
Fungal Diversity and Dynamics during Long-Term Immersion of Conventional and Biodegradable Plastics in the Marine Environment

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
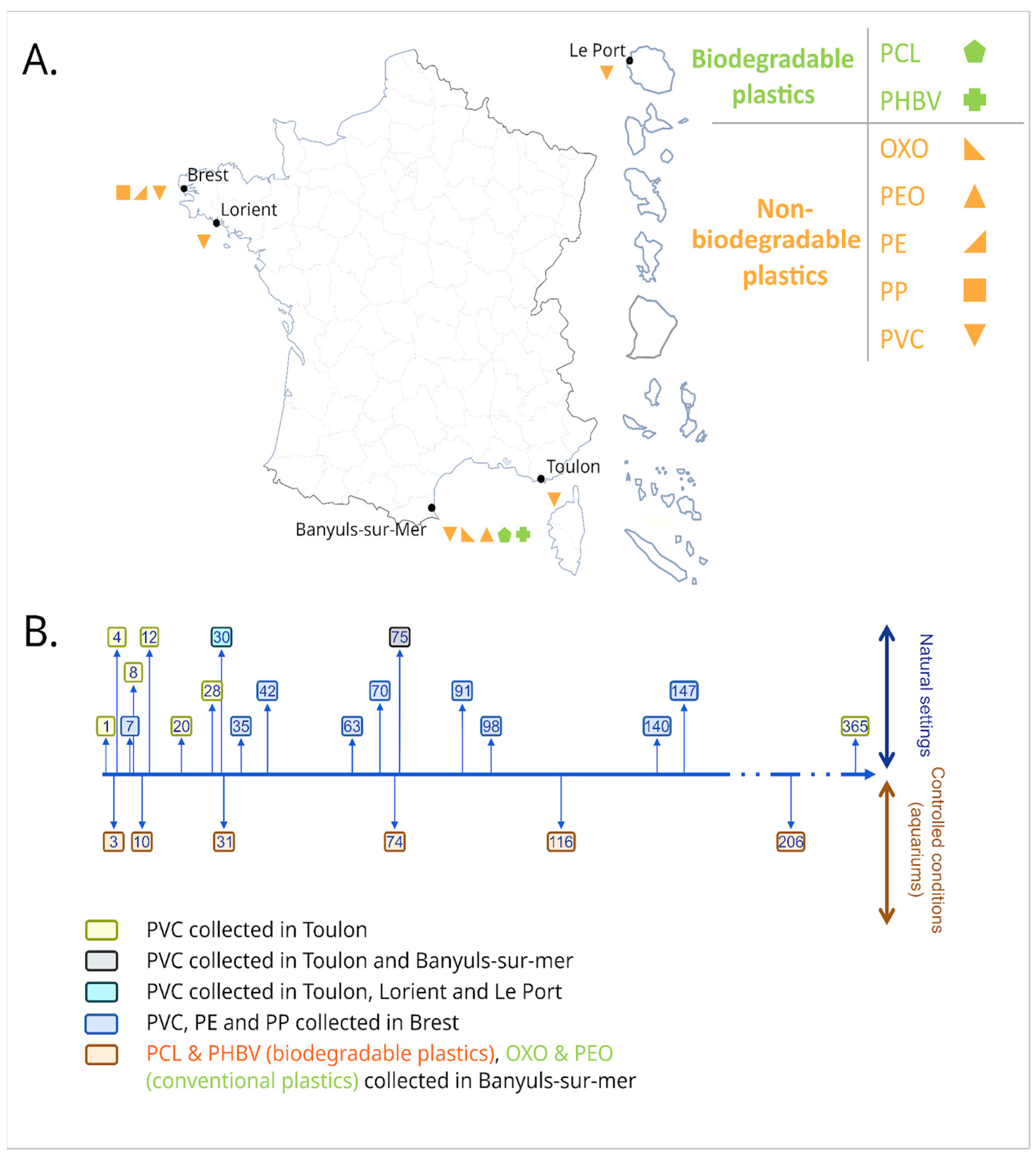
2.1. Sample Collection
2.2. DNA Extraction, Amplification and Sequencing
2.3. Bioinformatics and Statistical Analyses
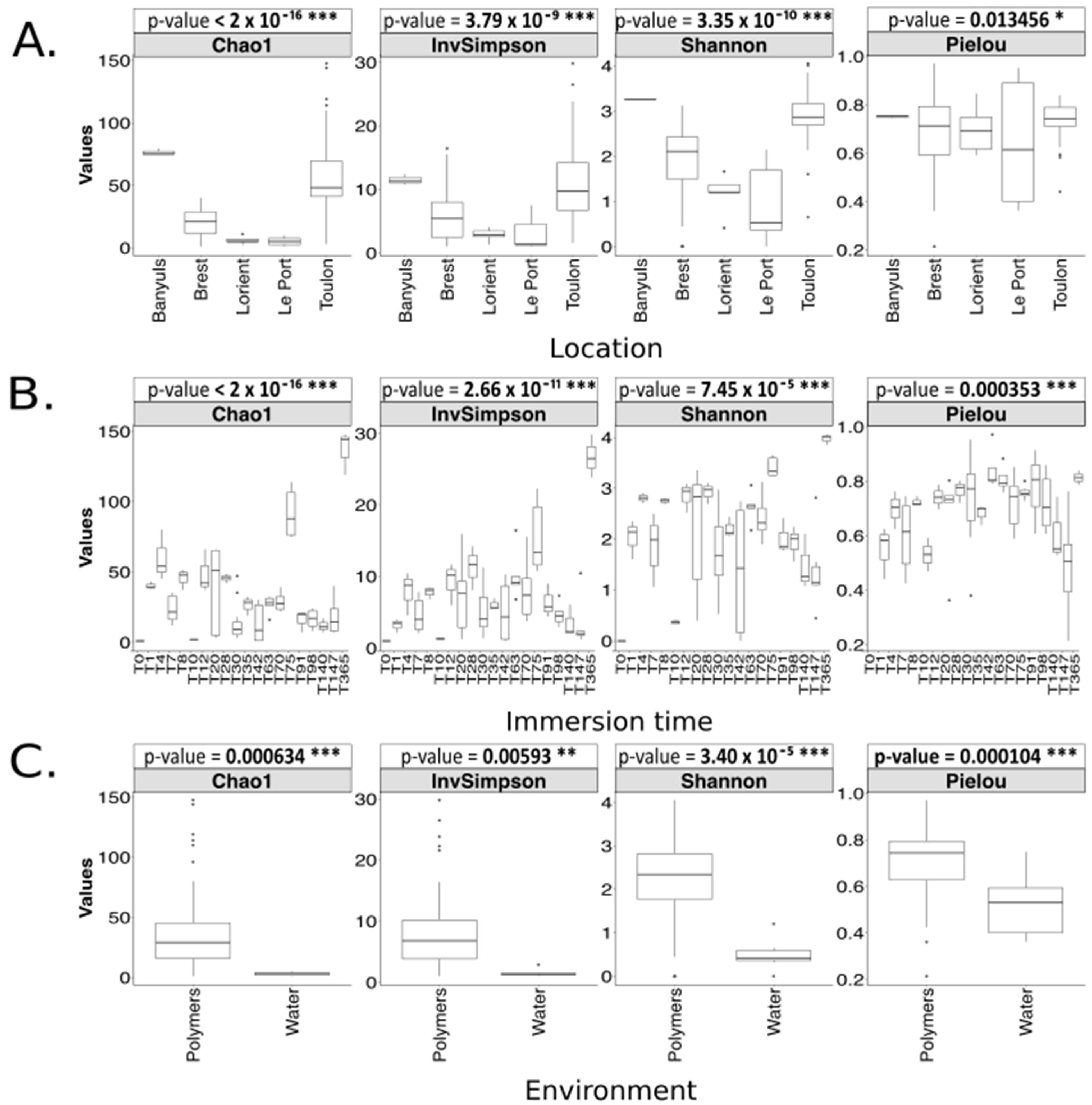
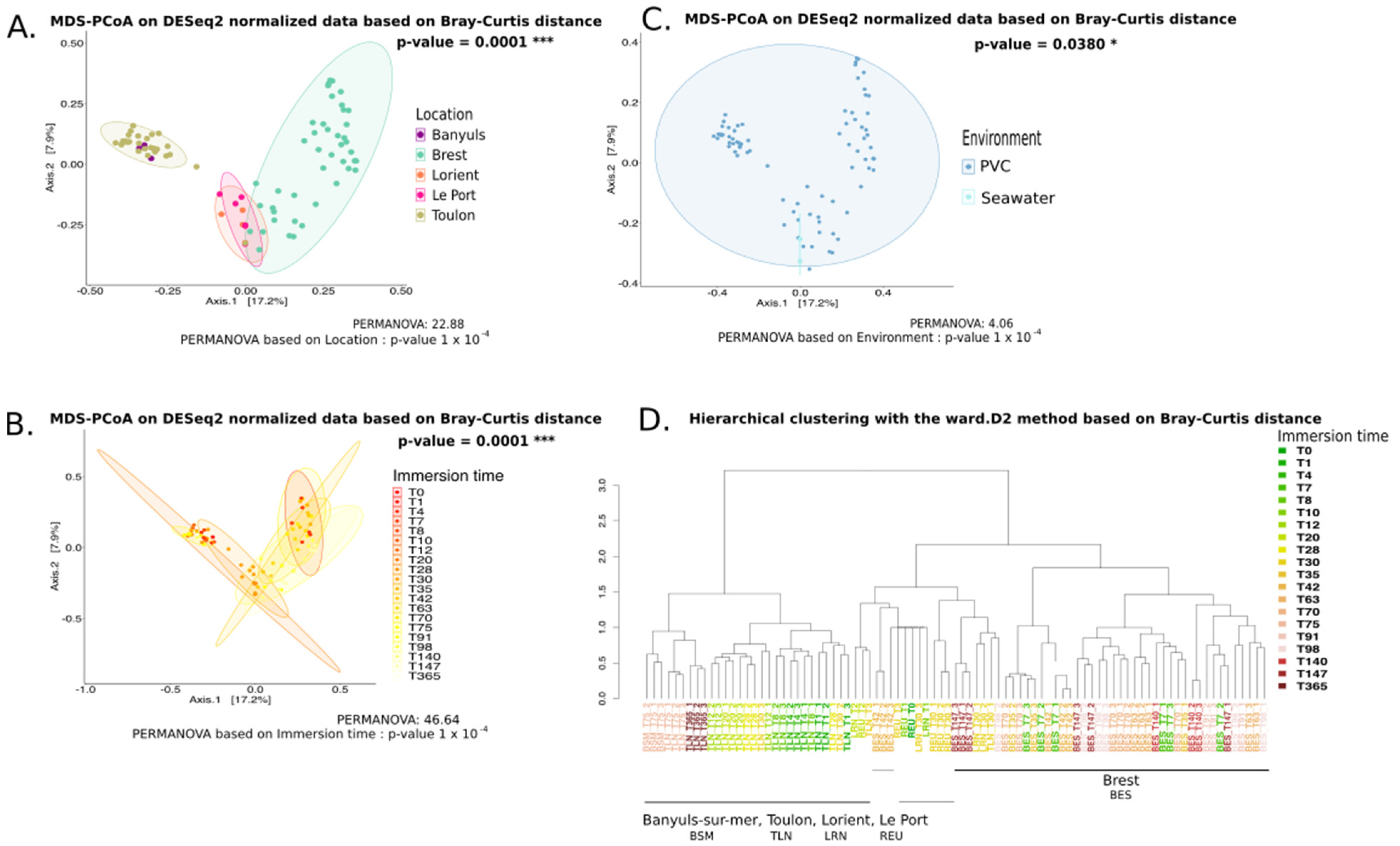
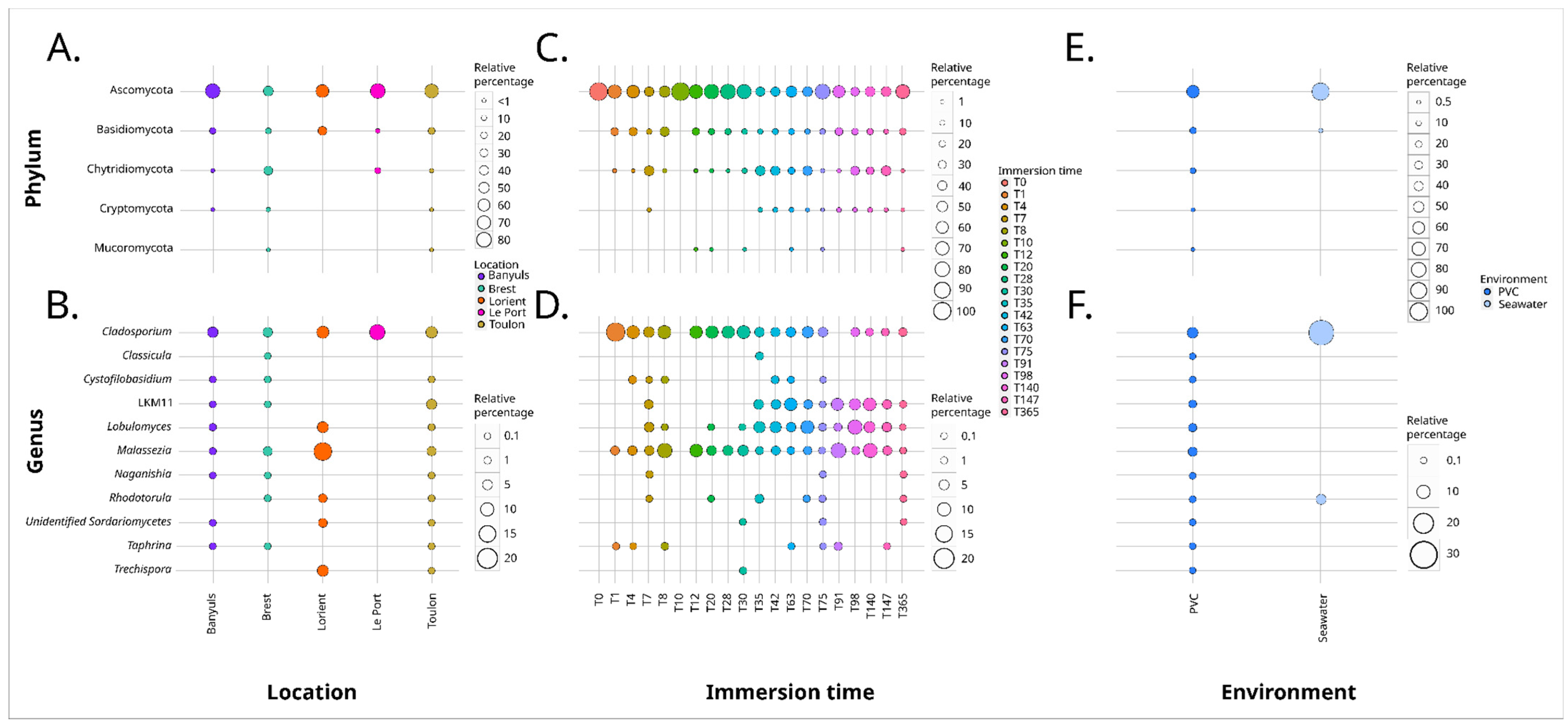
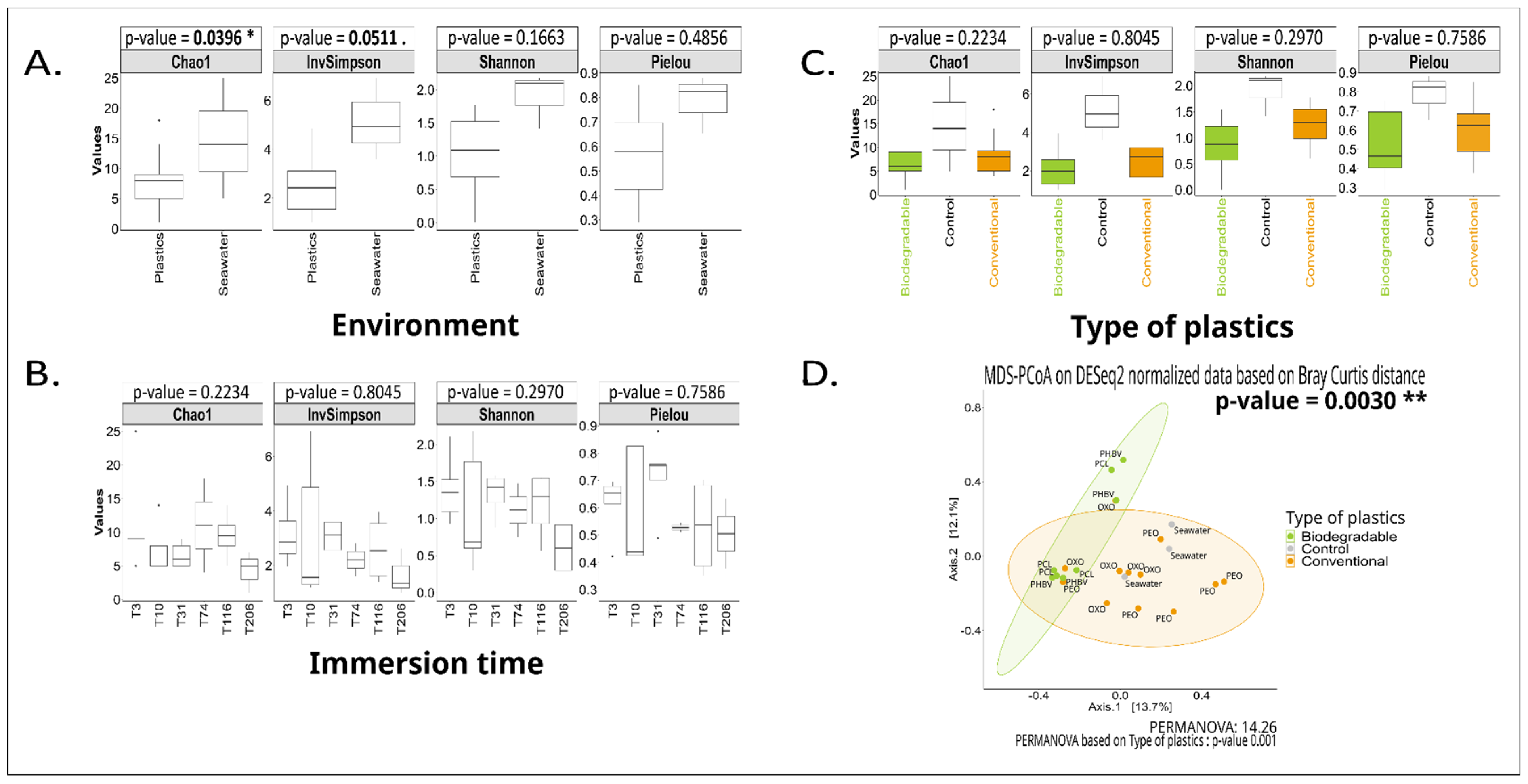
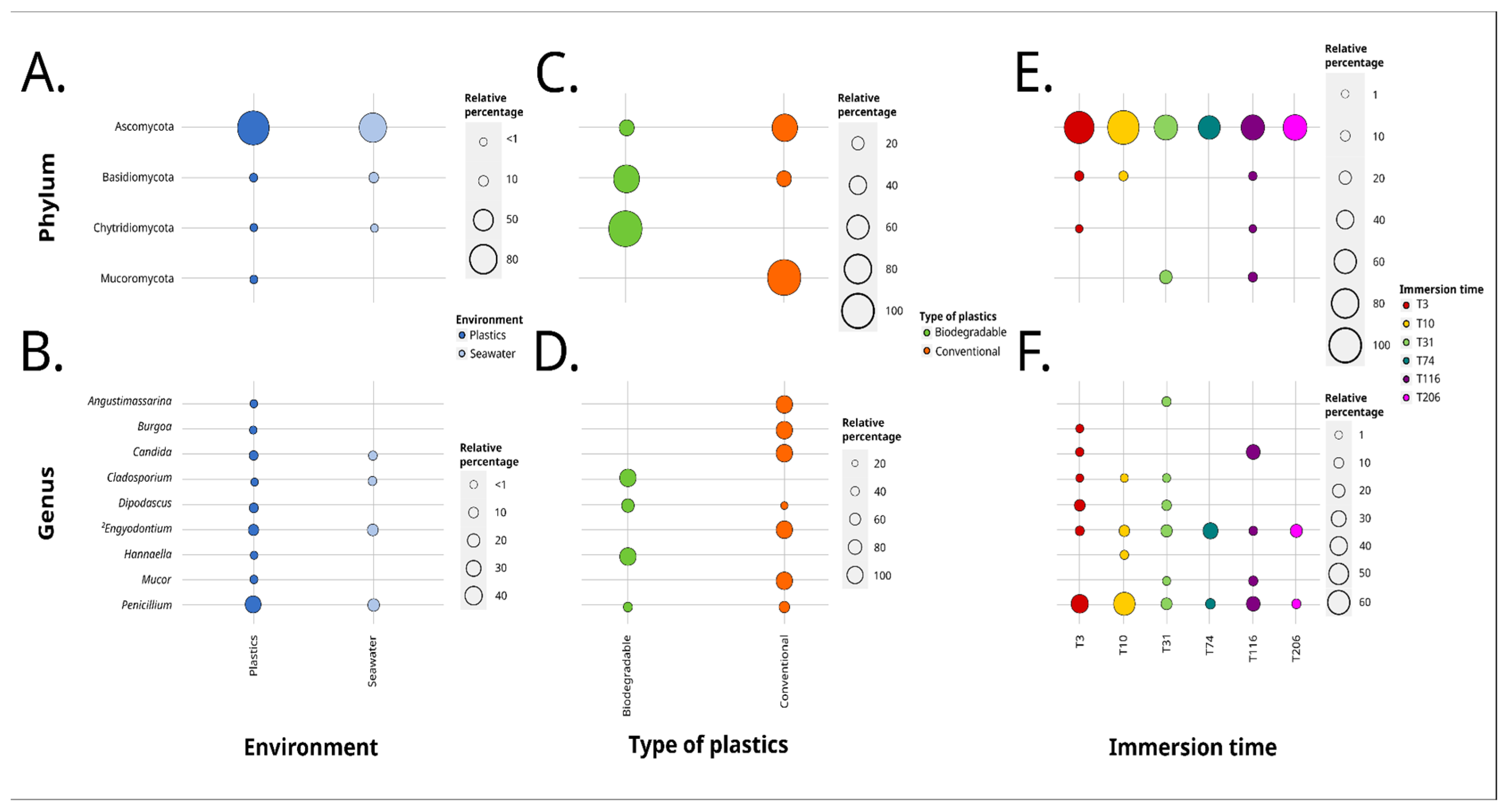
3. Results
3.1. Long-Term Immersion of PVC in Natural Settings
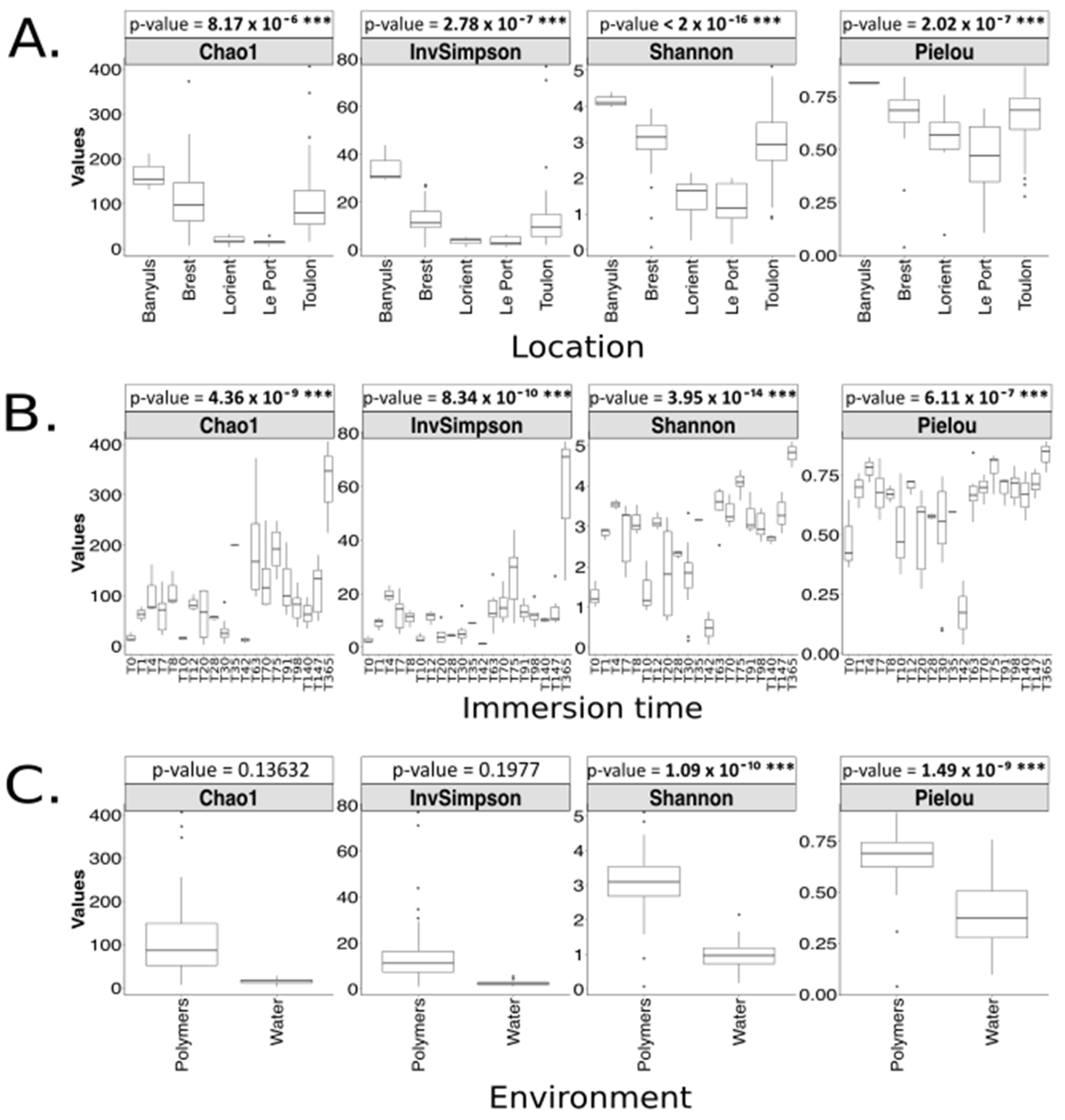
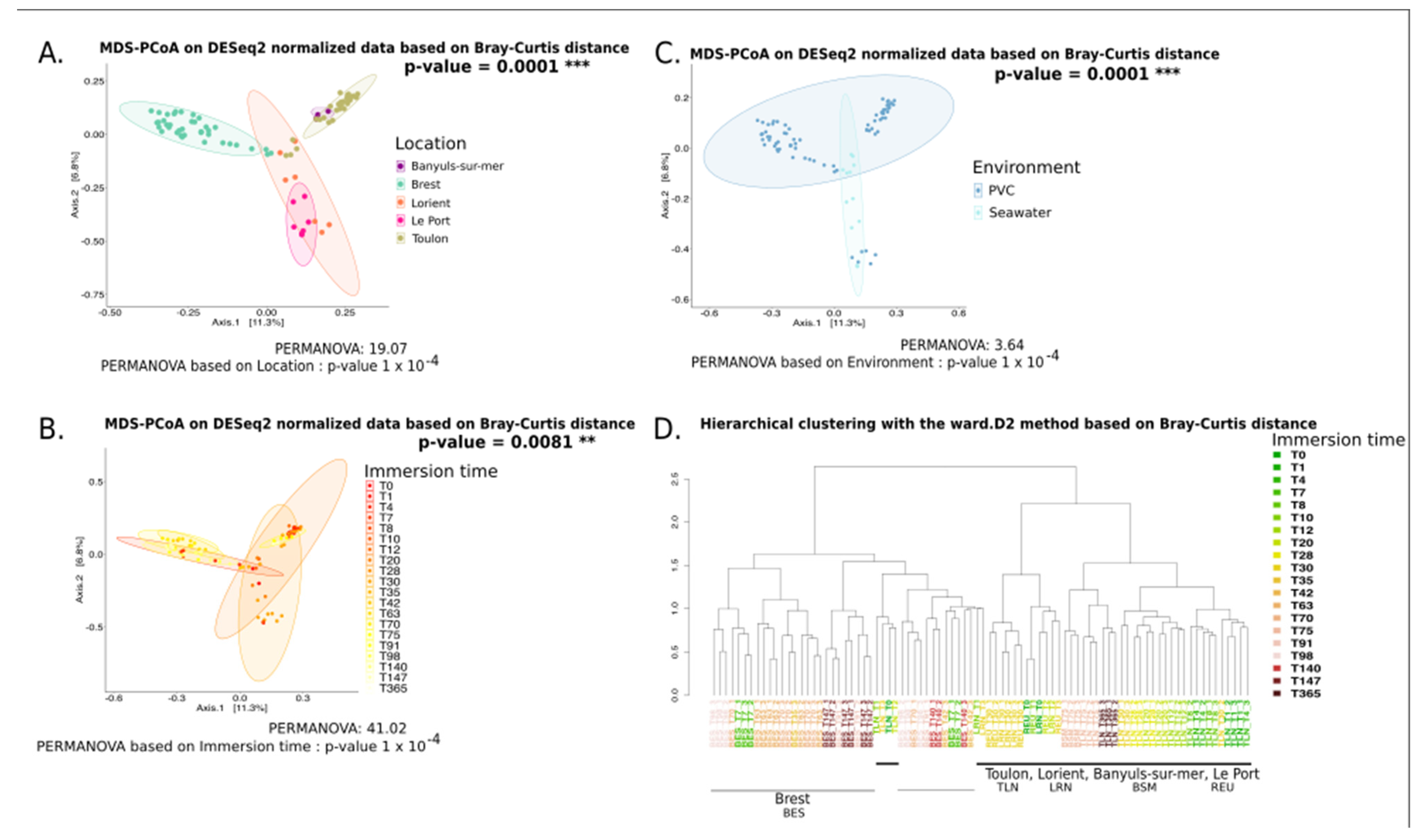
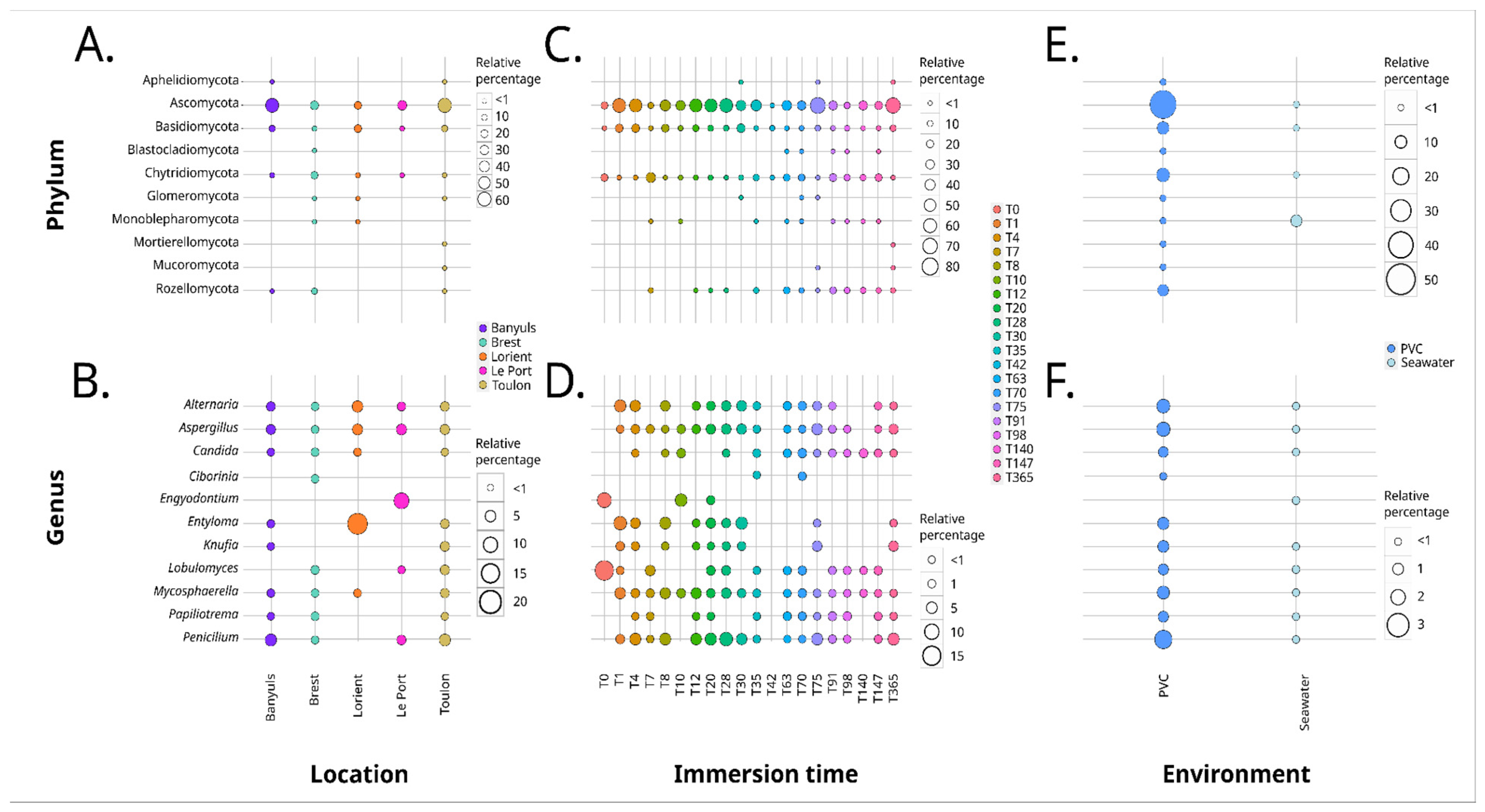
3.1.1. Analysis Based on the ITS2 Dataset
3.1.2. Analysis Based on the V4 Region of the 18S rRNA Gene
3.2. Long-Term Immersion of Plastics in Controlled Conditions
4. Discussion
4.1. Plastics Influence Marine Fungal Communities
4.2. The Biodegradable and Conventional Nature of Polymers Impacts the Plastic-Associated Fungal Community
4.3. Geographical Location Impacts the Plastic-Associated Fungal Community
4.4. Immersion Duration Impacts the Plastic-Associated Fungal Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plastics Europe Plastics—The Facts 2020: An Analysis of European Plastics Production, Demand and Waste Data 2020. Available online: https://plasticseurope.org/knowledge-hub/plastics-the-facts-2020/ (accessed on 4 March 2023).
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic Waste Inputs from Land into the Ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Regoli, F. Plastics and Microplastics in the Oceans: From Emerging Pollutants to Emerged Threat. Mar. Environ. Res. 2017, 128, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H.; Roser, M. Plastic Pollution. Our World Data 2018; UN Environment Programme: Nairobi, Kenya, 2018. [Google Scholar]
- Ruiz, I.; Ana J., A.; Basurko, O.C.; Rubio, A. Modelling the Distribution of Fishing-Related Floating Marine Litter within the Bay of Biscay and Its Marine Protected Areas. Environ. Pollut. 2022, 292, 118216. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Kunz, A.; Hu, C.-S.; Yen, N.; Lin, H.-T. Rapid-Survey Methodology to Assess Litter Volumes along Large River Systems—A Case Study of the Tamsui River in Taiwan. Sustainability 2021, 13, 8765. [Google Scholar] [CrossRef]
- UNEP. From Pollution to Solution: A Global Assessment of Marine Litter and Plastic Pollution 2021. Available online: https://www.unep.org/resources/pollution-solution-global-assessment-marine-litter-and-plastic-pollution (accessed on 4 March 2023).
- Zalasiewicz, J.; Waters, C.N.; Ivar do Sul, J.A.; Corcoran, P.L.; Barnosky, A.D.; Cearreta, A.; Edgeworth, M.; Gałuszka, A.; Jeandel, C.; Leinfelder, R.; et al. The Geological Cycle of Plastics and Their Use as a Stratigraphic Indicator of the Anthropocene. Anthropocene 2016, 13, 4–17. [Google Scholar] [CrossRef]
- Porta, R. Anthropocene, the Plastic Age and Future Perspectives. FEBS Open Bio. 2021, 11, 948–953. [Google Scholar] [CrossRef]
- García Rellán, A.; Vázquez Ares, D.; Vázquez Brea, C.; Francisco López, A.; Bello Bugallo, P.M. Sources, Sinks and Transformations of Plastics in Our Oceans: Review, Management Strategies and Modelling. Sci. Total Environ. 2023, 854, 158745. [Google Scholar] [CrossRef]
- Liu, X.; Gao, C.; Sangwan, P.; Yu, L.; Tong, Z. Accelerating the Degradation of Polyolefins through Additives and Blending. J. Appl. Polym. Sci. 2014, 131, 1–15. [Google Scholar] [CrossRef]
- O’Brine, T.; Thompson, R.C. Degradation of Plastic Carrier Bags in the Marine Environment. Mar. Pollut. Bull. 2010, 60, 2279–2283. [Google Scholar] [CrossRef]
- Alvarez-Zeferino, J.C.; Beltrán-Villavicencio, M.; Vázquez-Morillas, A. Degradation of Plastics in Seawater in Laboratory. Open J. Polym. Chem. 2015, 5, 55–62. [Google Scholar] [CrossRef]
- Sharma, P.; Gaur, V.K.; Sirohi, R.; Varjani, S.; Hyoun Kim, S.; Wong, J.W.C. Sustainable Processing of Food Waste for Production of Bio-Based Products for Circular Bioeconomy. Bioresour. Technol. 2021, 325, 124684. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Jones, B.H. Roadmap to Biodegradable Plastics—Current State and Research Needs. ACS Sustain. Chem. Eng. 2021, 9, 6170–6187. [Google Scholar] [CrossRef]
- Havstad, M.R. Biodegradable Plastics. In Plastic Waste and Recycling; Elsevier: Amsterdam, The Netherlands, 2020; pp. 97–129. ISBN 978-0-12-817880-5. [Google Scholar]
- Melchor-Martínez, E.M.; Macías-Garbett, R.; Alvarado-Ramírez, L.; Araújo, R.G.; Sosa-Hernández, J.E.; Ramírez-Gamboa, D.; Parra-Arroyo, L.; Alvarez, A.G.; Monteverde, R.P.B.; Cazares, K.A.S.; et al. Towards a Circular Economy of Plastics: An Evaluation of the Systematic Transition to a New Generation of Bioplastics. Polymers 2022, 14, 1203. [Google Scholar] [CrossRef] [PubMed]
- Jacquin, J.; Callac, N.; Cheng, J.; Giraud, C.; Gorand, Y.; Denoual, C.; Pujo-Pay, M.; Conan, P.; Meistertzheim, A.-L.; Barbe, V.; et al. Microbial Diversity and Activity during the Biodegradation in Seawater of Various Substitutes to Conventional Plastic Cotton Swab Sticks. Front. Microbiol. 2021, 12, 604395. [Google Scholar] [CrossRef]
- Deroiné, M.; César, G.; Duigou, A.; Davies, P.; Bruzaud, S. Natural Degradation and Biodegradation of Poly (3-Hydroxybutyrate-Co-3-Hydroxyvalerate) in Liquid and Solid Marine Environments. J. Polym. Environ. 2015, 23, 493–505. [Google Scholar] [CrossRef]
- Suzuki, S.; Ikada, Y. Medical Applications. In Poly(Lactic Acid); John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 581–604. ISBN 978-1-119-76748-0. [Google Scholar]
- Wang, G.-X.; Huang, D.; Ji, J.-H.; Völker, C.; Wurm, F.R. Seawater-Degradable Polymers—Fighting the Marine Plastic Pollution. Adv. Sci. 2021, 8, 2001121. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Tachibana, Y.; Kasuya, K. Biodegradability of Poly(3-Hydroxyalkanoate) and Poly(ε-Caprolactone) via Biological Carbon Cycles in Marine Environments. Polym. J. 2021, 53, 47–66. [Google Scholar] [CrossRef]
- Wright, R.J.; Erni-Cassola, G.; Zadjelovic, V.; Latva, M.; Christie-Oleza, J.A. Marine Plastic Debris: A New Surface for Microbial Colonization. Environ. Sci. Technol. 2020, 54, 11657–11672. [Google Scholar] [CrossRef]
- ter Halle, A.; Ghiglione, J.F. Nanoplastics: A Complex, Polluting Terra Incognita. Environ. Sci. Technol. 2021, 55, 14466–14469. [Google Scholar] [CrossRef]
- Latva, M.; Dedman, C.J.; Wright, R.J.; Polin, M.; Christie-Oleza, J.A. Microbial Pioneers of Plastic Colonisation in Coastal Seawaters. Mar. Pollut. Bull. 2022, 179, 113701. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, I.V.; Wichels, A.; Krohne, G.; Gerdts, G. Mature Biofilm Communities on Synthetic Polymers in Seawater-Specific or General? Mar. Environ. Res. 2018, 142, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Jacquin, J.; Conan, P.; Pujo-Pay, M.; Barbe, V.; George, M.; Fabre, P.; Bruzaud, S.; Ter Halle, A.; Meistertzheim, A.-L.; et al. Relative Influence of Plastic Debris Size and Shape, Chemical Composition and Phytoplankton-Bacteria Interactions in Driving Seawater Plastisphere Abundance, Diversity and Activity. Front. Microbiol. 2021, 11, 610231. [Google Scholar] [CrossRef] [PubMed]
- Pollet, T.; Berdjeb, L.; Garnier, C.; Durrieu, G.; Le Poupon, C.; Misson, B.; Briand, J.-F. Prokaryotic Community Successions and Interactions in Marine Biofilms: The Key Role of Flavobacteriia. FEMS Microbiol. Ecol. 2018, 94, fiy083. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2015, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.J.; Langille, M.G.I.; Walker, T.R. Food or Just a Free Ride? A Meta-Analysis Reveals the Global Diversity of the Plastisphere. ISME J. 2021, 15, 789–806. [Google Scholar] [CrossRef]
- Bryant, J.A.; Clemente, T.M.; Viviani, D.A.; Fong, A.A.; Thomas, K.A.; Kemp, P.; Karl, D.M.; White, A.E.; DeLong, E.F. Diversity and Activity of Communities Inhabiting Plastic Debris in the North Pacific Gyre. mSystems 2016, 1. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Osborn, A.M.; Duhaime, M.B. Microbes on a Bottle: Substrate, Season and Geography Influence Community Composition of Microbes Colonizing Marine Plastic Debris. PLoS ONE 2016, 11, e0159289. [Google Scholar] [CrossRef]
- Debroas, D.; Domaizon, I.; Humbert, J.-F.; Jardillier, L.; Lepère, C.; Oudart, A.; Taïb, N. Overview of Freshwater Microbial Eukaryotes Diversity: A First Analysis of Publicly Available Metabarcoding Data. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Ballerini, T.; Zettler, E.R.; Asbun, A.A.; Adame, A.; Casotti, R.; Dumontet, B.; Donnarumma, V.; Engelmann, J.C.; Frère, L.; et al. Diversity and Predicted Inter- and Intra-Domain Interactions in the Mediterranean Plastisphere. Environ. Pollut. 2021, 286, 117439. [Google Scholar] [CrossRef]
- Djaoudi, K.; Onrubia, J.A.T.; Boukra, A.; Guesnay, L.; Portas, A.; Barry-Martinet, R.; Angeletti, B.; Mounier, S.; Lenoble, V.; Briand, J.-F. Seawater Copper Content Controls Biofilm Bioaccumulation and Microbial Community on Microplastics. Sci. Total Environ. 2022, 814, 152278. [Google Scholar] [CrossRef] [PubMed]
- Kettner, M.T.; Rojas-Jimenez, K.; Oberbeckmann, S.; Labrenz, M.; Grossart, H.-P. Microplastics Alter Composition of Fungal Communities in Aquatic Ecosystems: Fungal Communities on Microplastics. Environ. Microbiol. 2017, 19, 4447–4459. [Google Scholar] [CrossRef] [PubMed]
- De Tender, C.; Devriese, L.I.; Haegeman, A.; Maes, S.; Vangeyte, J.; Cattrijsse, A.; Dawyndt, P.; Ruttink, T. Temporal Dynamics of Bacterial and Fungal Colonization on Plastic Debris in the North Sea. Environ. Sci. Technol. 2017, 51, 7350–7360. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, A.L.d.F.; Proietti, M.C.; Secchi, E.R.; Taylor, J.D. Diverse Groups of Fungi Are Associated with Plastics in the Surface Waters of the Western South Atlantic and the Antarctic Peninsula. Mol. Ecol. 2020, 29, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, J.; Liu, X.; Wang, L. Differences in the Plastispheres of Biodegradable and Non-Biodegradable Plastics: A Mini Review. Front. Microbiol. 2022, 13, 849147. [Google Scholar] [CrossRef] [PubMed]
- Dussud, C.; Meistertzheim, A.L.; Conan, P.; Pujo-Pay, M.; George, M.; Fabre, P.; Coudane, J.; Higgs, P.; Elineau, A.; Pedrotti, M.L.; et al. Evidence of Niche Partitioning among Bacteria Living on Plastics, Organic Particles and Surrounding Seawaters. Environ. Pollut. 2018, 236, 807–816. [Google Scholar] [CrossRef]
- Florio Furno, M.; Poli, A.; Ferrero, D.; Tardelli, F.; Manzini, C.; Oliva, M.; Pretti, C.; Campani, T.; Casini, S.; Fossi, M.C.; et al. The Culturable Mycobiota of Sediments and Associated Microplastics: From a Harbor to a Marine Protected Area, a Comparative Study. J. Fungi 2022, 8, 927. [Google Scholar] [CrossRef]
- Paço, A.; Duarte, K.; da Costa, J.P.; Santos, P.S.M.; Pereira, R.; Pereira, M.E.; Freitas, A.C.; Duarte, A.C.; Rocha-Santos, T.A.P. Biodegradation of Polyethylene Microplastics by the Marine Fungus Zalerion Maritimum. Sci. Total Environ. 2017, 586, 10–15. [Google Scholar] [CrossRef]
- Gao, R.; Liu, R.; Sun, C. A Marine Fungus Alternaria Alternata FB1 Efficiently Degrades Polyethylene. J. Hazard. Mater. 2022, 431, 128617. [Google Scholar] [CrossRef]
- Catao, E.C.P.; Gallois, N.; Fay, F.; Misson, B.; Briand, J.-F. Metal Resistance Genes Enrichment in Marine Biofilm Communities Selected by Biocide-Containing Surfaces in Temperate and Tropical Coastal Environments. Environ. Pollut. 2021, 268, 115835. [Google Scholar] [CrossRef]
- Lemonnier, C.; Chalopin, M.; Huvet, A.; Le Roux, F.; Labreuche, Y.; Petton, B.; Maignien, L.; Paul-Pont, I.; Reveillaud, J. Time-Series Incubations in a Coastal Environment Illuminates the Importance of Early Colonizers and the Complexity of Bacterial Biofilm Dynamics on Marine Plastics. Environ. Pollut. 2022, 312, 119994. [Google Scholar] [CrossRef] [PubMed]
- Odobel, C.; Dussud, C.; Philip, L.; Derippe, G.; Lauters, M.; Eyheraguibel, B.; Burgaud, G.; Ter Halle, A.; Meistertzheim, A.-L.; Bruzaud, S.; et al. Bacterial Abundance, Diversity and Activity During Long-Term Colonization of Non-Biodegradable and Biodegradable Plastics in Seawater. Front. Microbiol. 2021, 12, 734782. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Walters, W.A.; Lennon, N.J.; Bochicchio, J.; Krohn, A.; Caporaso, J.G.; Pennanen, T. Accurate Estimation of Fungal Diversity and Abundance through Improved Lineage-Specific Primers Optimized for Illumina Amplicon Sequencing. Appl. Environ. Microbiol. 2016, 82, 7217–7226. [Google Scholar] [CrossRef]
- Borneman, J.; Hartin, R.J. PCR Primers That Amplify Fungal RRNA Genes from Environmental Samples. Appl. Environ. Microbiol. 2000, 66, 4356–4360. [Google Scholar] [CrossRef] [PubMed]
- Pachiadaki, M.G.; Rédou, V.; Beaudoin, D.J.; Burgaud, G.; Edgcomb, V.P. Fungal and Prokaryotic Activities in the Marine Subsurface Biosphere at Peru Margin and Canterbury Basin Inferred from RNA-Based Analyses and Microscopy. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Noel, C.; Cormier, A.; Durand, P.; Leroi, L. SAMBA: Standardized and Automated MetaBarcoding Analyses Workflow. 2022. WorkflowHub. Available online: https://workflowhub.eu/workflows/156 (accessed on 4 March 2023).
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow Enables Reproducible Computational Workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Olesen, S.W.; Duvallet, C.; Alm, E.J. DbOTU3: A New Implementation of Distribution-Based OTU Calling. PLoS ONE 2017, 12, e0176335. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE Database for Molecular Identification of Fungi: Handling Dark Taxa and Parallel Taxonomic Classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Frère, L.; Maignien, L.; Chalopin, M.; Huvet, A.; Rinnert, E.; Morrison, H.; Kerninon, S.; Cassone, A.-L.; Lambert, C.; Reveillaud, J.; et al. Microplastic Bacterial Communities in the Bay of Brest: Influence of Polymer Type and Size. Environ. Pollut. Barking Essex 1987 2018, 242, 614–625. [Google Scholar] [CrossRef]
- Kettner, M.T.; Oberbeckmann, S.; Labrenz, M.; Grossart, H.-P. The Eukaryotic Life on Microplastics in Brackish Ecosystems. Front. Microbiol. 2019, 10, 538. [Google Scholar] [CrossRef]
- Yang, L.-Y.; Huang, X.-R.; Neilson, R.; Zhou, S.-Y.-D.; Li, Z.-L.; Yang, X.-R.; Su, X.-X. Characterization of Microbial Community, Ecological Functions and Antibiotic Resistance in Estuarine Plastisphere. Sci. Total Environ. 2023, 866, 161322. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, W.; Zhang, Z.; Grossart, H.-P.; Gadd, G.M. Microplastics Provide New Microbial Niches in Aquatic Environments. Appl. Microbiol. Biotechnol. 2020, 104, 6501–6511. [Google Scholar] [CrossRef]
- Tetu, S.G.; Sarker, I.; Schrameyer, V.; Pickford, R.; Elbourne, L.D.H.; Moore, L.R.; Paulsen, I.T. Plastic Leachates Impair Growth and Oxygen Production in Prochlorococcus, the Ocean’s Most Abundant Photosynthetic Bacteria. Commun. Biol. 2019, 2, 1–9. [Google Scholar] [CrossRef]
- Wallbank, J.A.; Lear, G.; Kingsbury, J.M.; Weaver, L.; Doake, F.; Smith, D.A.; Audrézet, F.; Maday, S.D.M.; Gambarini, V.; Donaldson, L.; et al. Into the Plastisphere, Where Only the Generalists Thrive: Early Insights in Plastisphere Microbial Community Succession. Front. Mar. Sci. 2022, 9, 841142. [Google Scholar] [CrossRef]
- Gerphagnon, M.; Colombet, J.; Latour, D.; Sime-Ngando, T. Spatial and Temporal Changes of Parasitic Chytrids of Cyanobacteria. Sci. Rep. 2017, 7, 6056. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; de Bruin, A.; Ibelings, B.W.; Van Donk, E. Parasitic Chytrids: Their Effects on Phytoplankton Communities and Food-Web Dynamics. Hydrobiologia 2007, 578, 113–129. [Google Scholar] [CrossRef]
- Lacerda, A.L.d.F.; Rodrigues, L. dos S.; van Sebille, E.; Rodrigues, F.L.; Ribeiro, L.; Secchi, E.R.; Kessler, F.; Proietti, M.C. Plastics in Sea Surface Waters around the Antarctic Peninsula. Sci. Rep. 2019, 9, 3977. [Google Scholar] [CrossRef]
- Srikanth, M.; Sandeep, T.S.R.S.; Sucharitha, K.; Godi, S. Biodegradation of Plastic Polymers by Fungi: A Brief Review. Bioresour. Bioprocess. 2022, 9, 42. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Environmental Factors Support the Formation of Specific Bacterial Assemblages on Microplastics. Front. Microbiol. 2018, 8, 2709. [Google Scholar] [CrossRef]
- Pinto, M.; Langer, T.M.; Hüffer, T.; Hofmann, T.; Herndl, G.J. The Composition of Bacterial Communities Associated with Plastic Biofilms Differs between Different Polymers and Stages of Biofilm Succession. PLoS ONE 2019, 14, e0217165. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tong, J.; Li, Y.; Zhu, J.; Zhang, W.; Niu, L.; Zhang, H. Bacterial and Fungal Assemblages and Functions Associated with Biofilms Differ between Diverse Types of Plastic Debris in a Freshwater System. Environ. Res. 2021, 196, 110371. [Google Scholar] [CrossRef] [PubMed]
- Dussud, C.; Hudec, C.; George, M.; Fabre, P.; Higgs, P.; Bruzaud, S.; Delort, A.-M.; Eyheraguibel, B.; Meistertzheim, A.-L.; Jacquin, J.; et al. Colonization of Non-Biodegradable and Biodegradable Plastics by Marine Microorganisms. Front. Microbiol. 2018, 9, 1571. [Google Scholar] [CrossRef]
- Eich, A.; Mildenberger, T.; Laforsch, C.; Weber, M. Biofilm and Diatom Succession on Polyethylene (PE) and Biodegradable Plastic Bags in Two Marine Habitats: Early Signs of Degradation in the Pelagic and Benthic Zone? PLoS ONE 2015, 10, e0137201. [Google Scholar] [CrossRef]
- Miao, L.; Li, W.; Adyel, T.M.; Yao, Y.; Deng, Y.; Wu, J.; Zhou, Y.; Yu, Y.; Hou, J. Spatio-Temporal Succession of Microbial Communities in Plastisphere and Their Potentials for Plastic Degradation in Freshwater Ecosystems. Water Res. 2023, 229, 119406. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Campos, S.; González-Pleiter, M.; Fernández-Piñas, F.; Rosal, R.; Leganés, F. Early and Differential Bacterial Colonization on Microplastics Deployed into the Effluents of Wastewater Treatment Plants. Sci. Total Environ. 2021, 757, 143832. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Jia, W.; Xu, L.; Zhang, M.; Huang, Y. The Plastisphere of Biodegradable and Conventional Microplastics from Residues Exhibit Distinct Microbial Structure, Network and Function in Plastic-Mulching Farmland. J. Hazard. Mater. 2023, 442, 130011. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, S.; Liquet y González, J.E.; Henderson, K.B.; Anunciado, M.B.; Hayes, D.G.; DeBruyn, J.M. Soil Microbial Communities Associated with Biodegradable Plastic Mulch Films. Front. Microbiol. 2020, 11, 587074. [Google Scholar] [CrossRef] [PubMed]
- Mercier, A.; Gravouil, K.; Aucher, W.; Brosset-Vincent, S.; Kadri, L.; Colas, J.; Bouchon, D.; Ferreira, T. Fate of Eight Different Polymers under Uncontrolled Composting Conditions: Relationships Between Deterioration, Biofilm Formation, and the Material Surface Properties. Environ. Sci. Technol. 2017, 51, 1988–1997. [Google Scholar] [CrossRef]
- Spadaro, D.; Bustos-Lopez, M.p.; Gullino, M.l.; Piano, S.; Tabacco, E.; Borreani, G. Evolution of Fungal Populations in Corn Silage Conserved under Polyethylene or Biodegradable Films. J. Appl. Microbiol. 2015, 119, 510–520. [Google Scholar] [CrossRef]
- Jeyakumar, D.; Chirsteen, J.; Doble, M. Synergistic Effects of Pretreatment and Blending on Fungi Mediated Biodegradation of Polypropylenes. Bioresour. Technol. 2013, 148, 78–85. [Google Scholar] [CrossRef]
- Harrison, J.P.; Schratzberger, M.; Sapp, M.; Osborn, A.M. Rapid Bacterial Colonization of Low-Density Polyethylene Microplastics in Coastal Sediment Microcosms. BMC Microbiol. 2014, 14, 232. [Google Scholar] [CrossRef]
- De Tender, C.A.; Devriese, L.I.; Haegeman, A.; Maes, S.; Ruttink, T.; Dawyndt, P. Bacterial Community Profiling of Plastic Litter in the Belgian Part of the North Sea. Environ. Sci. Technol. 2015, 49, 9629–9638. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Loeder, M.G.J.; Gerdts, G.; Osborn, A.M. Spatial and Seasonal Variation in Diversity and Structure of Microbial Biofilms on Marine Plastics in Northern European Waters. FEMS Microbiol. Ecol. 2014, 90, 478–492. [Google Scholar] [CrossRef]
- Basili, M.; Quero, G.M.; Giovannelli, D.; Manini, E.; Vignaroli, C.; Avio, C.G.; De Marco, R.; Luna, G.M. Major Role of Surrounding Environment in Shaping Biofilm Community Composition on Marine Plastic Debris. Front. Mar. Sci. 2020, 7, 262. [Google Scholar] [CrossRef]
- Panebianco, C. Temperature Requirements of Selected Marine Fungi. Bot. Mar. 1994, 37, 157–162. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Ramakrishna, S.; Vikineswary, S.; Das, D.; Bahkali, A.H.; Guo, S.-Y.; Pang, K.-L. How Do Fungi Survive in the Sea and Respond to Climate Change? J. Fungi 2022, 8, 291. [Google Scholar] [CrossRef] [PubMed]
- Oberbeckmann, S.; Labrenz, M. Marine Microbial Assemblages on Microplastics: Diversity, Adaptation, and Role in Degradation. Annu. Rev. Mar. Sci. 2020, 12, 209–232. [Google Scholar] [CrossRef]
- Caruso, G. Microbial Colonization in Marine Environments: Overview of Current Knowledge and Emerging Research Topics. J. Mar. Sci. Eng. 2020, 8, 78. [Google Scholar] [CrossRef]
- Jacquin, J.; Cheng, J.; Odobel, C.; Pandin, C.; Conan, P.; Pujo-Pay, M.; Barbe, V.; Meistertzheim, A.-L.; Ghiglione, J.-F. Microbial Ecotoxicology of Marine Plastic Debris: A Review on Colonization and Biodegradation by the “Plastisphere”. Front. Microbiol. 2019, 10, 865. [Google Scholar] [CrossRef]
- Miri, S.; Saini, R.; Davoodi, S.M.; Pulicharla, R.; Brar, S.K.; Magdouli, S. Biodegradation of Microplastics: Better Late than Never. Chemosphere 2022, 286, 131670. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Philippe, A.; Noël, C.; Eyheraguibel, B.; Briand, J.-F.; Paul-Pont, I.; Ghiglione, J.-F.; Coton, E.; Burgaud, G. Fungal Diversity and Dynamics during Long-Term Immersion of Conventional and Biodegradable Plastics in the Marine Environment. Diversity 2023, 15, 579. https://doi.org/10.3390/d15040579
Philippe A, Noël C, Eyheraguibel B, Briand J-F, Paul-Pont I, Ghiglione J-F, Coton E, Burgaud G. Fungal Diversity and Dynamics during Long-Term Immersion of Conventional and Biodegradable Plastics in the Marine Environment. Diversity. 2023; 15(4):579. https://doi.org/10.3390/d15040579
Chicago/Turabian StylePhilippe, Aurélie, Cyril Noël, Boris Eyheraguibel, Jean-François Briand, Ika Paul-Pont, Jean-François Ghiglione, Emmanuel Coton, and Gaëtan Burgaud. 2023. "Fungal Diversity and Dynamics during Long-Term Immersion of Conventional and Biodegradable Plastics in the Marine Environment" Diversity 15, no. 4: 579. https://doi.org/10.3390/d15040579
APA StylePhilippe, A., Noël, C., Eyheraguibel, B., Briand, J.-F., Paul-Pont, I., Ghiglione, J.-F., Coton, E., & Burgaud, G. (2023). Fungal Diversity and Dynamics during Long-Term Immersion of Conventional and Biodegradable Plastics in the Marine Environment. Diversity, 15(4), 579. https://doi.org/10.3390/d15040579