

A New Record of Pinctada fucata (Bivalvia: Pterioida: Pteriidae) in Mischief Reef: A Potential Invasive Species in the Nansha Islands, China

 ,
,
Abstract
:1. Introduction
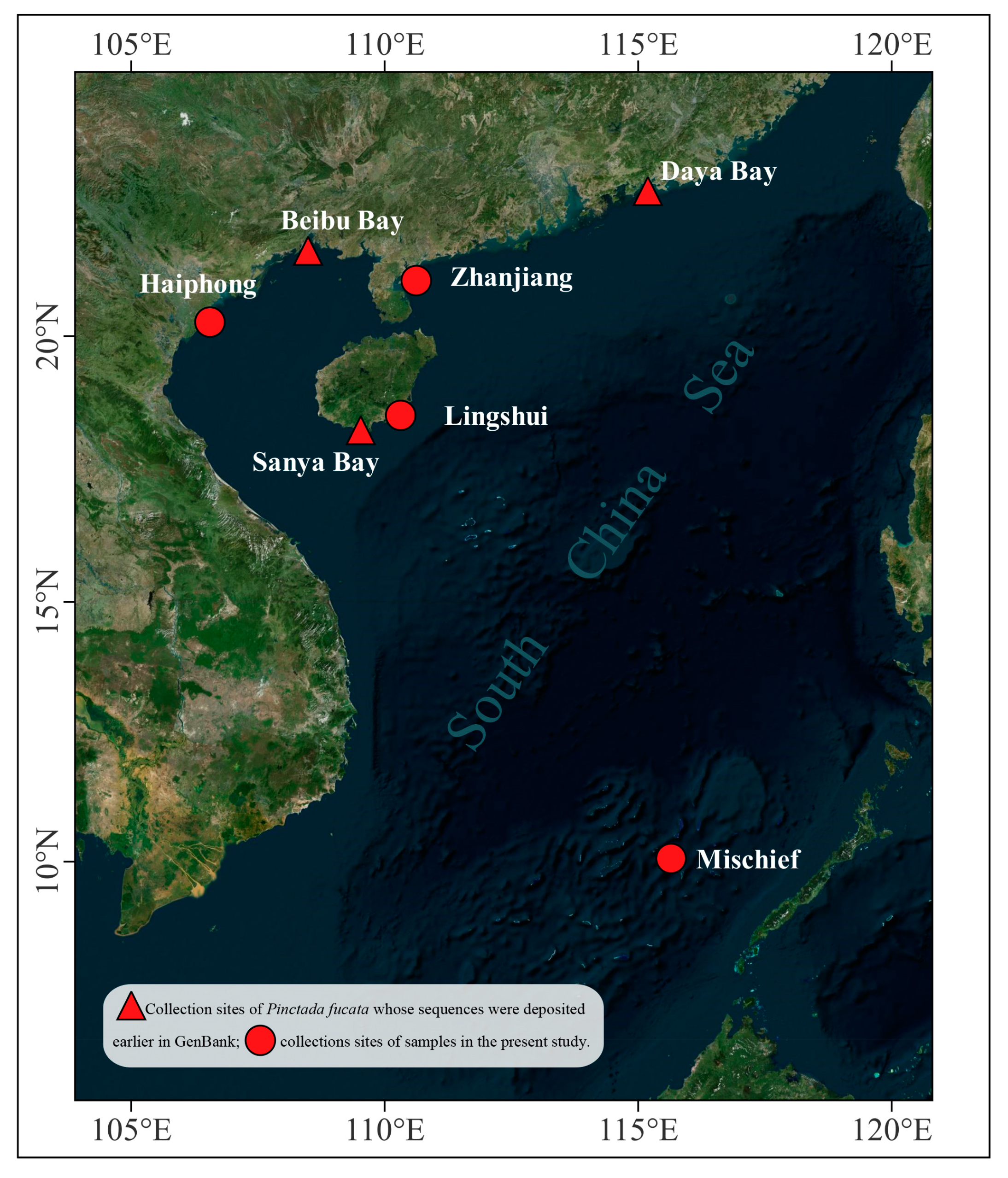
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Amplification and Sequencing
2.3. Statistical Anazlysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
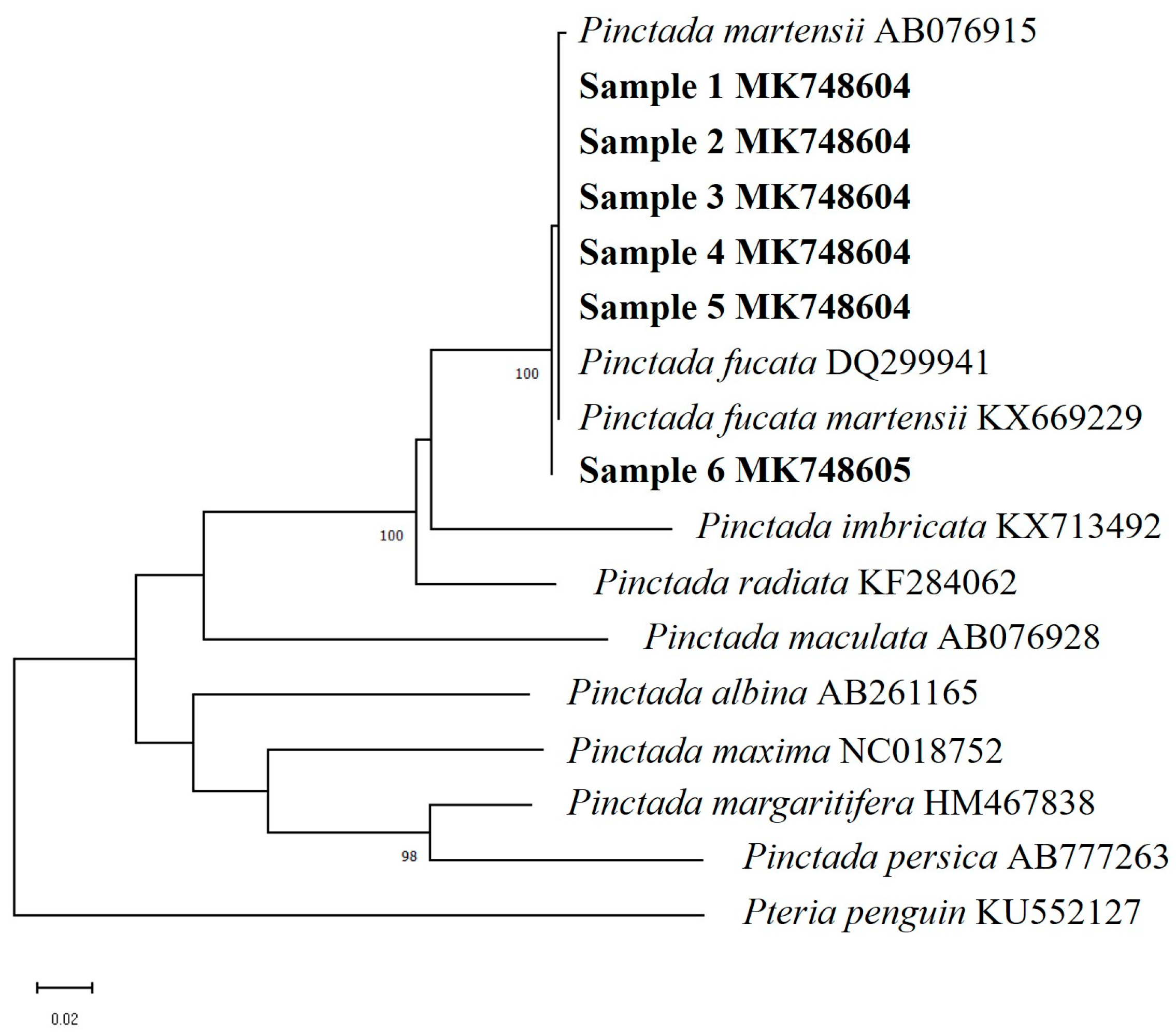
| Species | GenBank Accession No. | Sample Site |
|---|---|---|
| Pinctada fucata martensii * | KX669229 | Sanya, China [32] |
| Pinctada fucata | DQ299941 | China |
| Pinctada albina | AB261165 | Kagoshima, Japan [33] |
| Pinctada martensii * | AB076915 | Okinawa, Japan [18] |
| Pinctada imbricata | KX713492 | Florida Keys, USA [34] |
| Pinctada maculata | AB076928 | Okinawa, Japan [18] |
| Pinctada margaritifera | HM467838 | China |
| Pinctada persica | AB777263 | Hendurabi, Iran [35] |
| Pinctada radiata | KF284062 | Ras Al Khaimah, UAE [36] |
| Pinctada maxima | NC018752 | Not mentioned [37] |
| Pteria penguin | KU552127 | Sanya, China [32] |
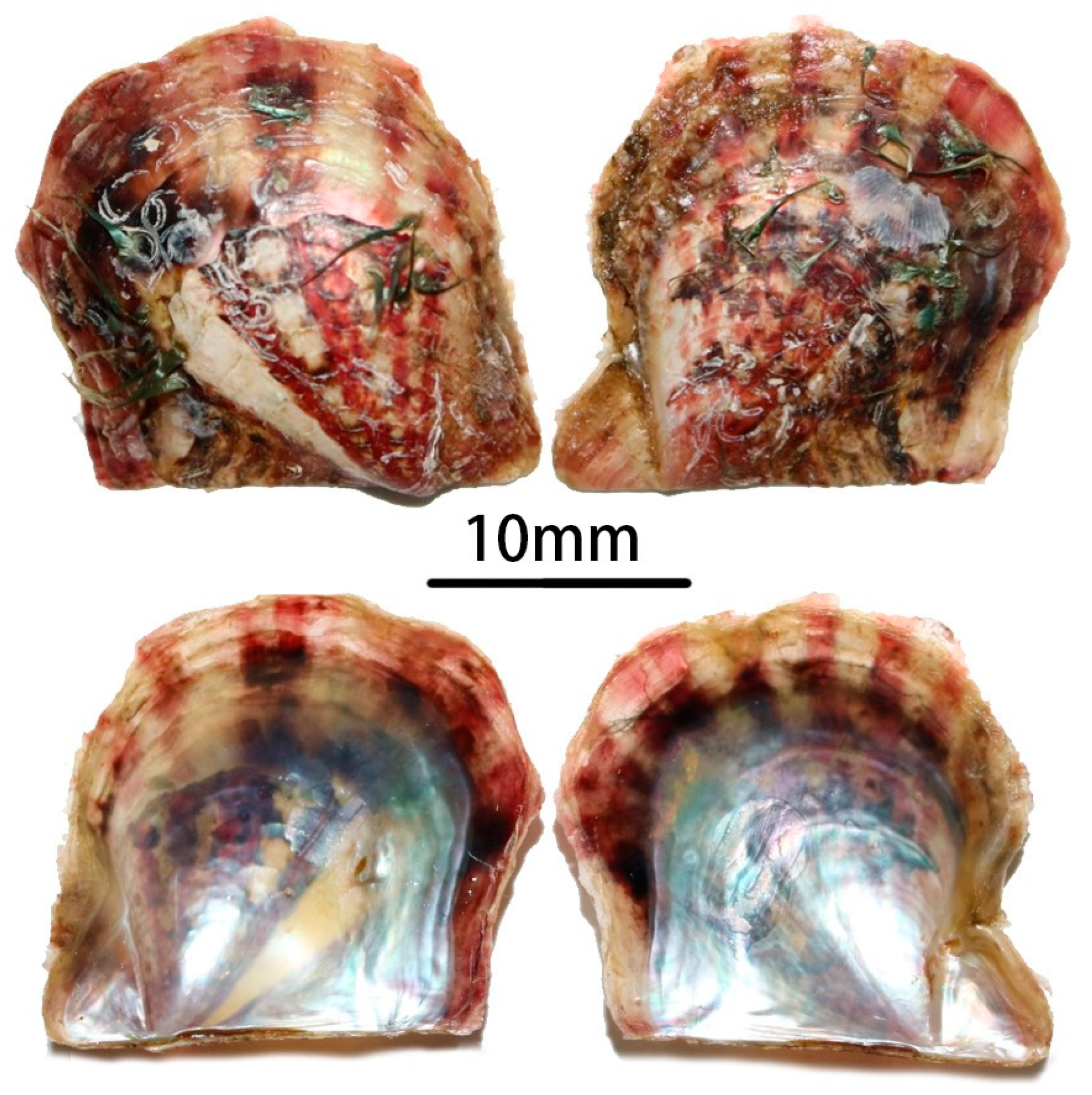
3. Results
3.1. Morphological Characteristics and DNA Barcoding Identification
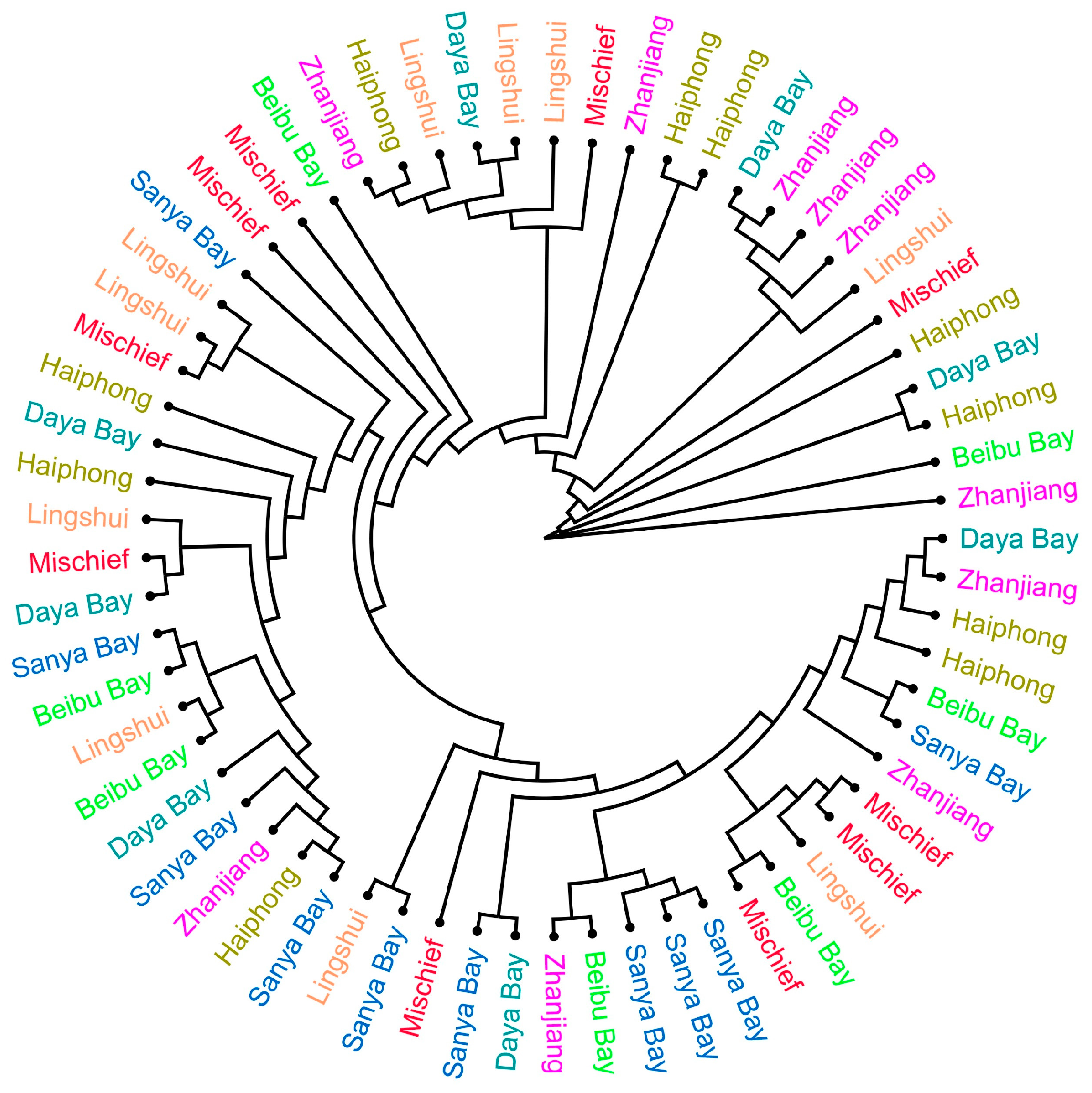
3.2. Population Genetic Diversity
3.3. Population Genetic Structure and Differentiation
4. Discussion
4.1. Identification of the Samples
4.2. Population Genetics Analysis
4.3. Possible Cause of Occurrence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tong, X.; Wang, R.; Chen, X.Y. Expansion or Invasion? A Response to Nackley et al. Trends. Ecol. Evol. 2018, 33, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Di Castri, F.; Drake, J.A.; Mooney, H.A.; Groves, R.H.; Kruger, F.J.; Rejmanek, M.; Williamson, M. History of biological invasions with emphasis on the Old World. In Biological Invasions: A Global Perspective; Wiley: Chichester, UK, 1989; pp. 1–30. [Google Scholar]
- Werschkun, B.; Banerji, S.; Basurko, O.C.; David, M.; Fuhr, F.; Gollasch, S.; Grummt, T.; Haarich, M.; Jha, A.N.; Kacan, S.; et al. Emerging risks from ballast water treatment: The run-up to the International Ballast Water Management Convention. Chemosphere 2014, 112, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Bax, N.; Williamson, A.; Aguero, M.; Gonzalez, E.; Geeves, W. Marine invasive alien species: A threat to global biodiversity. Mar. Policy 2003, 27, 313–323. [Google Scholar] [CrossRef]
- Bortolus, A.; Carlton, J.T.; Schwindt, E. Reimagining South American coasts: Unveiling the hidden invasion history of an iconic ecological engineer. Divers. Distrib. 2015, 21, 1267–1283. [Google Scholar] [CrossRef]
- Blakeslee, A.M.H. Parasites and genetics in marine invertebrate introductions: Signatures of diversity declines across systems. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning-Clode, J., Ed.; Sciendo Migration: Warsaw, Poland, 2016; pp. 138–182. [Google Scholar]
- Southgate, P.C.; Lucas, J.S. The Pearl Oyster; Elsevier Science: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Yu, D.H.; Zhu, J.H. Species status of Chinese Pinctada fucata, Japanese P. fucata martensii and Australian P. imbricata using ITS and AFLP markers. South China Fish. Sci. 2006, 2, 36–44. [Google Scholar]
- Yu, D.H.; Chu, K.H.; Jia, X.P. Preliminary analysis on genetic relationship of common pearl oysters of Pinctada in China. Oceanol. Limnol. Sin. 2006, 37, 211–217. [Google Scholar]
- Cunha, R.L.; Blanc, F.; Bonhomme, F.; Arnaud-Haond, S. Evolutionary patterns in pearl oysters of the genus Pinctada (Bivalvia: Pteriidae). Mar. Biotechnol. 2011, 13, 181–192. [Google Scholar] [CrossRef]
- Choi, Y.H.; Chang, Y.J. Gametogenic cycle of the transplanted-cultured pearl oyster, Pinctada fucata martensii (Bivalvia: Pteriidae) in Korea. Aquaculture 2003, 220, 781–790. [Google Scholar] [CrossRef]
- Tomaru, Y.; Udaka, N.; Kawabata, Z.; Nakano, S. Seasonal change of seston size distribution and phytoplankton composition in bivalve pearl oyster Pinctada fucata martensii culture farm. Hydrobiologia 2002, 481, 181–185. [Google Scholar] [CrossRef]
- Wang, Z.R.; Chen, R.Q. The species of the Pterioda from the Nansha Islands waters. In Studies on Marine Species of the Nansha Islands and Neighboring Waters; Chen, Q.C., Ed.; China Ocean Press: Beijing, China, 1991; pp. 150–151. [Google Scholar]
- Hynd, J.S. A Revision of the Australian Pearl-shells, Genus Pinctada (Lamelli-branchia). Mar. Freshwater. Res. 1955, 6, 98–138. [Google Scholar] [CrossRef]
- Waller, T.R. Morphology, morphoclines and a new classification of the Pteriomorphia (Mollusca: Bivalvia). Philos. Trans. R. Soc. B 1978, 284, 345–365. [Google Scholar]
- Masaoka, T.; Kobayashi, T. Natural hybridization between Pinctada fucata and Pinctada maculata inferred from internal transcribed spacer regions of nuclear ribosomal RNA genes. Fish. Sci. 2005, 71, 829–836. [Google Scholar] [CrossRef]
- Wada, K.T.; Tëmkin, I. Taxonomy and Phylogeny. In The Pearl Oyster; Southgate, P.C., Lucas, J.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Matsumoto, M. Phylogenetic analysis of the subclass Pteriomorphia (Bivalvia) from mtDNA COI sequences. Mol. Phylogenet. Evol. 2003, 27, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.R.; Apte, S.; MacAvoy, E.S.; Gardner, J.P. A molecular phylogeny of the marine mussel genus Perna (Bivalvia: Mytilidae) based on nuclear (ITS1&2) and mitochondrial (COI) DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 685–698. [Google Scholar] [PubMed]
- Gavrilović, A.; Piria, M.; Guo, X.Z.; Jug-Dujaković, J.; Ljubučić, A.; Krkić, A.; Iveša, N.; Marshall, B.A.; Gardner, J.P. First evidence of establishment of the rayed pearl oyster, Pinctada imbricata radiata (Leach, 1814), in the eastern Adriatic Sea. Mar. Pollut. Bull. 2017, 125, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Somrup, S.; Sangsawang, A.; McMillan, N.; Winitchai, S.; Inthoncharoen, J.; Liu, S.; Muangmai, N. Pinctada phuketensis sp. nov. (Bivalvia, Ostreida, Margaritidae), a new pearl oyster species from Phuket, western coast of Thailand. ZooKeys 2022, 1119, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Muirhead, J.R.; Gray, D.K.; Kelly, D.W.; Ellis, S.M.; Heath, D.D.; Macisaac, H.J. Identifying the source of species invasions: Sampling intensity vs. genetic diversity. Mol. Ecol. 2008, 17, 1020–1035. [Google Scholar] [CrossRef]
- Cristescu, M.E. Genetic reconstructions of invasion history. Mol. Ecol. 2015, 24, 2212–2225. [Google Scholar] [CrossRef]
- Wu, L. Studies of Genetic Diversity on Pinctada martensii and Sinonovacula Constricta; China Ocean University: Qingdao, China, 2013. [Google Scholar]
- Yu, D.H.; Chu, K.H. Species identity and phylogenetic relationship of the pearl oysters in Pinctada Röding, 1798 based on ITS sequence analysis. Biochem. Syst. Ecol. 2006, 34, 240–250. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate base substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Zhan, X.; Zhang, S.; Gu, Z.; Wang, A. Complete mitochondrial genomes of two pearl oyster species (Bivalvia: Pteriomorphia) reveal novel gene arrangements. J. Shellfish Res. 2018, 37, 1039–1050. [Google Scholar] [CrossRef]
- Takakura, D.; Wada, K.T.; Kobayashi, T.; Masaoka, T.; Samata, T. Phylogenetic relationships of several pearl oyster species in Pinctada (Pteriidae; Bivalvia) inferred from mitochondrial 12S rDNA and cytochrome c oxidase subunit I genes sequences. J. Foss. Res. 2008, 41, 33–40. [Google Scholar]
- Combosch, D.J.; Collins, T.M.; Glover, E.A.; Daniel, L.G.; Elizabeth, M.H.; John, M.H.; Gisele, Y.K.; Sarah, L.; Erin, M.; Ellen, E.S.; et al. A family-level tree of life for bivalves based on a Sanger-sequencing approach. Mol. Phylogenet. Evol. 2017, 107, 191–208. [Google Scholar] [CrossRef]
- Sharif Ranjbar, M.; Zolgharnien, H.; Yavari, V.; Bita, A.; Mohammad, A.S.; Sophie, A.H.; Regina, L.C. Rising the Persian Gulf black-lip pearl oyster to the species level: Fragmented habitat and chaotic genetic patchiness in Pinctada persica. Evol. Biol. 2016, 43, 131–143. [Google Scholar] [CrossRef]
- Meyer, J.B.; Cartier, L.E.; Pinto-Figueroa, E.A.; Michael, S.K.; Henry, A.H.; Bruce, A.M. DNA fingerprinting of pearls to determine their origins. PLoS ONE 2013, 8, e75606. [Google Scholar] [CrossRef]
- Wu, X.; Li, X.; Li, L.; Yu, Z. A unique tRNA gene family and a novel, highly expressed ORF in the mitochondrial genome of the silver-lip pearl oyster, Pinctada maxima (Bivalvia: Pteriidae). Gene 2012, 510, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Hayami, I. Superfamily pteriacea. In Marine Mollusks in Japan; Okutani, T., Ed.; Tokai University: Tokyo, Japan, 2000; pp. 879–883. [Google Scholar]
- Takemura, Y.; Okutani, T. On the identification of species of Pinctada found attached to Pinctada maxima (Jameson) in the Arafura Sea. Bull. Tokai Reg. Fish. Res. Lab. 1958, 20, 47–60. [Google Scholar]
- Chen, Q.C. Studies on Marine Biodiversity of the Nasha Islands and Neighboring Waters I.I.; China Ocean Press: Beijing, China, 1996; p. 101. [Google Scholar]
- Sousa, R.; Gutiérrez, J.; Aldridge, D. Non-indigenous invasive bivalves as ecosystem engineers. Biol. Invas. 2009, 11, 2367–2385. [Google Scholar] [CrossRef]
- Terranova, M.S.; Brutto, S.L.; Arculeo, M.; Mitton, J.B. Population structure of Brachidontes pharaonis (P. Fisher, 1870) (Bivalvia, Mytilidae) in the Mediterranean Sea, and evolution of a novel mtDNA polymorphism. Mar. Biol. 2006, 150, 89–101. [Google Scholar] [CrossRef]
- Fernández-Pérez, J.; Froufe, E.; Nantón, A.; Gaspar, M.B.; Méndez, J. Genetic diversity and population genetic analysis of Donax vittatus (Mollusca: Bivalvia) and phylogeny of the genus with mitochondrial and nuclear markers. Estuar. Coast. Shelf Sci. 2017, 197, 126–135. [Google Scholar] [CrossRef]
- Takeuchi, T.; Masaoka, T.; Aoki, H.; Koyanagi, R.; Fujie, M.; Satoh, N. Divergent northern and southern populations and demographic history of the pearl oyster in the western Pacific revealed with genomic SNPs. Evol. Appl. 2019, 13, 837–853. [Google Scholar] [CrossRef]
- Gerlach, G.; Jueterbock, A.; Kraemer, P.; Deppermann, J.; Harmand, P. Calculations of population differentiation based on GST and D: Forget GST but not all of statistics. Mol. Ecol. 2010, 19, 3845–3852. [Google Scholar] [CrossRef]
- Chen, J.; Luo, H.; Zhai, Z.; Wang, H.; Liu, B.; Bai, L.; Yu, D. Estimates of additive and non-additive genetic effects on growth traits in a diallel cross of three strains of pearl oyster (Pinctada fucata). Aquacult. Int. 2021, 29, 1359–1371. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Fofonoff, P.W.; Steves, B.P.; Carlton, J.T. Invasion history and vector dynamics in coastal marine ecosystems: A North American perspective. Aquat. Ecosyst. Health 2015, 18, 299–311. [Google Scholar] [CrossRef]
- Chang, Y.Q. Aquaculture of Shellfish; China Agriculture Press: Beijing, China, 2017. [Google Scholar]
- Chen, M.; Chen, X.; Li, Y.; Wu, Q.; Yu, G.; Ma, Z.; Xing, K.; Wang, L. Cage culture of pearl oyster Pinctada martensii in Meiji Reef. Fish. Sci. 2014, 35, 379–383. (In Chinese) [Google Scholar]
- Cheng, C. Meiji Reef grouper pelagic culture, a beautiful aquatic scenery line. Curr. Fish. 2014, 39, 62–63. (In Chinese) [Google Scholar]
- Marques, R.C.; Breves, A. First record of Pinctada imbricata Röding, 1798 (Bivalvia: Pteroidea) attached to a rafting item: A potentially invasive species on the Uruguayan coast. Mar. Biodivers. 2015, 45, 333–337. [Google Scholar] [CrossRef]
- Nackley, L.L.; West, A.G.; Skowno, A.L.; Bond, W.J. The nebulous ecology of native invasions. Trends. Ecol. Evol. 2017, 32, 814–824. [Google Scholar] [CrossRef] [PubMed]





| Zhanjiang | Lingshui | Mischief | Haiphong | |
|---|---|---|---|---|
| Numbers of haplotypes | 8 | 10 | 10 | 9 |
| Haplotype diversity | 0.9333 | 1.0000 | 1.0000 | 0.9778 |
| Nucleotide diversity | 0.0129 | 0.0148 | 0.0151 | 0.0142 |
| Mean number of pairwise difference | 7.1556 | 8.1556 | 8.3111 | 7.8222 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, B.; Deng, Z.; Ma, S.; Sun, D.; Liu, Y.; Yang, C.; Wu, Q.; Yu, G. A New Record of Pinctada fucata (Bivalvia: Pterioida: Pteriidae) in Mischief Reef: A Potential Invasive Species in the Nansha Islands, China. Diversity 2023, 15, 578. https://doi.org/10.3390/d15040578
Shan B, Deng Z, Ma S, Sun D, Liu Y, Yang C, Wu Q, Yu G. A New Record of Pinctada fucata (Bivalvia: Pterioida: Pteriidae) in Mischief Reef: A Potential Invasive Species in the Nansha Islands, China. Diversity. 2023; 15(4):578. https://doi.org/10.3390/d15040578
Chicago/Turabian StyleShan, Binbin, Zhenghua Deng, Shengwei Ma, Dianrong Sun, Yan Liu, Changping Yang, Qiaer Wu, and Gang Yu. 2023. "A New Record of Pinctada fucata (Bivalvia: Pterioida: Pteriidae) in Mischief Reef: A Potential Invasive Species in the Nansha Islands, China" Diversity 15, no. 4: 578. https://doi.org/10.3390/d15040578
APA StyleShan, B., Deng, Z., Ma, S., Sun, D., Liu, Y., Yang, C., Wu, Q., & Yu, G. (2023). A New Record of Pinctada fucata (Bivalvia: Pterioida: Pteriidae) in Mischief Reef: A Potential Invasive Species in the Nansha Islands, China. Diversity, 15(4), 578. https://doi.org/10.3390/d15040578