
Development of Conventional Multiplex PCR Assays for the Identification of 21 West Palaearctic Biting Midge Taxa (Diptera: Ceratopogonidae) Belonging to the Culicoides Subgenus Culicoides, including Recently Discovered Species and Genetic Variants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biting Midge Collection
2.2. Genetic Identification of Field-Collected Biting Midges
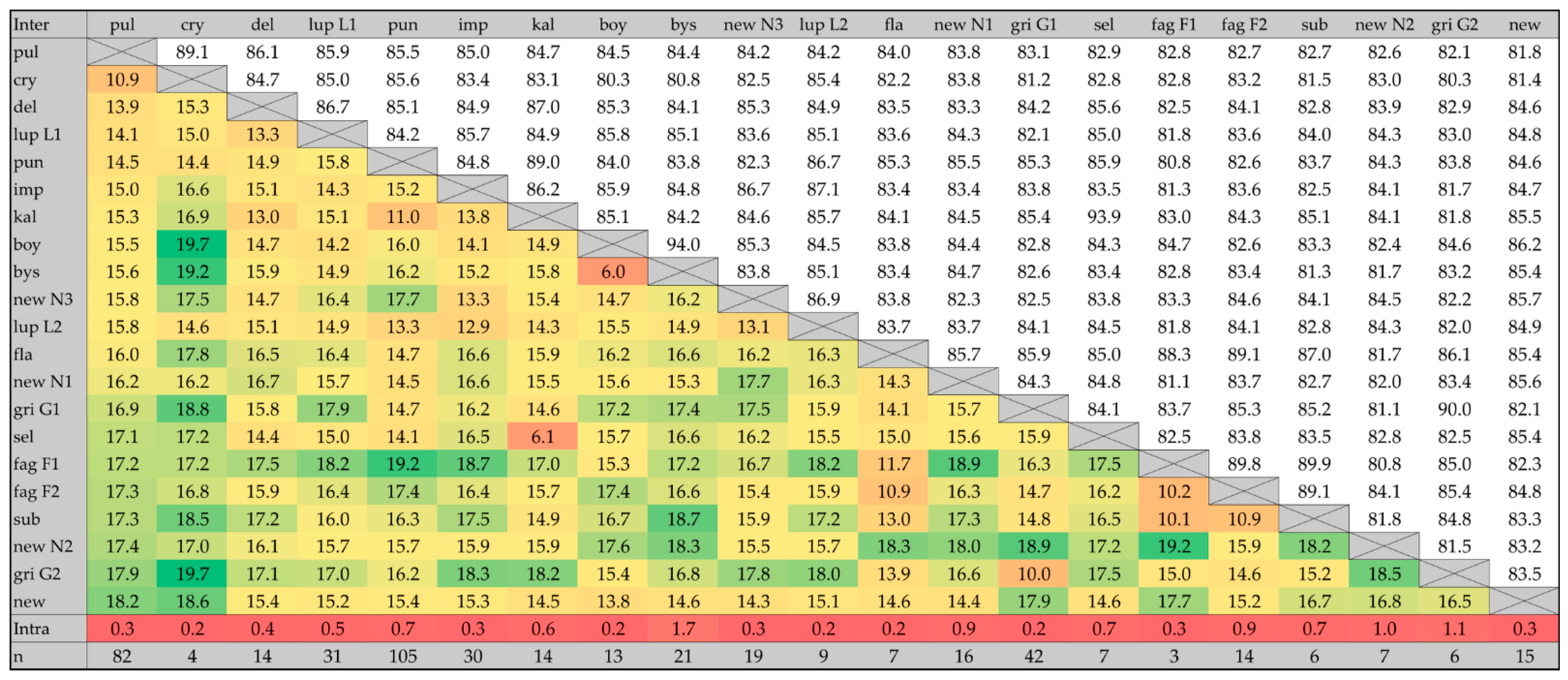
2.3. COI Data Analysis and Primer Design
2.4. Multiplex PCRs
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides biting midges: Their role as arbovirus vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Meiswinkel, R.; Gomulski, L.M.; Delécolle, J.-C.; Goffredo, M.; Gasperi, G. The taxonomy of Culicoides vector complexes-unfinished business. Vet. Ital. 2004, 40, 151–159. [Google Scholar]
- Borkent, A. The biting midges, the Ceratopogonidae (Diptera). In Biology of Disease Vectors, 2nd ed.; Marquardt, W., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 113–126. [Google Scholar]
- Mellor, P.S.; Pitzolis, G. Observations on breeding sites and light-trap collections of Culicoides during an outbreak of bluetongue in Cyprus. Bull. Entomol. Res. 1979, 69, 229–234. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boned, J.; Hamblin, C.; Graham, S. Isolations of African horse sickness virus from vector insects made during the 1988 epizootic in Spain. Epidemiol. Infect. 1990, 105, 447–454. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Walldorf, V.; Klimpel, S.; Jahn, B.; Jaeger, F.; Eschweiler, J.; Hoffmann, B.; Beer, M. First occurrence of Culicoides obsoletus-transmitted Bluetongue virus epidemic in Central Europe. Parasitol. Res. 2007, 101, 219–228. [Google Scholar] [CrossRef]
- Meiswinkel, R.; van Rijn, P.; Leijs, P.; Goffredo, M. Potential new Culicoides vector of bluetongue virus in northern Europe. Vet. Rec. 2007, 161, 564–565. [Google Scholar] [CrossRef]
- Dijkstra, E.; van der Ven, I.J.K.; Meiswinkel, R.; Hölzel, D.R.; van Rijn, P.A. Culicoides chiopterus as a potential vector of bluetongue virus in Europe. Vet. Rec. 2008, 162, 422. [Google Scholar] [CrossRef]
- Hoffmann, B.; Bauer, B.; Bauer, C.; Bätza, H.J.; Beer, M.; Clausen, P.H.; Geier, M.; Gethmann, J.M.; Kiel, E.; Liebisch, G.; et al. Monitoring of putative vectors of bluetongue virus serotype 8, Germany. Emerg. Infect. Dis. 2009, 15, 1481–1484. [Google Scholar] [CrossRef]
- Vanbinst, T.; Vandenbussche, F.; Vandemeulebroucke, E.; de Leeuw, I.; Deblauwe, I.; de Deken, G.; Madder, M.; Haubruge, E.; Losson, B.; de Clercq, K. Bluetongue virus detection by real-time RT-PCR in Culicoides captured during the 2006 epizootic in Belgium and development of an internal control. Transbound. Emerg. Dis. 2009, 56, 170–177. [Google Scholar] [CrossRef]
- Romón, P.; Higuera, M.; Delécolle, J.-C.; Baldet, T.; Aduriz, G.; Goldarazena, A. Phenology and attraction of potential Culicoides vectors of bluetongue virus in Basque Country (northern Spain). Vet. Parasitol. 2012, 186, 415–424. [Google Scholar] [CrossRef]
- Goffredo, M.; Catalani, M.; Federici, V.; Portanti, O.; Marini, V.; Mancini, G.; Quaglia, M.; Santilli, A.; Teodori, L.; Savini, G. Vector species of Culicoides midges implicated in the 2012–2014 Bluetongue epidemics in Italy. Vet. Ital. 2015, 51, 131–138. [Google Scholar] [CrossRef]
- Foxi, C.; Delrio, G.; Falchi, G.; Marche, M.G.; Satta, G.; Ruiu, L. Role of different Culicoides vectors (Diptera: Ceratopogonidae) in bluetongue virus transmission and overwintering in Sardinia (Italy). Parasit. Vectors 2016, 9, 440. [Google Scholar] [CrossRef]
- Foxi, C.; Meloni, G.; Puggioni, G.; Manunta, D.; Rocchigiani, A.; Vento, L.; Cabras, P.; Satta, G. Bluetongue virus detection in new Culicoides species in Sardinia, Italy. Vet. Rec. 2019, 184, 621. [Google Scholar] [CrossRef]
- Rasmussen, L.D.; Kristensen, B.; Kirkeby, C.; Rasmussen, T.B.; Belsham, G.J.; Bødker, R.; Bøtner, A. Culicoids as vectors of Schmallenberg virus. Emerg. Infect. Dis. 2012, 18, 1204–1206. [Google Scholar] [CrossRef]
- de Regge, N.; Deblauwe, I.; de Deken, R.; Vantieghem, P.; Madder, M.; Geysen, D.; Smeets, F.; Losson, B.; van den Berg, T.; Cay, A.B. Detection of Schmallenberg virus in different Culicoides spp. by real-time RT-PCR. Transbound. Emerg. Dis. 2012, 59, 471–475. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Meiswinkel, R.; van Weezep, E.; van Sloet Oldruitenborgh-Oosterbaan, M.M.; Kooi, E.A. Schmallenberg virus in Culicoides spp. biting midges, the Netherlands, 2011. Emerg. Infect. Dis. 2013, 19, 106–109. [Google Scholar] [CrossRef]
- Goffredo, M.; Monaco, F.; Capelli, G.; Quaglia, M.; Federici, V.; Catalani, M.; Montarsi, F.; Polci, A.; Pinoni, C.; Calistri, P.; et al. Schmallenberg virus in Italy: A retrospective survey in Culicoides stored during the bluetongue Italian surveillance program. Prev. Vet. Med. 2013, 111, 230–236. [Google Scholar] [CrossRef]
- Larska, M.; Polak, M.P.; Grochowska, M.; Lechowski, L.; Związek, J.S.; Zmudziński, J.F. First report of Schmallenberg virus infection in cattle and midges in Poland. Transbound. Emerg. Dis. 2013, 60, 97–101. [Google Scholar] [CrossRef]
- Larska, M.; Lechowski, L.; Grochowska, M.; Żmudziński, J.F. Detection of the Schmallenberg virus in nulliparous Culicoides obsoletus/scoticus complex and C. punctatus—The possibility of transovarial virus transmission in the midge population and of a new vector. Vet. Microbiol. 2013, 166, 467–473. [Google Scholar] [CrossRef]
- Balenghien, T.; Pagès, N.; Goffredo, M.; Carpenter, S.; Augot, D.; Jacquier, E.; Talavera, S.; Monaco, F.; Depaquit, J.; Grillet, C.; et al. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev. Vet. Med. 2014, 116, 360–369. [Google Scholar] [CrossRef]
- Rasmussen, L.D.; Kirkeby, C.; Bødker, R.; Kristensen, B.; Rasmussen, T.B.; Belsham, G.J.; Bøtner, A. Rapid spread of Schmallenberg virus-infected biting midges (Culicoides spp.) across Denmark in 2012. Transbound. Emerg. Dis. 2014, 61, 12–16. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Meiswinkel, R.; van Weezep, E.; Kooi, E.A.; van der Poel, W.H.M. Schmallenberg virus in Culicoides biting midges in the Netherlands in 2012. Transbound. Emerg. Dis. 2015, 62, 339–342. [Google Scholar] [CrossRef]
- de Regge, N.; de Deken, R.; Fassotte, C.; Losson, B.; Deblauwe, I.; Madder, M.; Vantieghem, P.; Tomme, M.; Smeets, F.; Cay, A.B. Culicoides monitoring in Belgium in 2011: Analysis of spatiotemporal abundance, species diversity and Schmallenberg virus detection. Med. Vet. Entomol. 2015, 29, 263–275. [Google Scholar] [CrossRef]
- Pagès, N.; Talavera, S.; Verdún, M.; Pujol, N.; Valle, M.; Bensaid, A.; Pujols, J. Schmallenberg virus detection in Culicoides biting midges in Spain: First laboratory evidence for highly efficient infection of Culicoides of the Obsoletus Complex and Culicoides imicola. Transbound. Emerg. Dis. 2018, 65, e1–e6. [Google Scholar] [CrossRef]
- Ségard, A.; Gardès, L.; Jacquier, E.; Grillet, C.; Mathieu, B.; Rakotoarivony, I.; Setier-Rio, M.-L.; Chavernac, D.; Cêtre-Sossah, C.; Balenghien, T.; et al. Schmallenberg virus in Culicoides Latreille (Diptera: Ceratopogonidae) populations in France during 2011–2012 outbreak. Transbound. Emerg. Dis. 2018, 65, e94–e103. [Google Scholar] [CrossRef]
- Borkent, A.; Dominiak, P. Catalog of the biting midges of the world (Diptera: Ceratopogonidae). Zootaxa 2020, 4787, 1–377. [Google Scholar] [CrossRef]
- Lassen, S.B.; Nielsen, S.A.; Skovgård, H.; Kristensen, M. Molecular differentiation of Culicoides biting midges (Diptera: Ceratopogonidae) from the subgenus Culicoides Latreille in Denmark. Parasitol. Res. 2012, 110, 1765–1771. [Google Scholar] [CrossRef]
- Ramilo, D.W.; Diaz, S.; Da Pereira Fonseca, I.; Delécolle, J.-C.; Wilson, A.; Meireles, J.; Lucientes, J.; Ribeiro, R.; Boinas, F. First report of 13 species of Culicoides (Diptera: Ceratopogonidae) in mainland Portugal and Azores by morphological and molecular characterization. PLoS ONE 2012, 7, e34896. [Google Scholar] [CrossRef]
- Ramilo, D.; Garros, C.; Mathieu, B.; Benedet, C.; Allène, X.; Silva, E.; Alexandre-Pires, G.; Da Fonseca, I.P.; Carpenter, S.; Rádrová, J.; et al. Description of Culicoides paradoxalis sp. nov. from France and Portugal (Diptera: Ceratopogonidae). Zootaxa 2013, 3745, 243–256. [Google Scholar] [CrossRef]
- Talavera, S.; Muñoz-Muñoz, F.; Verdún, M.; Pagès, N. Morphology and DNA barcoding reveal three species in one: Description of Culicoides cryptipulicaris sp. nov. and Culicoides quasipulicaris sp. nov. in the subgenus Culicoides. Med. Vet. Entomol. 2017, 31, 178–191. [Google Scholar] [CrossRef]
- Goffredo, M.; Meiswinkel, R. Entomological surveillance of bluetongue in Italy: Methods of capture, catch analysis and identification of Culicoides biting midges. Vet. Ital. 2004, 40, 260–265. [Google Scholar] [PubMed]
- Meiswinkel, R.; Baldet, T.; de Deken, R.; Takken, W.; Delécolle, J.-C.; Mellor, P.S. The 2006 outbreak of bluetongue in northern Europe–the entomological perspective. Prev. Vet. Med. 2008, 87, 55–63. [Google Scholar] [CrossRef]
- Schwenkenbecher, J.M.; Mordue, A.J.; Switek, K.; Piertney, S.B. Discrimination of Culicoides midge larvae using multiplex polymerase chain reaction assays based on DNA sequence variation at the mitochondrial cytochrome C oxidase I gene. J. Med. Entomol. 2009, 46, 610–614. [Google Scholar] [CrossRef]
- Yanase, T.; Matsumoto, Y.; Matsumori, Y.; Aizawa, M.; Hirata, M.; Kato, T.; Shirafuji, H.; Yamakawa, M.; Tsuda, T.; Noda, H. Molecular identification of field-collected Culicoides larvae in the southern part of Japan. J. Med. Entomol. 2013, 50, 1105–1110. [Google Scholar] [CrossRef]
- Jin, T.; Husseneder, C.; Foil, L. Assigning Culicoides larvae to species using DNA barcoding of adult females and phylogenetic associations. Parasit. Vectors 2022, 15, 349. [Google Scholar] [CrossRef]
- Schwenkenbecher, J.M.; Mordue, A.J.; Piertney, S.B. Phylogenetic analysis indicates that Culicoides dewulfi should not be considered part of the Culicoides obsoletus complex. Bull. Entomol. Res. 2009, 99, 371–375. [Google Scholar] [CrossRef]
- Delécolle, J.-C. Nouvelle Contribution à L’étude Systématique et Iconographique des Espèces du Genre Culicoides (Diptera: Ceratopogonidae) du Nord-Est de la France. Ph.D. Thesis, University of Strasbourg, Strasbourg, France, 1985. [Google Scholar]
- Harrup, L.E.; Bellis, G.A.; Balenghien, T.; Garros, C. Culicoides Latreille (Diptera: Ceratopogonidae) taxonomy: Current challenges and future directions. Infect. Genet. Evol. 2015, 30, 249–266. [Google Scholar] [CrossRef]
- Lunt, D.H.; Zhang, D.X.; Szymura, J.M.; Hewitt, G.M. The insect cytochrome oxidase I gene: Evolutionary patterns and conserved primers for phylogenetic studies. Insect Mol. Biol. 1996, 5, 153–165. [Google Scholar] [CrossRef]
- Dobler, S.; Farrell, B.D. Host use evolution in Chrysochus milkweed beetles: Evidence from behaviour, population genetics and phylogeny. Mol. Ecol. 1999, 8, 1297–1307. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Pagès, N.; Muñoz-Muñoz, F.; Talavera, S.; Sarto, V.; Lorca, C.; Núñez, J.I. Identification of cryptic species of Culicoides (Diptera: Ceratopogonidae) in the subgenus Culicoides and development of species-specific PCR assays based on barcode regions. Vet. Parasitol. 2009, 165, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.A.; Kristensen, M. Delineation of Culicoides species by morphology and barcode exemplified by three new species of the subgenus Culicoides (Diptera: Ceratopogonidae) from Scandinavia. Parasit. Vectors 2015, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Wenk, C.E.; Kaufmann, C.; Schaffner, F.; Mathis, A. Molecular characterization of Swiss Ceratopogonidae (Diptera) and evaluation of real-time PCR assays for the identification of Culicoides biting midges. Vet. Parasitol. 2012, 184, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Ander, M.; Troell, K.; Chirico, J. Barcoding of biting midges in the genus Culicoides: A tool for species determination. Med. Vet. Entomol. 2013, 27, 323–331. [Google Scholar] [CrossRef]
- Sarvašová, A.; Kočišová, A.; Halán, M.; Delécolle, J.-C.; Mathieu, B. Morphological and molecular analysis of the genus Culicoides (Diptera: Ceratopogonidae) in Slovakia with five new records. Zootaxa 2014, 3872, 541–560. [Google Scholar] [CrossRef]
- Yildirim, A.; Dik, B.; Duzlu, O.; Onder, Z.; Ciloglu, A.; Yetismis, G.; Inci, A. Genetic diversity of Culicoides species within the Pulicaris Complex (Diptera: Ceratopogonidae) in Turkey inferred from mitochondrial COI gene sequences. Acta Trop. 2019, 190, 380–388. [Google Scholar] [CrossRef]
- Zittra, C.; Wöss, G.; van der Vloet, L.; Bakran-Lebl, K.; Shahi Barogh, B.; Sehnal, P.; Fuehrer, H.-P. Barcoding of the genus Culicoides (Diptera: Ceratopogonidae) in Austria—An update of the species inventory including the first records of three species in Austria. Pathogens 2020, 9, 406. [Google Scholar] [CrossRef]
- Nolan, D.V.; Carpenter, S.; Barber, J.; Mellor, P.S.; Dallas, J.F.; Mordue Luntz, A.J.; Piertney, S.B. Rapid diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet. Microbiol. 2007, 124, 82–94. [Google Scholar] [CrossRef]
- Bellis, G.; Kim, H.-C.; Kim, M.-S.; Klein, T.A.; Lee, D.-K.; Gopurenko, D. Three species of Culicoides Latreille (Diptera: Ceratopogonidae) newly recorded from the Republic of Korea. Zootaxa 2013, 3718, 171–182. [Google Scholar] [CrossRef]
- Rot, A.; Meiswinkel, R.; Fleker, M.; Blum, S.E.; Behar, A. Towards modernizing the taxonomy of Mediterranean Culicoides using classical morphology, mtDNA barcoding, and MALDI-TOF MS protein profiling. Acta Trop. 2020, 211, 105628. [Google Scholar] [CrossRef]
- Campbell, J.A.; Pelham-Clinton, E.C. A taxonomic review of the British species of Culicoides Latreille (Diptera, Ceratopogonidæ). Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1960, 67, 181–302. [Google Scholar] [CrossRef]
- Glukhova, V.M. Krovososuščie Mokrecy Rodov Culicoides i Forcipomyia (Ceratopogonidae); Nauka: Leningrad, Russia, 1989; ISBN 9785020257603. [Google Scholar]
- Mathieu, B.; Cêtre-Sossah, C.; Garros, C.; Chavernac, D.; Balenghien, T.; Carpenter, S.; Setier-Rio, M.L.; Vignes-Lebbe, R.; Ung, V.; Candolfi, E.; et al. Development and validation of IIKC: An interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasit. Vectors 2012, 5, 137. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, K.; Werner, D.; Hoffmann, B.; Kampen, H. PCR identification of culicoid biting midges (Diptera, Ceratopogonidae) of the Obsoletus Complex including putative vectors of bluetongue and Schmallenberg viruses. Parasit. Vectors 2012, 5, 213. [Google Scholar] [CrossRef] [PubMed]
- Henegariu, O.; Heerema, N.A.; Dlouhy, S.R.; Vance, G.H.; Vogt, P.H. Multiplex PCR: Critical parameters and step-by-step protocol. Biotechniques 1997, 23, 504–511. [Google Scholar] [CrossRef]
- Elnifro, E.M.; Ashshi, A.M.; Cooper, R.J.; Klapper, P.E. Multiplex PCR: Optimization and application in diagnostic virology. Cli. Microbiol. Rev. 2000, 13, 559–570. [Google Scholar] [CrossRef]
- Newton, C.R.; Graham, A. PCR; Spektrum Akad. Verl.: Heidelberg, Germany, 1994; ISBN 3-86025-236-4. [Google Scholar]
- Pagès, N.; Sarto i Monteys, V. Differentiation of Culicoides obsoletus and Culicoides scoticus (Diptera: Ceratopogonidae) based on mitochondrial cytochrome oxidase subunit I. J. Med. Entomol. 2005, 42, 1026–1034. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Baldet, T.; Delécolle, J.-C.; Mathieu, B.; Perrin, A.; Grillet, C.; Albina, E. Molecular detection of Culicoides spp. and Culicoides imicola, the principal vector of bluetongue (BT) and African horse sickness (AHS) in Africa and Europe. Vet. Res. 2004, 35, 325–337. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Mathieu, B.; Setier-Rio, M.-L.; Grillet, C.; Baldet, T.; Delécolle, J.-C.; Albina, E. Development and evaluation of a real-time quantitative PCR assay for Culicoides imicola, one of the main vectors of bluetongue (BT) and African horse sickness (AHS) in Africa and Europe. Res. Vet. Sci. 2008, 85, 372–382. [Google Scholar] [CrossRef]
- Mathieu, B.; Perrin, A.; Baldet, T.; Delécolle, J.-C.; Albina, E.; Cêtre-Sossah, C. Molecular identification of western European species of Obsoletus Complex (Diptera: Ceratopogonidae) by an internal transcribed spacer-1 rDNA multiplex polymerase chain reaction assay. J. Med. Entomol. 2007, 44, 1019–1025. [Google Scholar] [CrossRef]
- Stephan, A.; Clausen, P.-H.; Bauer, B.; Steuber, S. PCR identification of Culicoides dewulfi midges (Diptera: Ceratopogonidae), potential vectors of bluetongue in Germany. Parasitol. Res. 2009, 105, 367–371. [Google Scholar] [CrossRef]
- Mathieu, B.; Delecolle, J.-C.; Garros, C.; Balenghien, T.; Setier-Rio, M.-L.; Candolfi, E.; Cêtre-Sossah, C. Simultaneous quantification of the relative abundance of species complex members: Application to Culicoides obsoletus and Culicoides scoticus (Diptera: Ceratopogonidae), potential vectors of bluetongue virus. Vet. Parasitol. 2011, 182, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Gomulski, L.M.; Meiswinkel, R.; Delécolle, J.C.; Goffredo, M.; Gasperi, G. Phylogenetic relationships of the subgenus Avaritia Fox, 1955 including Culicoides obsoletus (Diptera, Ceratopogonidae) in Italy based on internal transcribed spacer 2 ribosomal DNA sequences. Syst. Entomol. 2005, 30, 619–631. [Google Scholar] [CrossRef]
- Monaco, F.; Benedetto, L.; Di Marcello, V.; Lelli, R.; Goffredo, M. Development and preliminary evaluation of a real-time polymerase chain reaction for the identification of Culicoides obsoletus sensu strictu, C. scoticus and C. montanus in the Obsoletus Complex in Italy. Vet. Ital. 2010, 46, 215–220. [Google Scholar]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef]
- Dallas, J.F.; Cruickshank, R.H.; Linton, Y.-M.; Nolan, D.V.; Patakakis, M.; Braverman, Y.; Capela, R.; Capela, M.; Pena, I.; Meiswinkel, R.; et al. Phylogenetic status and matrilineal structure of the biting midge, Culicoides imicola, in Portugal, Rhodes and Israel. Med. Vet. Entomol. 2003, 17, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Kwok, S.; Kellogg, D.E.; McKinney, N.; Spasic, D.; Goda, L.; Levenson, C.; Sninsky, J.J. Effects of primer-template mismatches on the polymerase chain reaction: Human immunodeficiency virus type 1 model studies. Nucleic Acids Res. 1990, 18, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Mendelman, L.V.; Petruska, J.; Goodman, M.F. Base mispair extension kinetics. Comparison of DNA polymerase alpha and reverse transcriptase. J. Biol. Chem. 1990, 265, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Creighton, S.; Huang, M.M.; Cai, H.; Arnheim, N.; Goodman, M.F. Base mispair extension kinetics. Binding of avian myeloblastosis reverse transcriptase to matched and mismatched base pair termini. J. Biol. Chem. 1992, 267, 2633–2639. [Google Scholar] [CrossRef]
- Huang, M.M.; Arnheim, N.; Goodman, M.F. Extension of base mispairs by Taq DNA polymerase: Implications for single nucleotide discrimination in PCR. Nucleic Acids Res. 1992, 20, 4567–4573. [Google Scholar] [CrossRef]
- Joyce, C.M.; Sun, X.C.; Grindley, N.D. Reactions at the polymerase active site that contribute to the fidelity of Escherichia coli DNA polymerase I (Klenow fragment). J. Biol. Chem. 1992, 267, 24485–24500. [Google Scholar] [CrossRef]
- Day, J.P.; Bergstrom, D.; Hammer, R.P.; Barany, F. Nucleotide analogs facilitate base conversion with 3′ mismatch primers. Nucleic Acids Res. 1999, 27, 1810–1818. [Google Scholar] [CrossRef]
- Stadhouders, R.; Pas, S.D.; Anber, J.; Voermans, J.; Mes, T.H.M.; Schutten, M. The effect of primer-template mismatches on the detection and quantification of nucleic acids using the 5′ nuclease assay. J. Mol. Diagn. 2010, 12, 109–117. [Google Scholar] [CrossRef]
- Kiel, E.; Liebisch, G.; Focke, R.; Liebisch, A.; Werner, D. Monitoring of Culicoides at 20 locations in northwest Germany. Parasitol. Res. 2009, 105, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H.; Walldorf, V.; Klimpel, S.; Schaub, G.; Kiel, E.; Focke, R.; Liebisch, G.; Liebisch, A.; Werner, D.; Bauer, C.; et al. Bluetongue disease in Germany (2007–2008): Monitoring of entomological aspects. Parasitol. Res. 2009, 105, 313–319. [Google Scholar] [CrossRef]
- Vorsprach, B.; Meiser, C.K.; Werner, D.; Balczun, C.; Schaub, G.A. Monitoring of Ceratopogonidae in southwest Germany. Parasitol. Res. 2009, 105, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Werner, D.; Groschupp, S.; Bauer, C.; Kampen, H. Breeding habitat preferences of major Culicoides species (Diptera: Ceratopogonidae) in Germany. Int. J. Environ. Res. Public Health 2020, 17, 5000. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Almeras, L.; Raoult, D.; Parola, P. Emerging tools for identification of arthropod vectors. Future Microbiol. 2016, 11, 549–566. [Google Scholar] [CrossRef]
- Kaufmann, C.; Ziegler, D.; Schaffner, F.; Carpenter, S.; Pflüger, V.; Mathis, A. Evaluation of matrix-assisted laser desorption/ionization time of flight mass spectrometry for characterization of Culicoides nubeculosus biting midges. Med. Vet. Entomol. 2011, 25, 32–38. [Google Scholar] [CrossRef]
- Kaufmann, C.; Schaffner, F.; Ziegler, D.; Pflüger, V.; Mathis, A. Identification of field-caught Culicoides biting midges using matrix-assisted laser desorption/ionization time of flight mass spectrometry. Parasitology 2012, 139, 248–258. [Google Scholar] [CrossRef]
- Kaufmann, C.; Steinmann, I.C.; Hegglin, D.; Schaffner, F.; Mathis, A. Spatio-temporal occurrence of Culicoides biting midges in the climatic regions of Switzerland, along with large scale species identification by MALDI-TOF mass spectrometry. Parasit. Vectors 2012, 5, 246. [Google Scholar] [CrossRef]
- Sambou, M.; Aubadie-Ladrix, M.; Fenollar, F.; Fall, B.; Bassene, H.; Almeras, L.; Sambe-Ba, B.; Perrot, N.; Chatellier, S.; Faye, N.; et al. Comparison of matrix-assisted laser desorption ionization-time of flight mass spectrometry and molecular biology techniques for identification of Culicoides (Diptera: Ceratopogonidae) biting midges in Senegal. J. Clin. Microbiol. 2015, 53, 410–418. [Google Scholar] [CrossRef]
- Uhlmann, K.R.; Gibb, S.; Kalkhof, S.; Arroyo-Abad, U.; Schulz, C.; Hoffmann, B.; Stubbins, F.; Carpenter, S.; Beer, M.; von Bergen, M.; et al. Species determination of Culicoides biting midges via peptide profiling using matrix-assisted laser desorption ionization mass spectrometry. Parasit. Vectors 2014, 7, 392. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, I.C.; Pflüger, V.; Schaffner, F.; Mathis, A.; Kaufmann, C. Evaluation of matrix-assisted laser desorption/ionization time of flight mass spectrometry for the identification of ceratopogonid and culicid larvae. Parasitology 2013, 140, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Socolovschi, C.; Flaudrops, C.; Ndiath, M.O.; Sougoufara, S.; Dehecq, J.-S.; Lacour, G.; Berenger, J.-M.; Sokhna, C.S.; Raoult, D.; et al. Matrix-assisted laser desorption ionization—Time of flight mass spectrometry: An emerging tool for the rapid identification of mosquito vectors. PLoS ONE 2013, 8, e72380. [Google Scholar] [CrossRef]
- Bellis, G. Studies on the Taxonomy of Australian Species of Culicoides Latreille (Diptera: Ceratopogonidae). Ph.D. Thesis, University of Queensland, St Lucia, Australia, 2013. [Google Scholar]
- Mignotte, A.; Garros, C.; Gardès, L.; Balenghien, T.; Duhayon, M.; Rakotoarivony, I.; Tabourin, L.; Poujol, L.; Mathieu, B.; Ibañez-Justicia, A.; et al. The tree that hides the forest: Cryptic diversity and phylogenetic relationships in the Palaearctic vector Obsoletus/Scoticus Complex (Diptera: Ceratopogonidae) at the European level. Parasit. Vectors 2020, 13, 265. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| mPCR | Species/Haplotype | Primer Code | Primer Sequence (5′-3′) | Modification (Position) | Amplicon (bp) |
|---|---|---|---|---|---|
| C. bysta | bys-COI-158F | AATCTTACTTCTCTTATCTCTRC | R-wobble (2) | 158 | |
| C. punctatus | pun-COI-227F | TCATATGCGATCAAACGGG | A > C (18) | 227 | |
| A | C. boyi | boy-COI-275F | AGCTATTTCATCAATTCTTGGA | G > C (20) | 275 |
| C. grisescens G2 | gri2-COI-346F | CCACACCTTTCTGCAAACA | C > A (15) | 346 | |
| C. kalix | kal-COI-419F | CCACCCTTCTCTAACATTGC | C > A (18) | 419 | |
| C. grisescens G1 | gri1-COI-463F | GATATAGCTTTCACACGAATG | C > A (9) | 463 | |
| C. fagineus F2 | fag2-COI-151F 3 | TTGCATCTTTCCCTCCCTGTA | T > A (17) | 151 | |
| C. flavipulicaris | fla-COI-215F 3 | CAATCGTATTACTTTTGATCGT | G > C (18) | 215 | |
| B | C. subfagineus | sub-COI-318F 3 | CTGTRGCTTCTGTAGATC | R-wobble (14), G > T (15) | 318 |
| C. fagineus F1 | fag1-COI-420F | TTCCTCCATCTCTTTCCCTAT | C > T (17) | 420 | |
| C. impunctatus | imp-COI-491F | ATTGGTTCCATTAATACTCGGA | none | 491 | |
| C. delta | del-COI-161F | TGCTATATTACTTCTTTTGTCAC | T > A (17) | 161 | |
| C. lupicaris L1 1 | lup1-COI-214F | AATGGAATGTCATTCGACCGT | T > G (13) | 214 | |
| C | C. pulicaris s.s. | pul-COI-313F 2,3 | GCATCCGTAGACTTGGCC | none | 313 |
| C. cryptipulicaris | cry-COI-405F | CGTTACTCTTATTGAGCAGAT | none | 405 | |
| C. lupicaris L2 | lup2-COI-467F | TCCTGATATAGCTTTTCCC | none | 467 | |
| C. newsteadi N2 | new2-COI-139F 3 | CTCCCAGTTCTTGCTGGT | none | 139 | |
| C. newsteadi s.s. | new-COI-231F | TTATTAATATGCGATCCGCC | none | 231 | |
| D | C. newsteadi N3 | new3-COI-296F 3 | CATCTTCTCCCTACACCTG | none | 296 |
| C. newsteadi N1 | new1-COI-351F | TATATCCGCCTCTTTCAAGA | none | 351 | |
| C. selandicus | sel-COI-403F | TGACTATTATTAAGTAGCTTGGTA | T > G (23) | 403 |
| Species/Haplotype DNA | No. Samples | GenBank Accession No. | Multiplex PCR A | Multiplex PCR B | Multiplex PCR C | Multiplex PCR D | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bys | pun | boy | G2 | kal | G1 | F2 | fla | sub | F1 | imp | del | L1 | pul | cry | L2 | N2 | new | N3 | N1 | sel | |||
| C. bysta 1 | 1 | n.a | + (1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. punctatus 2 | 5 | OQ789061-65 | - | + (5) | - | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | + (1) | - |
| C. boyi 1 | 1 | n.a | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. grisescens G2 2 | 1 | OQ789038 | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | + (1) |
| C. kalix 1 | 1 | n.a. | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. grisescens G1 2 | 1 | OQ789037 | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. fagineus F2 2 | 1 | OQ789036 | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. flavipulicaris 1 | 1 | n.a. | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | + (1) | - | - | - | - | - |
| C. subfagineus 1 | 1 | n.a. | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - | - |
| C. fagineus F1 1 | 1 | n.a. | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - | - | - |
| C. impunctatus 2 | 1 | OQ789034 | - | - | - | - | - | - | - | - | - | - | + (1) | + (1) | - | - | - | - | - | - | + (1) | - | - |
| C. delta 2 | 1 | OQ789035 | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - | - |
| C. lupicaris L1 2 | 1 | OQ789039 | - | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - | - | - |
| C. pulicaris s.s. 2 | 6 | OQ789055-60 | - | - | - | - | - | - | - | - | - | - | - | - | - | + (6) | - | - | - | - | - | - | - |
| C. cryptipulicaris 1 | 1 | n.a. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - | - | - |
| C. lupicaris L2 2 | 5 | OQ789040-44 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (5) | - | - | + (4) | - | - |
| C. newsteadi N2 1 | 1 | n.a. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - | - |
| C. newsteadi s.s. 1 | 1 | n.a. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (1) | - | - | - |
| C. newsteadi N3 2 | 4 | OQ789048-51 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (4) | - | - |
| C. newsteadi N1 2 | 3 | OQ789045-47 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (3) | - |
| C. selandicus 2 | 3 | OQ789052-54 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + (2) |
| Species/Haplotype DNA | GenBank Accession No. | Multiplex PCR A | Multiplex PCR B | Multiplex PCR C | Multiplex PCR D | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bys | pun | boy | G2 | kal | G1 | F2 | fla | sub | F1 | imp | del | L1 | pul | cry | L2 | N2 | new | N3 | N1 | sel | ||
| C. achrayi | OQ789066 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. alazanicus | OQ789067 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. chiopterus | OQ789068 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. dewulfi | OQ789069 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + 2,3 | + 2,3 | + 2,3 | + 2,3 | + 2,3 |
| C. festivipennis | OQ789070 | - | - | - | - | - | - | - | - | - | - | - | + 1 | + 1 | + 1 | + 1 | + 1 | - | - | - | - | - |
| C. griseidorsum | OQ789071 | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. imicola | OQ789072 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. kibunensis | OQ789073 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - |
| C. montanus | OQ789074 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. obsoletus clade O1 | OQ789075 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - |
| C. obsoletus clade O2 | OQ789076 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. obsoletus clade O3 | OQ789077 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. pallidicornis | OQ789078 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. pictipennis | OQ789079 | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. poperinghensis | OQ789080 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - |
| C. riethi | OQ789081 | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - |
| C. riouxi | OQ789082 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. salinarius | OQ789083 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. sanguisuga | MK760238 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + 4 | + 4 | + 4 | + 4 | + 4 |
| C. scoticus clade 2 | OQ789084 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C. sinanoensis | MK760244 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dähn, O.; Werner, D.; Mathieu, B.; Kampen, H. Development of Conventional Multiplex PCR Assays for the Identification of 21 West Palaearctic Biting Midge Taxa (Diptera: Ceratopogonidae) Belonging to the Culicoides Subgenus Culicoides, including Recently Discovered Species and Genetic Variants. Diversity 2023, 15, 699. https://doi.org/10.3390/d15060699
Dähn O, Werner D, Mathieu B, Kampen H. Development of Conventional Multiplex PCR Assays for the Identification of 21 West Palaearctic Biting Midge Taxa (Diptera: Ceratopogonidae) Belonging to the Culicoides Subgenus Culicoides, including Recently Discovered Species and Genetic Variants. Diversity. 2023; 15(6):699. https://doi.org/10.3390/d15060699
Chicago/Turabian StyleDähn, Oliver, Doreen Werner, Bruno Mathieu, and Helge Kampen. 2023. "Development of Conventional Multiplex PCR Assays for the Identification of 21 West Palaearctic Biting Midge Taxa (Diptera: Ceratopogonidae) Belonging to the Culicoides Subgenus Culicoides, including Recently Discovered Species and Genetic Variants" Diversity 15, no. 6: 699. https://doi.org/10.3390/d15060699
APA StyleDähn, O., Werner, D., Mathieu, B., & Kampen, H. (2023). Development of Conventional Multiplex PCR Assays for the Identification of 21 West Palaearctic Biting Midge Taxa (Diptera: Ceratopogonidae) Belonging to the Culicoides Subgenus Culicoides, including Recently Discovered Species and Genetic Variants. Diversity, 15(6), 699. https://doi.org/10.3390/d15060699