] [CrossRef]
- Folmer, O.F.; Black, M.B.; Hoeh, W.R.; Lutz, R.V.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Huys, R.; Gee, J.M.; Moore, C.G.; Hamond, R. Marine and brackish water harpacticoid copepods. Part 1. In Synopses of the British Fauna (New Series); Kermack, D.M., Barnes, R.S.K., Crothers, J.H., Eds.; Linnean Society of London and the Estuarine and Coastal Sciences Association by Field Studies Council: London UK, 1996; Volume 51, pp. 1–352. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef]
- Ingólfsson, A. Vital role of drift algae in the life history of the pelagic harpacticoid Parathalestris croni in the northern North Atlantic. J. Plankton Res. 1997, 19, 15–27. [Google Scholar] [CrossRef]
- Lang, K. Monographie der Harpacticiden; Håkan Ohlsson: Lund, Sweden, 1948; Volume 2, pp. 1–1683. [Google Scholar]
- Brady, G.S. A Monograph of the Free and Semi-Parasitic Copepoda of the British Islands; Ray Society: London, UK, 1880; pp. 182+82 plates. [Google Scholar]
- Claus, C.F.W. Die Frei Lebenden Copepoden mit Besonderer Berücksichtigung der Fauna Deutschlands, der Nordsee und des Mittelmeeres; Wilhelm Engelmann: Leipzig, Germany, 1863; pp. 230+37 plates. [Google Scholar]
- Sars, G.O. An Account of the Crustacea of Norway, with Short Descriptions and Figures of all the Species. In Copepoda Harpacticoida, Parts IX & X.; Bergen Museum: Bergen, Norway, 1905; Volume V, pp. 1903–1911. [Google Scholar] [CrossRef]
- Scott, T. Report on the marine and freshwater crustacea from Franz-Josef Land, collected by Mr. William S. Bruce, of the Jackson-Harmsworth Expedition. Zool. J. Linn. Soc.-Lond. 1899, 27, 60–126. [Google Scholar] [CrossRef]
- Roe, K.M. The littoral harpacticids of the Dalkey (Co. Dublin) area with descriptions of six new species. Proc. R. Ir. Acad. B 1957, 59, 221–255. [Google Scholar]
- Wells, J.B.J. Some additions to the Dale Fort marine fauna. Proc. Zool. Soc. Lond. 1964, 142, 453–458. [Google Scholar] [CrossRef]
- Apostolov, A. Harpacticöides nouveaux de la Mer Noire et de la faune bulgare. Acta Zool. Bulg. 1977, 7, 8–21. [Google Scholar]
- Apostolov, A.; Marinov, T.M. Copepoda Harpacticoida (morski kharpaktikoidi). Izd Vo Na Bŭlgarskata Akad. A Na Nauk 1988, 18, 11–384. [Google Scholar]
- Bodin, P. Copépodes Harpacticoïdes Marins des Environs de la Rochelle. IV—Espèces de la Zone Intertidale des Nauteries. Tethys 1973, 4, 651–682. [Google Scholar]
- Chislenko, L.L. New common forms of harpacticids (Copepoda, Harpacticoida) from Possjet Bay of the Sea of Japan. Issled. Fauny Morei 1971, 8, 151–181. [Google Scholar]
- Gurney, R. Two new species of estuarine Copepoda. Zool. Anz. 1930, 87, 321–326. [Google Scholar]
- Moore, C.G. The harpacticoid families Thalestridae and Ameiridae (Crustacea, Copepoda) from the Isle of Man. J. Nat. Hist. 1976, 10, 29–56. [Google Scholar] [CrossRef]
- Scott, T. XXIV.—The Entomostraca of the Scottish National Antarctic Expedition, 1902–1904. Earth Environ. Sci. Trans. R. Soc. Edinb. 1912, 48, 521–599. [Google Scholar] [CrossRef]
- Masunari, S. Parathalestris mourei, a new species seaweeddwelling copepod (Harpacticoida, Thalestridae) from Santos Bay, Brazil. Crustaceana 1988, 54, 104–112. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. areolata | 1 | 1 | 2 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 2 |
| P. bulbiseta | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 |
| P. cambriensis | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 |
| P. croni | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | – | – |
| P. dovi | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 2 |
| P. harpactoides | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | – | 1 | 1 | 1 |
| P. jacksoni | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | – | – | – | – |
| P. jejuensis | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| P. parviseta | 0 | 0 | 2 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
| P. xinzhengi sp. nov. | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 |
| P. yeemini | 0 | 0 | 2 | 0 | 0 | 1 | 2 | 1 | 0 | 2 | 2 | – |
| Setal Formulae | Exopod | Endopod |
|---|---|---|
| P1 | 0.1.122 | 1.1.121 |
| P2 | 1.1.223 | 1.2.221 |
| P3 | 1.1.323 | 1.1.321 |
| P4 | 1.1.323 | 1.1.221 |
| Group | I | ||||||||
| Species | P. croni (Krøyer in Gaimard, 1842) | P. harpactoides (Claus, 1863) | P. jacksoni (Scott T., 1899) | P. similis Lang, 1936 | P. irelandica Roe, 1958 | P. cambriensis Wells, 1964 | P. bulbiseta Lang, 1965 | ||
| Length-width ratio of female caudal ramus | ≈4 | ≈1 | ≈2 | ≤1 | <1 | ≤1 | ≤1, with one bulbous seta distally | ||
| Armature formula of A2 exopod | 2(1:1,3) | 2(2:1,2) | 2(2:1,2) | unknown | 2(2:1,2) | 2(2:1,2) | 2(2:1,2) | ||
| Inner edge of maxilliped basis | straight | straight | concave | straight or convex | straight | straight | concave | ||
| Length-width ratio of P1 endopod | ≈4 | ≈5.5 | ≈4.5 | ≈4 | ≈5 | ≈5.5 | ≈4.5 | ||
| Length of female P5 baseoendopod | about 1/3 the length of exopod | not exceeding distal end of exopod | about half the length of exopod | not exceeding distal end of exopod | not exceeding distal end of exopod | slightly exceeding distal end of exopod | not exceeding distal end of exopod | ||
| Rows of transversal spinules on female P5 baseoendopod | absent | absent | absent | absent | absent | absent | absent | ||
| Length and armature formula of female P5 | extending about mid-length along genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | 5/6 | reaching about mid-length along genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | ||
| Inner spine of male P1 basis | curved inward | unknown | unknown | unknown | very short, curved inward | curved inward | curved outward | ||
| Armature formula of male P5 | 3/6 | 3/6 | unknown | 3/6 | 3/6 | 3/6 | 3/6 | ||
| Outermost seta of male P5 baseoendopod | unknown | half-length of median | unknown | half-length of median | half-length of median | half-length of median | 1/3 length of median | ||
| References | [20,21] | [22,23] | [24,25] | [21] | [26] | [27] | [4] | ||
| Group | I | ||||||||
| Species | P. dovi Marcus, 1966 | P. areolata Itô Tat, 1972 | P. vinosa Pallares, 1975 | P. parviseta Chang & Song, 1997 | P. jejuensis Song & Hwang, 2010 | P. xinzhengi sp. nov. | |||
| Length-width ratio of female caudal ramus | ≤1 | 1.5–2.0, with bulbous setae laterodistally and distally | ≤1 | 1 | 0.7 | ≈2, with bulbous setae laterodistally and distally | |||
| Armature formula of A2 exopod | 2(2:1,3) | 2(2:1,3) | unknown | 2(2:1,3) | 2(2:1,3) | 2(2:1,3) | |||
| Inner edge of maxilliped basis | straight or convex | concave | straight | straight | straight | concave | |||
| Length-width ratio of P1 endopod | ≈6 | ≈3.5 | ≈4 | 4.75 | ≈3.5 | ≈6.7 | |||
| Length of female P5 baseoendopod | about 2/3 length of exopod | about 2/3 length of exopod | unknown | not exceeding distal end of exopod | not exceeding distal end of exopod | slightly exceeding distal end of exopod | |||
| Rows of transversal spinules on female P5 baseoendopod | absent | present | unknown | present | absent | present | |||
| Length and armature formula of female P5 | reaching about mid-length along genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | unknown | reaching end of genital double-somite, 5/6 | reaching end of genital double-somite, 5/6 | reaching end of genital double-somite, 5/6 | |||
| Inner spine of male P1 basis | curved | curved | unknown | curved outward | normal | curved | |||
| Armature formula of male P5 | 3/6 | 3/6 | 3/6 | 3/5 | 3/5 | 3/6 | |||
| Outermost seta of male P5 baseoendopod | 1/3 length of median | 1/4 length of median | unknown | as long as median weak | half-length of median weak | 1/3 length of median | |||
| References | [3,28,29,30] | [1,10] | [3] | [1] | [2] | This paper | |||
| Group | II | ||||||||
| Species | P. clausii (Norman, 1869) | P. 16ibernica (Brady & Robertson, 1873) | P. intermedia Gurney, 1930 | P. incerta Lang, 1936 | P. paraharpactoides Lang, 1939 | P. aurantiaca Pallares, 1975 | P. patagonica Pallares, 1975 | ||
| Length-width ratio of female caudal ramus | <1 | ≤1, with bulbous seta distally | ≤1, with bulbous seta laterodistally | unknown | <1 | unknown | unknown | ||
| Armature formula of A2 exopod | 2(2:1,2) | unknown | 2(2:1,3) | unknown | unknown | unknown | unknown | ||
| Inner edge of maxilliped basis | convex | concave | straight | concave | straight or convex | concave | concave | ||
| Length-width ratio of P1 endopod | ≈4–5 | ≈4–5 | >6 | ≈4–5 | ≈4–5 | ≈4–6 | ≈4–5 | ||
| Length of female P5 baseoendopod | not exceeding distal end of exopod | slightly exceeding distal end of exopod | not exceeding distal end of exopod | unknown | unknown | unknown | unknown | ||
| Rows of transversal spinules on female P5 baseoendopod | with only one row of transverse row spinules | absent | absent | unknown | absent | absent | absent | ||
| Length and armature formula of female P5 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | 5/6 | unknown | 6/7 | 5/6 | 5/6 | ||
| Inner spine of male P1 basis | very short, curved inward | unknown | curved inward | very short, sharp and curved | unknown | unknown | very long, blunt and weakly curved | ||
| Armature formula of male P5 | 3/6 | 3/5 | 3/6 | 3/6 | 3/6 | 3/7 | 3/6 | ||
| Outermost seta of male P5 baseoendopod | as long as median | half-length of median | 2/3 length of median | slightly shorter than median | unknown | unknown | unknown | ||
| References | [22,24] | [21,24] | [31,32,33] | [3,21] | [3,21] | [3] | [3] | ||
| Group | III | ||||||||
| Species | P. affinis Scott T., 1912 | P. coatsi Scott T., 1912 | P. ganio Brehm, 1938 | P. californica Lang, 1965 | P. verrucosa Itô Tat, 1970 | P. pacificus Chislenko, 1971 | P. infesta Ho & Hong, 1988 | P. mourei Masunari, 1988 | P. yeemini Chullasorn, Kangtia & Song, 2016 |
| Length-width ratio of female caudal ramus | ≤1 | ≤1 | 1.3 | ≤1 | 1.5–1.9 | ≈1 | 1, with bulbous seta laterodistally | 3 | <1 |
| Armature formula of A2 exopod | 2(2:1,1) | 2(0:3,3) | unknown | 2(2:1,3) | 2(2:1,4)/2,4 | 2(2:1,3) | 1–2(1:1,3) | 2(2:1,3) | 2(2:1,3) |
| Inner edge of maxilliped basis | straight | convex | unknown | concave | concave | concave | concave | straight | straight |
| Length-width ratio of P1 endopod | ≈3 | ≈3 | ≈4 | ≈5 | ≈4 | ≈6 | ≈5 | ≈3.5 | ≈3.34 |
| Length of female P5 baseoendopod | about 2/3 length of exopod | slightly exceeding mid-length of exopod | unknown | about 4/5 length of exopod | not exceeding distal end of exopod | slightly exceeding distal end of exopod | exceeding distal end of exopod | exceeding distal end of exopod | slightly exceeding distal end of exopod |
| Rows of transversal spinules on female P5 baseoendopod | absent | absent | unknown | present | absent | absent | absent | absent | present |
| Length and armature formula of female P5 | unknown | unknown | unknown | reaching at least last urosomite, 5/6 | exceeding mid-length but not reaching end of genital double-somite, 5/6 | reaching at least last urosomite, 5/6 | reaching about mid-length along genital double-somite, 5/6 | reaching distal margin of genital double-somite, 5/6 | exceeding distal margin of genital double-somite, close to penultimate somite, 5/6 |
| Inner spine of male P1 basis | unknown | unknown | unknown | curved inward | normal | unknown | transformed into a heavily sclerotized L-shaped bar | curved inward | normal |
| Armature formula of male P5 | unknown | unknown | 3/7 | 3/6 | 3/5 | 3/6 | 3/6 | 3/5 | 2/5 |
| Outermost seta of male P5 baseoendopod | unknown | unknown | unknown | overreaching half-length of median | half-length of median | 2/3 length of median | 3/4 length of median | as long as median weak | / |
| References | [34] | [34] | [3] | [3,21] | [1,9,15] | [31] | [6] | [35] | [8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Ma, L.; Kou, Q. Description of a New Species of Parathalestris Brady & Robertson, 1873 (Copepoda: Harpacticoida: Thalestridae) from China, with a Key to Its Affiliated Species Group. Diversity 2023, 15, 577. https://doi.org/10.3390/d15040577
Wu Y, Ma L, Kou Q. Description of a New Species of Parathalestris Brady & Robertson, 1873 (Copepoda: Harpacticoida: Thalestridae) from China, with a Key to Its Affiliated Species Group. Diversity. 2023; 15(4):577. https://doi.org/10.3390/d15040577
Chicago/Turabian StyleWu, Yihong, Lin Ma, and Qi Kou. 2023. "Description of a New Species of Parathalestris Brady & Robertson, 1873 (Copepoda: Harpacticoida: Thalestridae) from China, with a Key to Its Affiliated Species Group" Diversity 15, no. 4: 577. https://doi.org/10.3390/d15040577
APA StyleWu, Y., Ma, L., & Kou, Q. (2023). Description of a New Species of Parathalestris Brady & Robertson, 1873 (Copepoda: Harpacticoida: Thalestridae) from China, with a Key to Its Affiliated Species Group. Diversity, 15(4), 577. https://doi.org/10.3390/d15040577