


Appraisal of the Genus Pleurastrum (Chlorophyta) Based on Molecular and Climate Data


Abstract
:1. Introduction
2. Materials and Methods
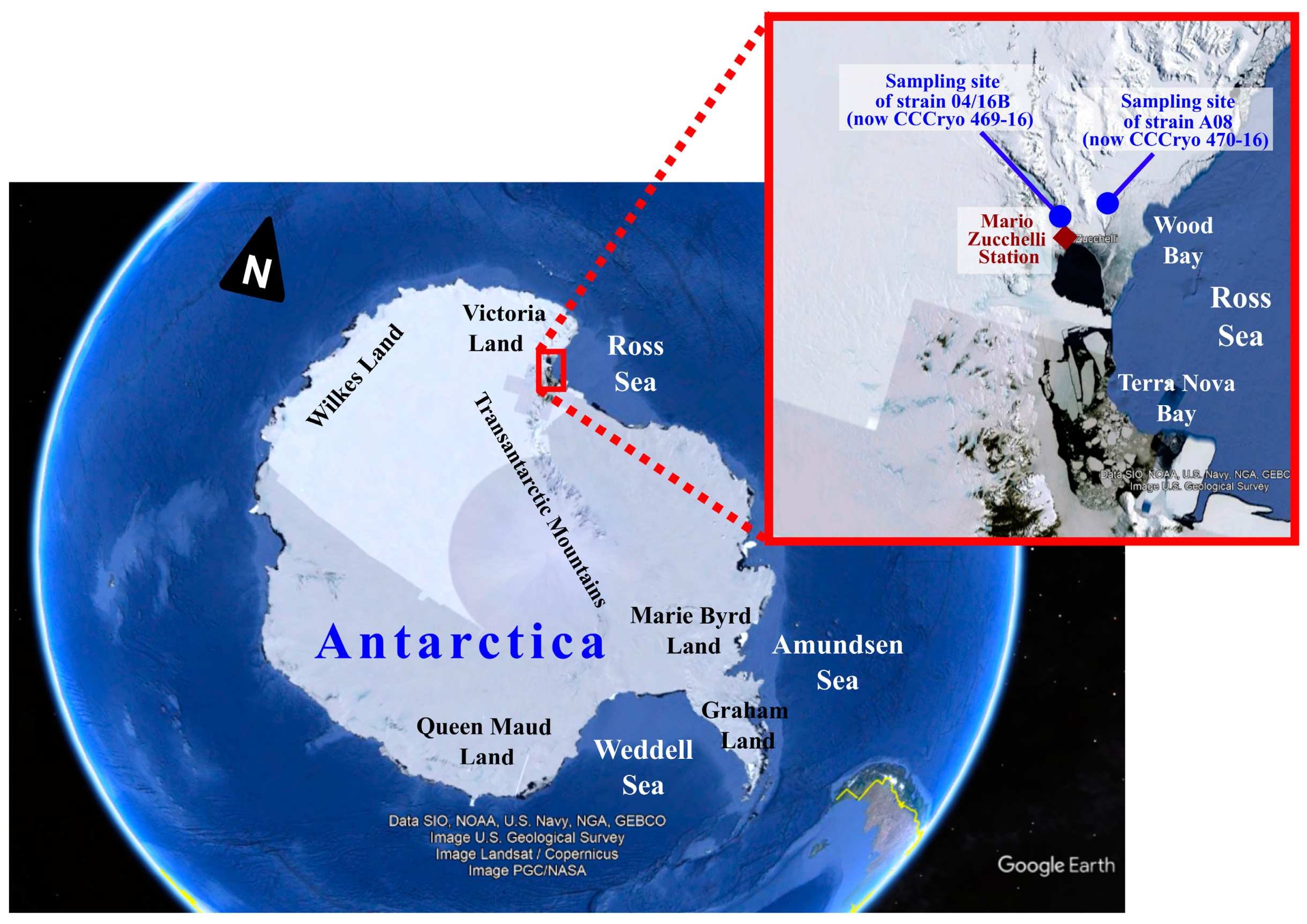
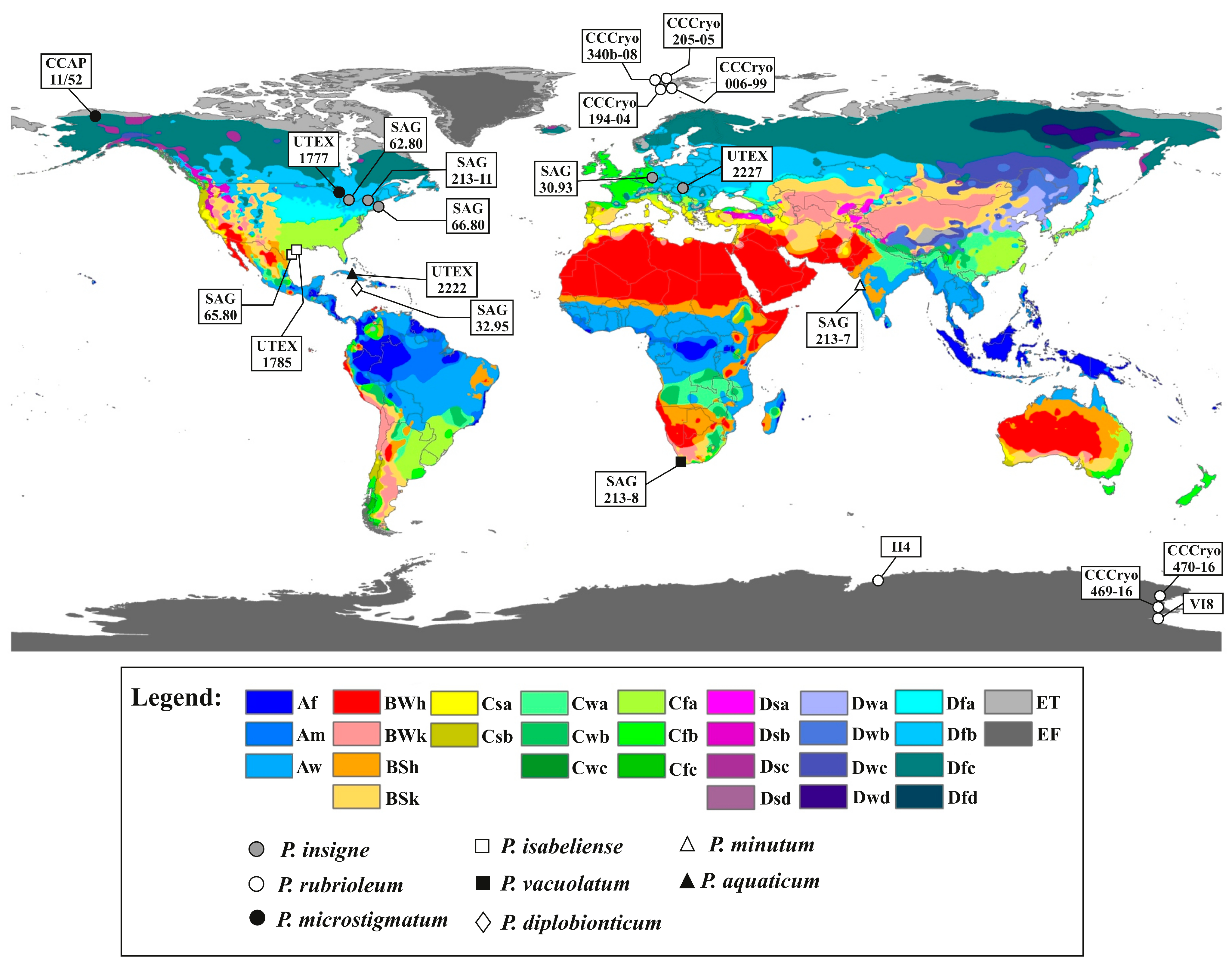
2.1. Sampling and Culture Set up of Antarctic Microalgal Strains
2.2. Taxon Selection, Culture Conditions, and Light Microscope Observations
2.3. Electron Microscope Observations
2.4. DNA Extraction, Amplification, and Sequencing
2.5. Molecular and Phylogenetic Analyses
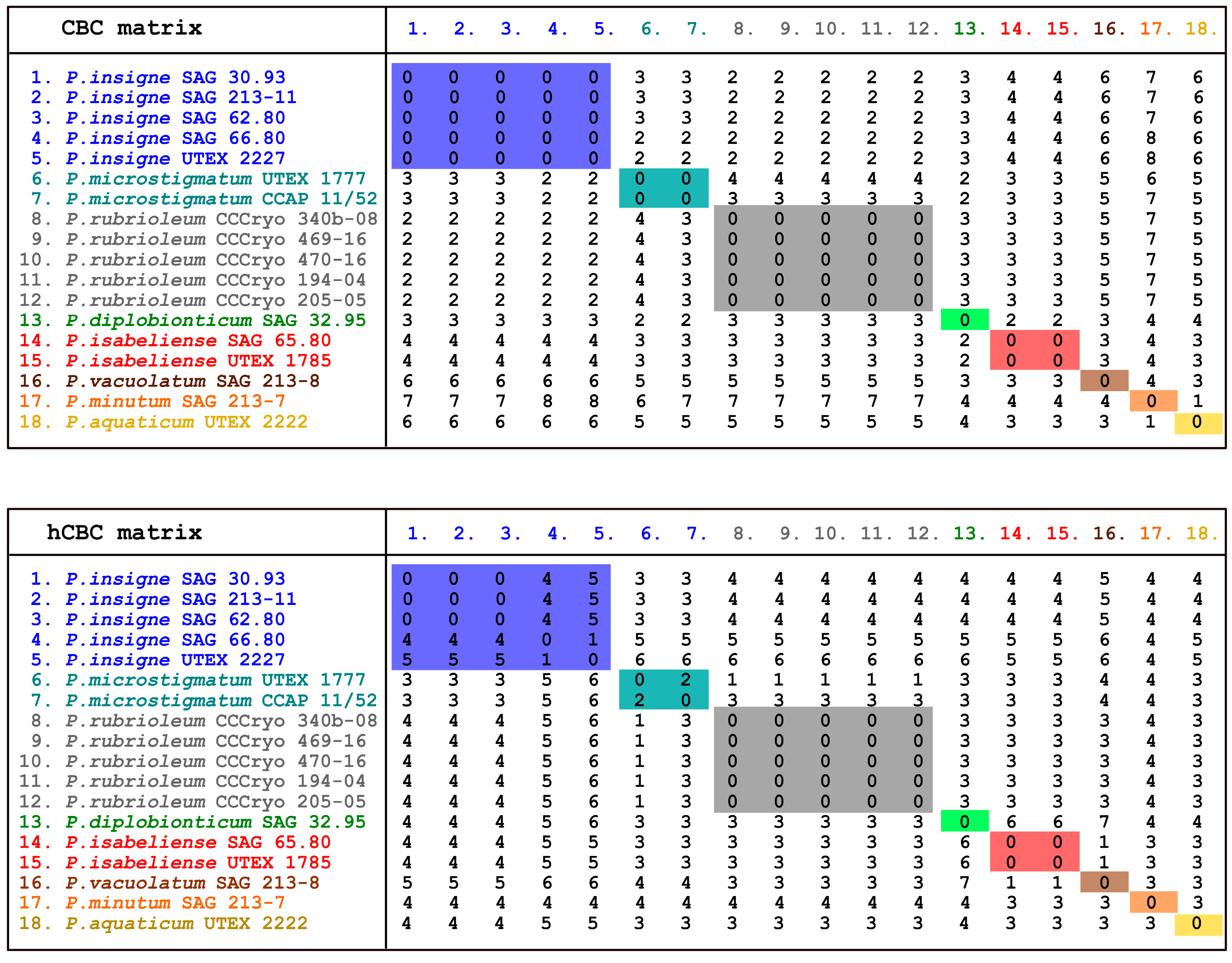
2.6. ITS2 Secondary Structure Predictions and Analyses
3. Results
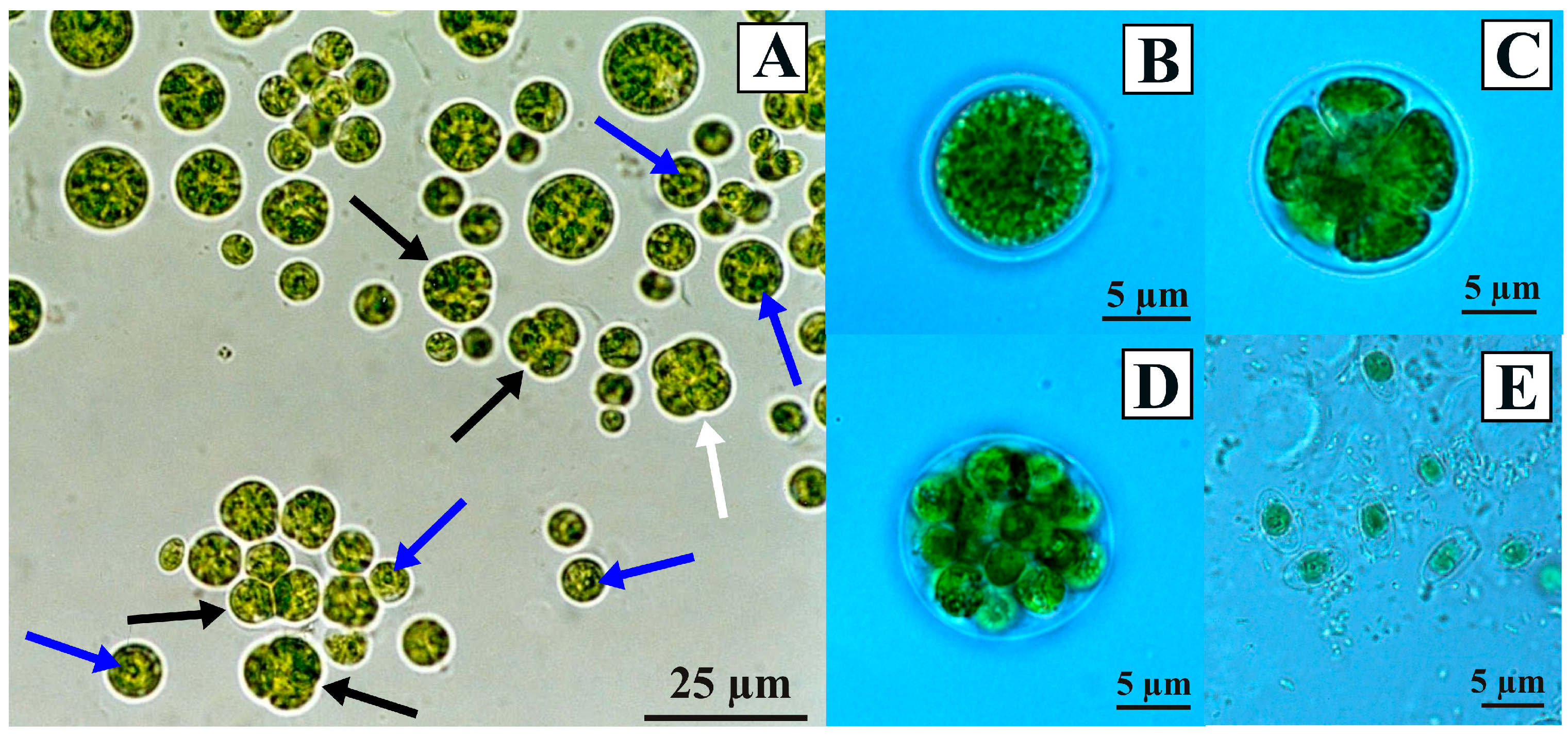
3.1. Morphological Observations
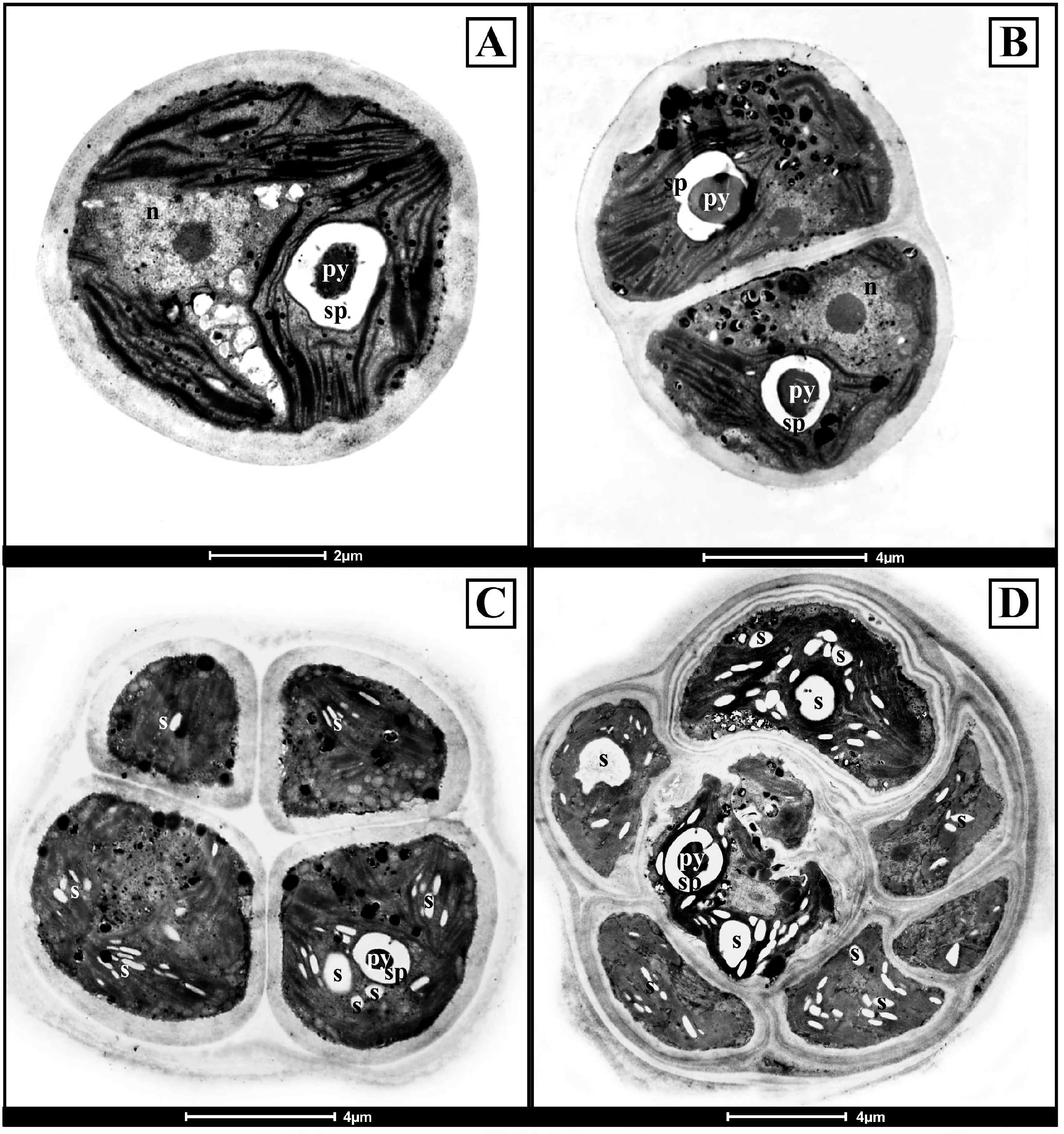
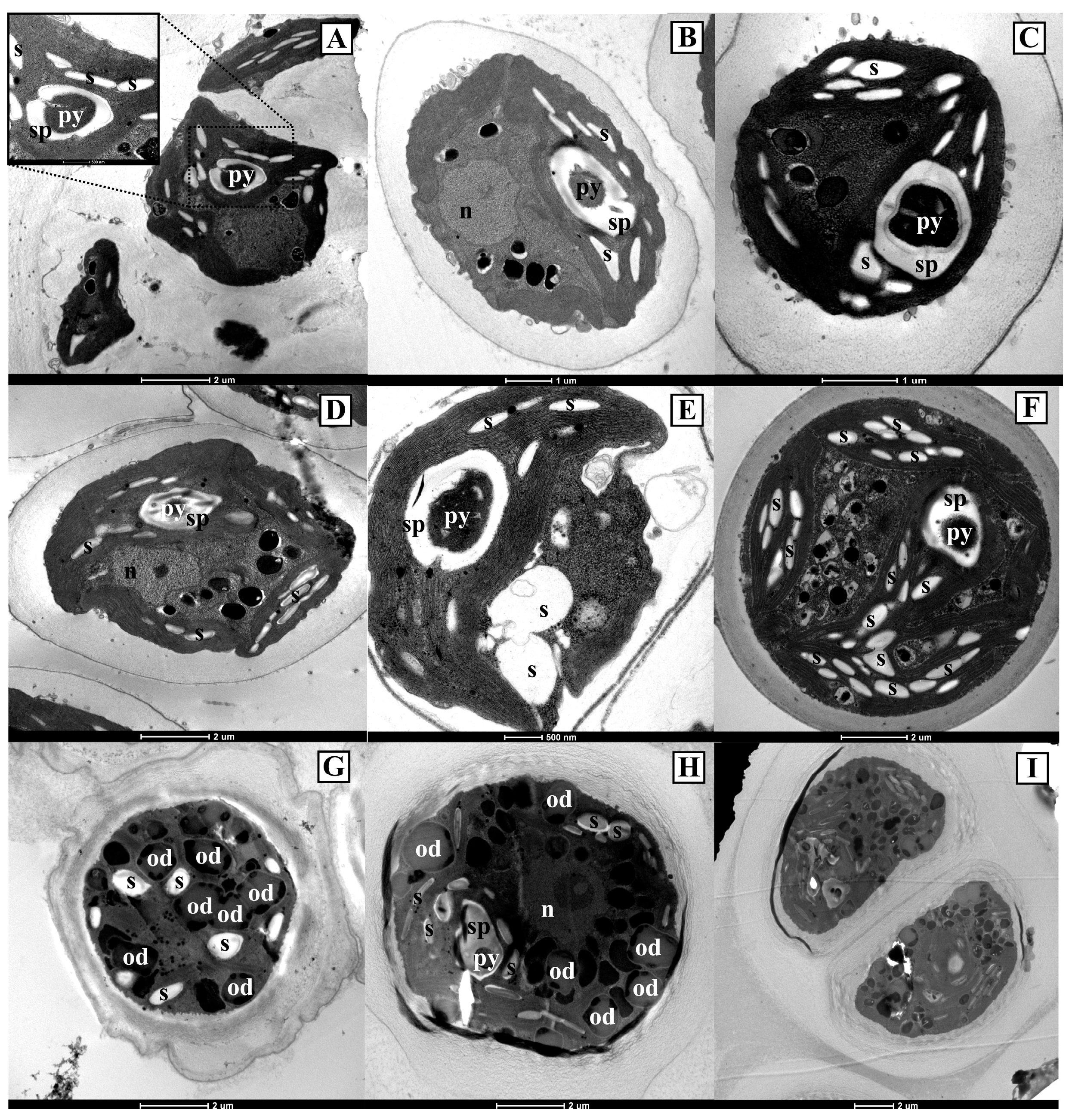
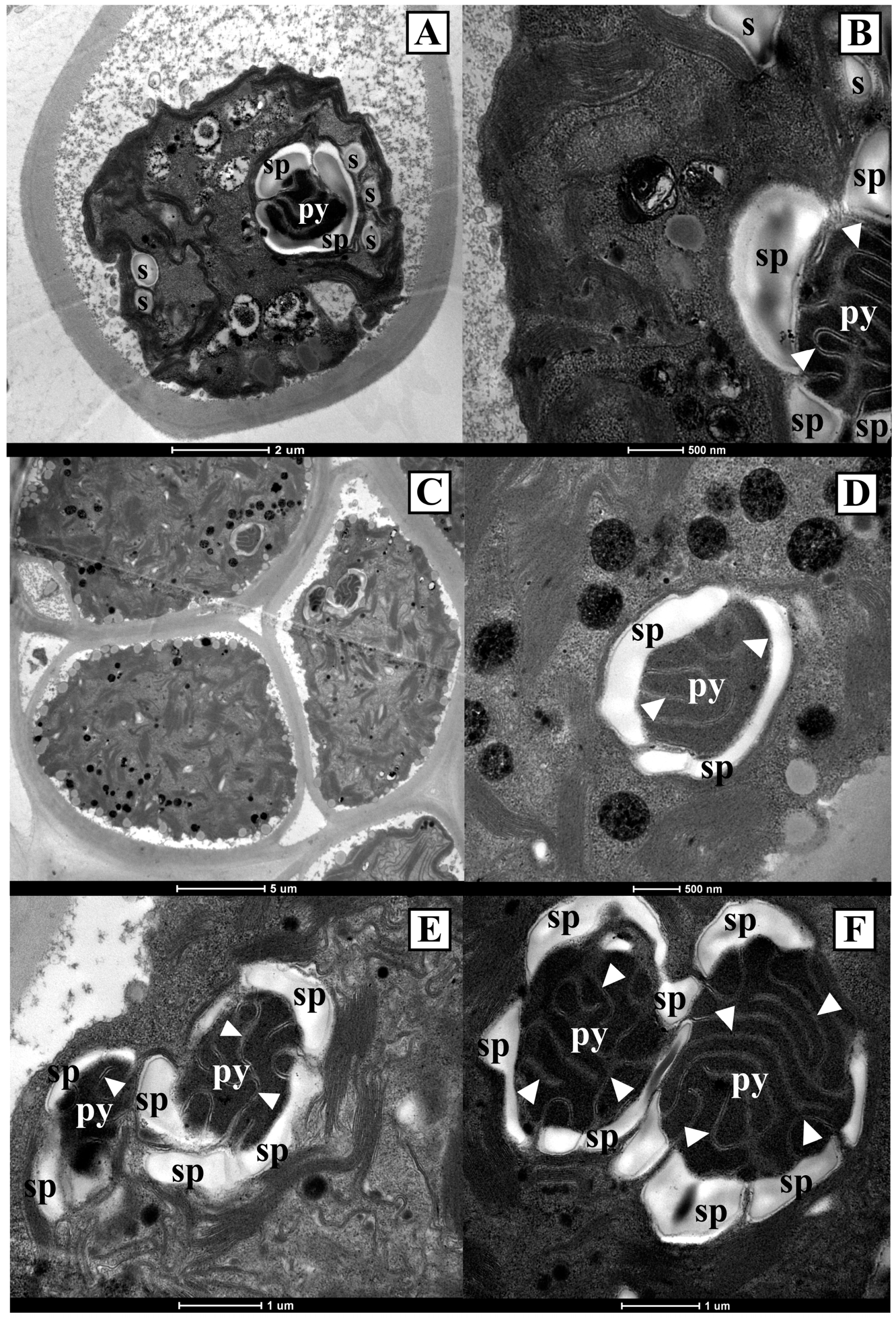
3.2. Ultrastructural Observations on the Antarctic Isolates
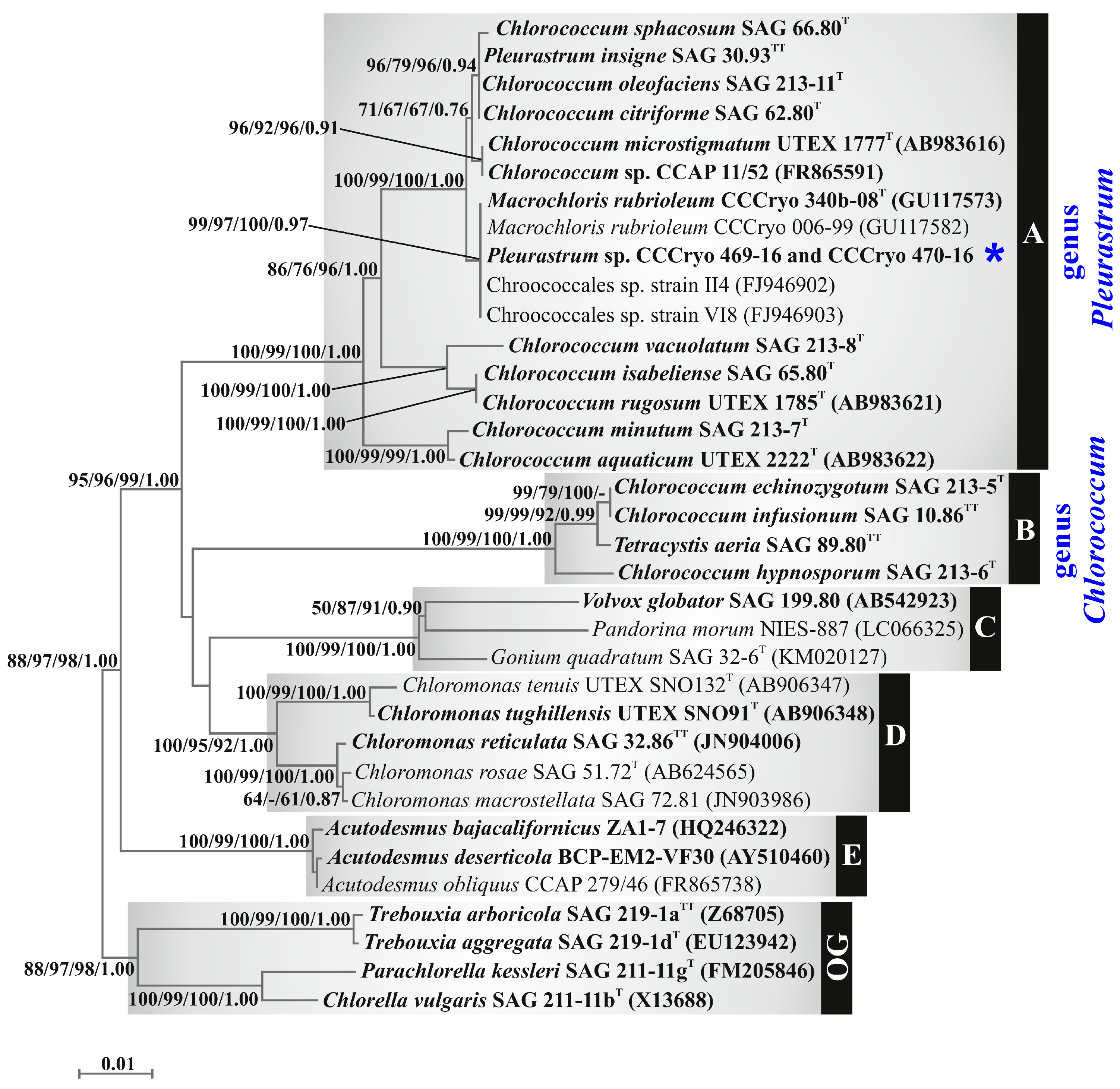
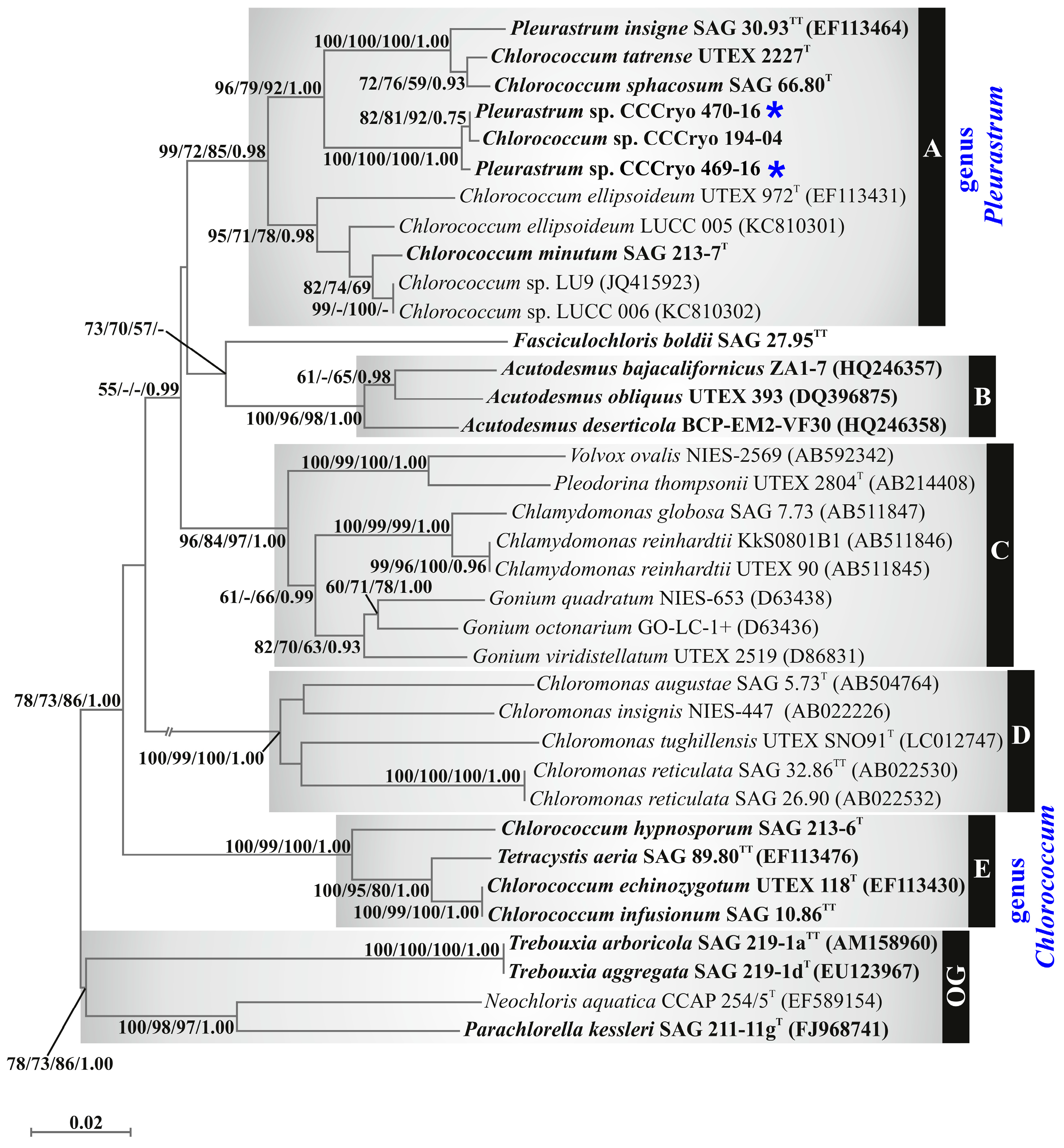
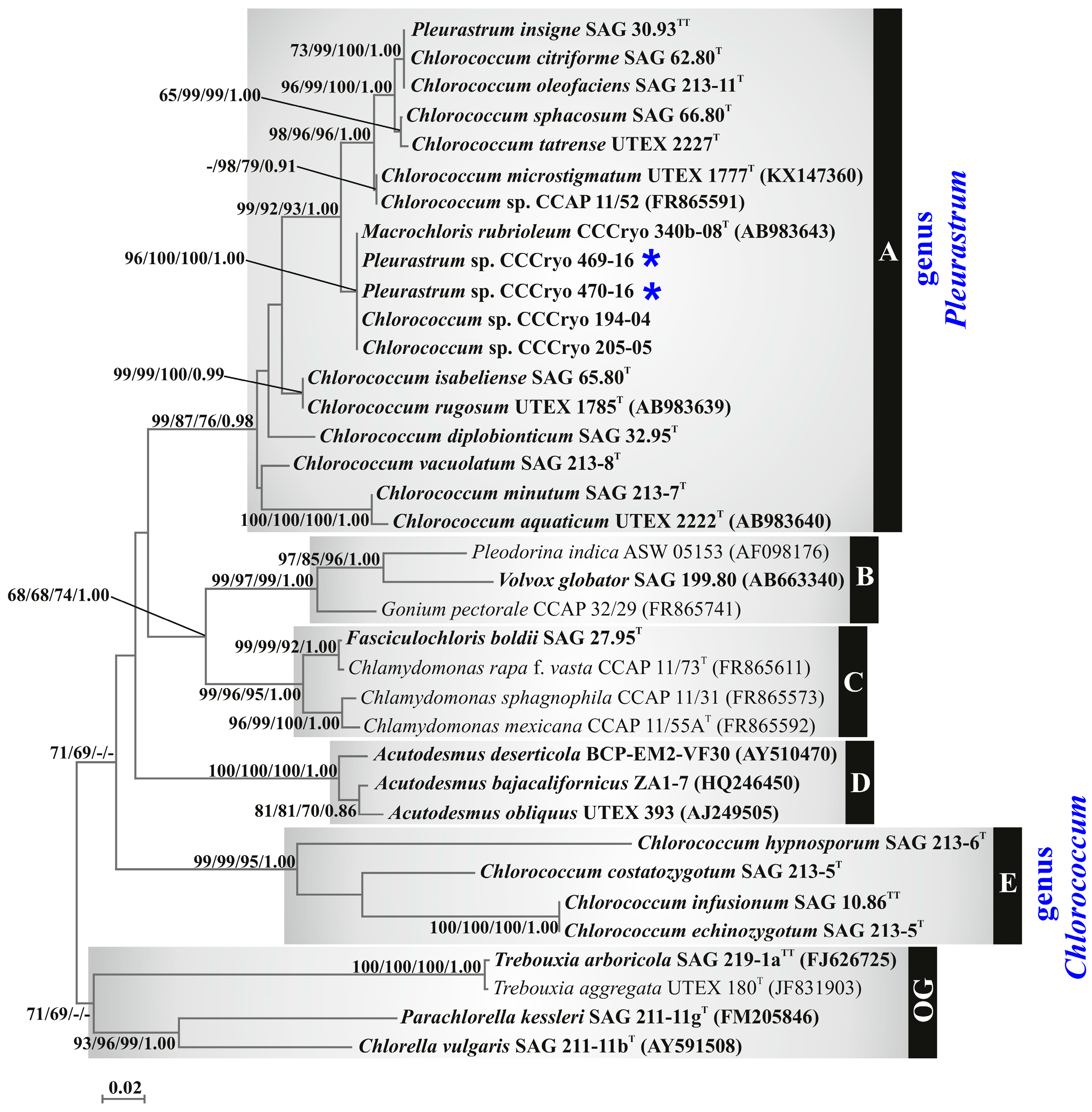
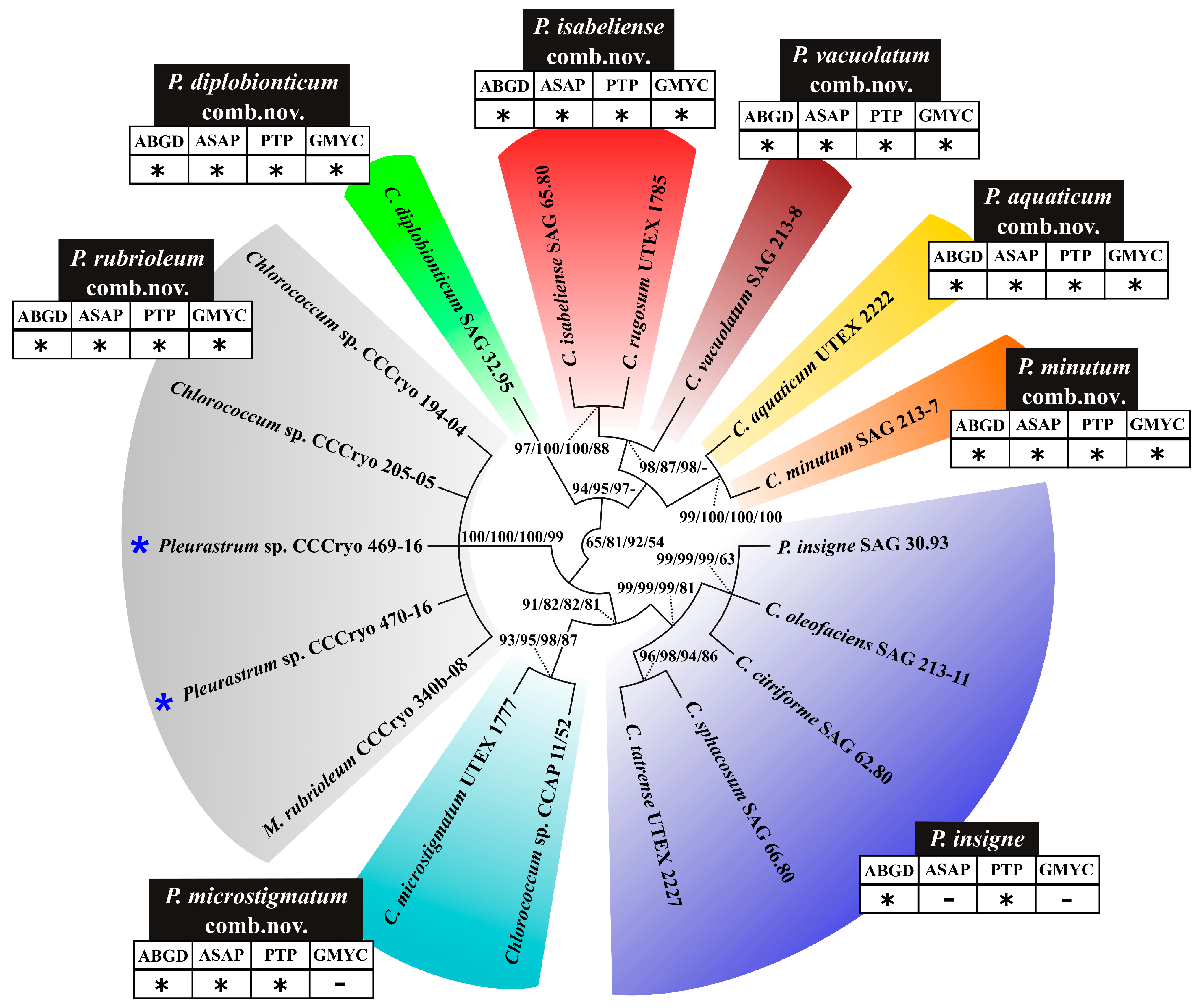
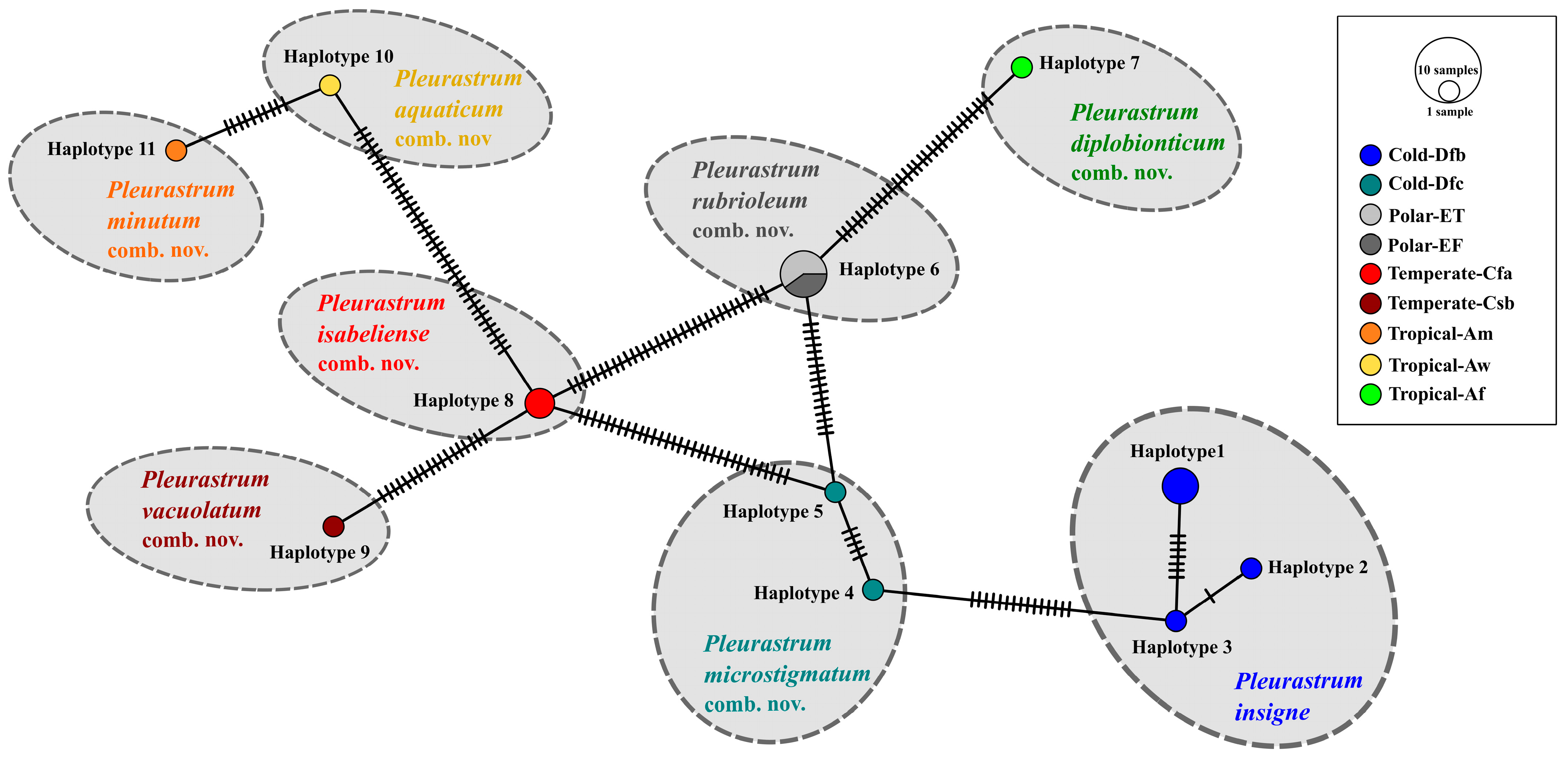
3.3. Molecular and Phylogenetic Results
3.4. Ultrastructural Observations on the Strains Obtained from International Culture Collections
3.5. ITS2 Secondary Structure Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andersen, R.A. Diversity of eukaryotic algae. Biodiver. Conserv. 1992, 10, 267–292. [Google Scholar] [CrossRef]
- Malcata, F.X.; Sousa Pinto, I.; Guedes, A.C. Marine Macro- and Microalgae: An Overview, 1st ed.; CRC Press: Boca Raton, FL, USA, 2018; p. 342. [Google Scholar] [CrossRef]
- Komárek, J.; Nedbalová, L. Green Cryosestic Algae. In Algae and Cyanobacteria in Extreme Environments, 1st ed.; Seckbach, J., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 321–342. [Google Scholar] [CrossRef]
- Leya, T.; Rahn, A.; Lütz, C.; Remias, D. Response of arctic snow and permafrost algae to high light and nitrogen stress by changes in pigment composition and applied aspects for biotechnology. FEMS Microbio. Ecol. 2009, 67, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Remias, D.; Karsten, U.; Lütz, C.; Leya, T. Physiological and morphological processes in the Alpine snow alga Chloromonas nivalis (Chlorophyceae) during cyst formation. Protoplasma 2010, 243, 73–86. [Google Scholar] [CrossRef]
- Remias, D.; Wastian, H.; Lütz, C.; Leya, T. Insights into the biology and phylogeny of Chloromonas polyptera (Chlorophyta), an alga causing orange snow in Maritime Antarctica. Antarct. Sci. 2013, 25, 648–656. [Google Scholar] [CrossRef]
- Procházková, L.; Leya, T.; Křížková, H.; Nedbalová, L. Sanguina nivaloides and Sanguina aurantia gen. et spp. nov. (Chlorophyta): The taxonomy, phylogeny, biogeography and ecology of two newly recognised algae causing red and orange snow. FEMS Microbiol. Ecol. 2019, 95, fiz064. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes, and antibacterial activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Levasseur, W.; Perré, P.; Pozzobon, V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol. Adv. 2020, 41, 107545. [Google Scholar] [CrossRef]
- Fernandes, T.; Cordeiro, N. Microalgae as Sustainable Biofactories to Produce High-Value Lipids: Biodiversity, Exploitation, and Biotechnological Applications. Mar. Drugs 2021, 19, 573. [Google Scholar] [CrossRef]
- Leya, T. The CCCryo Culture Collection of Cryophilic Algae as a valuable bioresource for algal biodiversity and for novel, industrially marketable metabolites. Appl. Phycol. 2020, 3, 167–188. [Google Scholar] [CrossRef]
- Andreoli, C.; Lokhorst, G.M.; Mani, A.M.; Scarabel, L.; Moro, I.; La Rocca, N.; Tognetto, L. Koliella antarctica sp. nov. (Klebsormidiales) a new marine green microalga from the Ross Sea (Antarctica). Arch. Hydrobiol. Algol. Stud. 1998, 90, 1–8. [Google Scholar] [CrossRef]
- Sciuto, K.; Lewis, L.A.; Verleyen, E.; Moro, I.; La Rocca, N. Chodatodesmus australis sp. nov. (Scenedesmaceae, Chlorophyta) from Antarctica, with the emended description of the genus Chodatodesmus, and circumscription of Flechtneria rotunda gen. et sp. nov. J. Phycol. 2015, 51, 1172–1188. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, F.E.; Saldarriaga-Córdoba, M.; Huovinen, P.; Silva, A.X.; Gómez, I. Revealing the Characteristics of the Antarctic Snow Alga Chlorominima collina gen. et sp. nov. Through Taxonomy, Physiology, and Transcriptomics. Front. Plant Sci. 2021, 12, 662298. [Google Scholar] [CrossRef] [PubMed]
- Trentin, R.; Negrisolo, E.; Moschin, E.; Veronese, D.; Cecchetto, M.; Moro, I. Microglena antarctica sp. nov. a New Antarctic Green Alga from Inexpressible Island (Terra Nova Bay, Ross Sea) Revealed through an Integrative Approach. Diversity 2022, 14, 337. [Google Scholar] [CrossRef]
- Chodat, R. Materiaux pour servir a l’histoire des Protococcoidées. Bull. Herb. Boissier 1894, 2, 585–616. [Google Scholar]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Freshwater Flora of Central Europe. Vol. 13: Chlorophyta: Ulvophyceae, 1st ed.; Springer Spektrum: Berlin, Heidelberg, 2018; pp. 123–127. [Google Scholar] [CrossRef]
- Sluiman, H.J.; Gärtner, G. Taxonomic studies on the genus Pleurastrum (Pleurastrales, Chlorophyta). I. The type species, P. insigne, rediscovered and isolated from soil. Phycologia 1990, 29, 133–138. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2018; Available online: https://www.algaebase.org (accessed on 2 January 2023).
- Kawasaki, Y.; Nakada, T.; Tomita, M. Taxonomic revision of oil-producing green algae, Chlorococcum oleofaciens (Volvocales, Chlorophyceae), and its relatives. J. Phycol. 2015, 51, 1000–1016. [Google Scholar] [CrossRef]
- Aaronson, S. Experimental Microbial Ecology; Academic Press: New York, NY, USA, 1970; p. 236. [Google Scholar]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of Cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Andreoli, C.; Moro, I.; La Rocca, I.; Rigoni, F.; Dalla Valle, L.; Bargelloni, L. Pseudopleurochloris antarctica gen. et sp. nov., a new coccoid xanthophycean from pack-ice of Wood Bay (Ross Sea, Antarctica): Ultrastructure, pigments and 18S rRNA gene sequence. Eur. J. Phycol. 1999, 34, 149–159. [Google Scholar] [CrossRef]
- Famà, P.; Wysor, B.; Kooistra, W.; Zuccarello, G.C. Molecular phylogeny of the genus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufA gene. J. Phycol. 2002, 38, 1040–1050. [Google Scholar] [CrossRef]
- Hall, J.D.; Fucikova, K.; Lo, C.; Lewis, L.A.; Karol, K.G. An assessment of proposed DNA barcodes in freshwater green algae. Cryptogam. Algol. 2010, 31, 529–555. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of the AIC and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer, Version 1.5. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 2 December 2022).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 2.71. 2009. Available online: http://mesquiteproject.org (accessed on 5 December 2022).
- Schultz, J.; Müller, T.; Achtziger, M.; Seibel, P.N.; Dandekar, T.; Wolf, M. The internal transcribed spacer 2 database—A web server for (not only) low level phylogenetic analyses. Nucleic Acids Res. 2006, 34, W704–W707. [Google Scholar] [CrossRef] [PubMed]
- Selig, C.; Wolf, M.; Müller, T.; Dandekar, T.; Schultz, J. The ITS2 Database II: Homology modelling RNA structure for molecular systematics. Nucleic Acids Res. 2008, 36, D377–D380. [Google Scholar] [CrossRef] [PubMed]
- Koetschan, C.; Förster, F.; Keller, A.; Schleicher, T.; Ruderisch, B.; Schwarz, R.; Müller, T.; Wolf, M.; Schultz, J. The ITS2 database III—Sequences and structures for phylogeny. Nucleic Acids Res. 2010, 38, D275–D279. [Google Scholar] [CrossRef] [PubMed]
- Koetschan, C.; Hackl, T.; Müller, T.; Wolf, M.; Förster, F.; Schultz, J. ITS2 database IV: Interactive taxon sampling for internal transcribed spacer 2 based phylogenies. Mol. Phylogenet. Evol. 2012, 63, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Schultz, J.; Wolf, M. 4SALE-A tool for synchronous RNA sequence and secondary structure alignment and editing. BMC Bioinform. 2006, 7, 498. [Google Scholar] [CrossRef]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Wolf, M. Synchronous visual analysis and editing of RNA sequence and secondary structure alignments using 4SALE. BMC Res. Notes 2008, 1, 91. [Google Scholar] [CrossRef] [PubMed]
- Siebert, S.; Backofen, R. MARNA: Multiple alignment and consensus structure prediction of RNAs based on sequence structure comparisons. Bioinformatics 2005, 21, 3352–3359. [Google Scholar] [CrossRef]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [CrossRef]
- Wolf, M.; Friedrich, J.; Dandekar, T.; Müller, T. CBCAnalyzer: Inferring phylogenies based on compensatory base changes in RNA secondary structures. In Silico Biol. 2005, 5, 291–294. [Google Scholar]
- Wolf, M.; Ruderisch, B.; Dandekar, T.; Müller, T. ProfdistS: (Profile-) distance based phylogeny on sequence-structure alignments. Bioinformatics 2008, 24, 2401–2402. [Google Scholar] [CrossRef]
- Müller, T.; Rahmann, S.; Dandekar, T.; Wolf, M. Accurate and robust phylogeny estimation based on profile distances: A study of the Chlorophyceae (Chlorophyta). BMC Evol. Biol. 2004, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Darienko, T.; Gustavs, L.; Eggert, A.; Wolf, W.; Pröschold, T. Evaluating the species boundaries of green microalgae (Coccomyxa, Trebouxiophyceae, Chlorophyta) using integrative taxonomy and DNA barcoding with further implications for the species identification in environmental samples. PLoS ONE 2015, 10, e0127838. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Meneghini, G. Monographia Nostochinearum italicarum addito specimine de Rivulariis. Mem. R. Accad. Delle Sci. Torino Ser. 2 1843, 5, 1–143. [Google Scholar]
- Korshikov, A.A. On some new organisms from the groups Volvocales and Protococcales, and on the genetic relations of these groups. Archiv. Protistenkd. 1926, 55, 439–503. [Google Scholar]
- Darling, R.B.; Friedmann, E.I.; Broady, P.A. Heterococcus endolithicus sp. nov. (Xanthophyceae) and other terrestrial Heterococcus species from Antarctica—Morphological changes during life-history and response to temperature. J. Phycol. 1987, 23, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Neustupa, J.; Št’astný, J.; Hodač, L. Temperature-related phenotypic plasticity in the green microalga Micrasterias rotata. Aquat. Microb. Ecol. 2008, 51, 77–86. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Howe, A.; James, E.R.; Keeling, P.J. Morphological Diversity between Culture Strains of a Chlorarachniophyte, Lotharella globosa. PLoS ONE 2011, 6, e23193. [Google Scholar] [CrossRef]
- Rybalka, N.; Wolf, M.; Andersen, R.A.; Friedl, T. Congruence of chloroplast- and nuclear-encoded DNA sequence variations used to assess species boundaries in the soil microalga Heterococcus (Stramenopiles, Xanthophyceae). BMC Evol. Biol. 2013, 13, 39. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.-H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef]
- Watanabe, S.; Lewis, L.A. Phylogenetic interpretation of light and electron microscopic features of selected members of the phylogroup Moewusinia (Chlorophyceae), with new generic taxonomy. Phycologia 2017, 56, 329–353. [Google Scholar] [CrossRef]
- Brown, R.M., Jr.; McLean, R.J. New taxonomic criteria in classification of Chlorococcum species. II. Pyrenoid fine structure (2). J. Phycol. 1969, 5, 114–118. [Google Scholar] [CrossRef]
- Temraleeva, A.D.; Moslalenko, S.V. Application of Morphological and Molecular Systematics for Identification of Green Microalgae of the Genus Chlorococcum and Some Closely Related Taxa. Microbiology 2019, 88, 27–38. [Google Scholar] [CrossRef]
- Müller, T.; Philippi, N.; Dandekar, T.; Schultz, J.; Wolf, M. Distinguishing species. RNA 2007, 13, 1469–1472. [Google Scholar] [CrossRef]
- Rindi, F.; McIvor, L.; Guiry, M.D. The Prasiolales (Chlorophyta) of Atlantic Europe: An assessment based on morphological, molecular, and ecological data, including the characterization of Rosenvingiella radicans (Kützing) comb. nov. J. Phycol. 2004, 40, 977–997. [Google Scholar] [CrossRef]
- Peksa, O.; Škaloud, P. Do photobionts influence the ecology of lichens? A case study of environmental preferences in symbiotic green alga Asterochloris (Trebouxiophyceae). Mol. Ecol. 2011, 20, 3936–3948. [Google Scholar] [CrossRef]
- Ryšánek, D.; Holzinger, A.; Škaloud, P. Influence of substrate and pH on the diversity of the aeroterrestrial alga Klebsormidium (Klebsormidiales, Streptophyta): A potentially important factor for sympatric speciation. Phycologia 2016, 55, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Malavasi, V.; Škaloud, P.; Rindi, F.; Tempesta, S.; Paoletti, M.; Pasqualetti, M. DNA-Based Taxonomy in Ecologically Versatile Microalgae: A Re- Evaluation of the Species Concept within the Coccoid Green Algal Genus Coccomyxa (Trebouxiophyceae, Chlorophyta). PLoS ONE 2016, 11, e0151137. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.R.; Casamatta, D.A. Recognizing cyanobacterial diversity through adoption of a new species paradigm. Algol. Stud. 2005, 116, 71–93. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain Identifier | 18S rRNA | rbcL | tufA | ITS |
|---|---|---|---|---|---|
| Chlorococcum citriforme | SAG 62.80 | KM020100 | - | LT594531 | LT594562 |
| Chlorococcum costatozygotum | SAG 20.95 | - | - | LT594535 | LT594566 |
| Chlorococcum diplobionticum | SAG 32.95 | - | - | LT594536 | LT594567 |
| Chlorococcum echinozygotum | SAG 213-5 | KM020131 | EF113430 | LT594526 | LT594557 |
| Chlorococcum hypnosporum | SAG 213-6 | JN904003 | LT594545 | LT594527 | LT594558 |
| Chlorococcum infusionum | SAG 10.86 | KM020174 | LT594548 | LT594534 | LT594565 |
| Chlorococcum isabeliense | SAG 65.80 | KM020106 | - | LT594532 | LT594563 |
| Chlorococcum minutum | SAG 213-7 | KM020099 | LT594546 | LT594528 | LT594559 |
| Chlorococcum oleofaciens | SAG 213-11 | AB983608 | - | LT594530 | LT594561 |
| Chlorococcum sphacosum | SAG 66.80 | KM020102 | LT594547 | LT594533 | LT594564 |
| Chlorococcum tatrense | UTEX 2227 | - | LT594550 | LT594538 | LT594569 |
| Chlorococcum vacuolatum | SAG 213-8 | KM020107 | - | LT594529 | LT594560 |
| Chlorococcum sp. | CCCryo 194-04 | - | LT594551 | LT594539 | HQ404881 |
| Chlorococcum sp. | CCCryo 205-05 | - | - | LT594540 | HQ404882 |
| Fasciculochloris boldii | SAG 27.95 | - | LT594552 | LT594541 | LT594570 |
| Macrochloris rubrioleum | CCCryo 340b-08 | GU117573 | - | LT989896 | AB983643 |
| Pleurastrum insigne | SAG 30.93 | AB983614 | EF113464 | LT594537 | LT594568 |
| Pleurastrum sp. 04/16B * | CCCryo 469-16 | LT594553 | LT594543 | LT594524 | LT594555 |
| Pleurastrum sp. A08 * | CCCryo 470-16 | LT594554 | LT594544 | LT594525 | LT594556 |
| Tetracystis aeria | SAG 89.80 | JN903990 | EF113476 | LT594542 | - |
| Updated Species Name | Strain Identifier | Geographical Data | Climate Zone |
|---|---|---|---|
| Pleurastrum insigne Chodat | SAG 30.93T | Soil from banks of brooklet Kahlquelle, Biebergemuend/Spessart, Hessen, Germany | Dfb |
| SAG 62.80T | Soil from peat bog near Elkart, IN, USA | Dfb | |
| SAG 66.80T | Soil from Sphagnum bog, Falmouth, MA, USA | Dfb | |
| SAG 213-11T | Field soil at Duanesburg, NY, USA | Dfb | |
| UTEX 2227T | Snow detritus, Belianske Tatras (Belianske Tatry) mountains, Slovakia | Dfb | |
| Pleurastrum microstigmatum (P.A. Archibald & Bold K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | UTEX 1777T | Peat bog at Elkhart, IN, USA | Dfc |
| CCAP 11/52 | Snow at Port Barrow, AK, USA | Dfc | |
| Pleurastrum rubrioleum (Yur. Kawasaki & Nakada) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | CCCryo 340b-08T | Plastic bin/sewage cover in the vicinity west of the “Blaues Haus” in Ny-Ålesund, Spitsbergen, Svalbard, Norway | ET |
| CCCryo 006-99 | East end of Zeppelinhamna near Ny-Ålesund, Spitsbergen, Svalbard, Norway | ET | |
| CCCryo 194-04 | Plastic bin/sewage cover in the vicinity west of the “Blaues Haus” in Ny-Ålesund, Spitsbergen, Svalbard, Norway | ET | |
| CCCryo 205-05 | Waste water runoff, Longyearbyen,, Spitsbergen, Svalbard, Norway | ET | |
| CCCryo 469-16 | Water pond near the OASI telescope hill, Mario Zucchelli Station, Terra Nova Bay, Victoria Land, Antarctica | EF | |
| CCCryo 470-16 | Snow at Edmonson Point, Wood Bay, Victoria Land, Antarctica | EF | |
| II4 | Ace Lake, Vestfold Hills, Antarctica | EF | |
| VI8 | Lake Fryxell, McMurdo Dry Valleys, Antarctica | EF | |
| Pleurastrum isabeliense (P.A. Archibald & Bold) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | SAG 65.80T | Coastal sand, Port Isabel, TX, USA | Cfa |
| UTEX 1785T | Coastal sand, Port Isabel, TX, USA | Cfa | |
| Pleurastrum vacuolatum (R.C. Starr) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | SAG 213-8T | Soil from Cape Flats, South Africa | Csb |
| Pleurastrum minutum (R.C. Starr) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | SAG 213-7T | Soil from Mumbai, India | Am |
| Pleurastrum aquaticum (Archibald) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | UTEX 2222T | Treasure Lake (Lake El Tesero), Zapata, Cuba | Aw |
| Pleurastrum diplobionticum (Herndon) K. Sciuto, M.A. Wolf, M. Mistri & I. Moro comb. nov. | SAG 32.95T | Soil from corn field near Daniel Town, Jamaica | Af |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciuto, K.; Wolf, M.A.; Mistri, M.; Moro, I. Appraisal of the Genus Pleurastrum (Chlorophyta) Based on Molecular and Climate Data. Diversity 2023, 15, 650. https://doi.org/10.3390/d15050650
Sciuto K, Wolf MA, Mistri M, Moro I. Appraisal of the Genus Pleurastrum (Chlorophyta) Based on Molecular and Climate Data. Diversity. 2023; 15(5):650. https://doi.org/10.3390/d15050650
Chicago/Turabian StyleSciuto, Katia, Marion A. Wolf, Michele Mistri, and Isabella Moro. 2023. "Appraisal of the Genus Pleurastrum (Chlorophyta) Based on Molecular and Climate Data" Diversity 15, no. 5: 650. https://doi.org/10.3390/d15050650
APA StyleSciuto, K., Wolf, M. A., Mistri, M., & Moro, I. (2023). Appraisal of the Genus Pleurastrum (Chlorophyta) Based on Molecular and Climate Data. Diversity, 15(5), 650. https://doi.org/10.3390/d15050650