
Morphological Strategies in Ant Communities along Elevational Gradients in Three Mountain Ranges

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
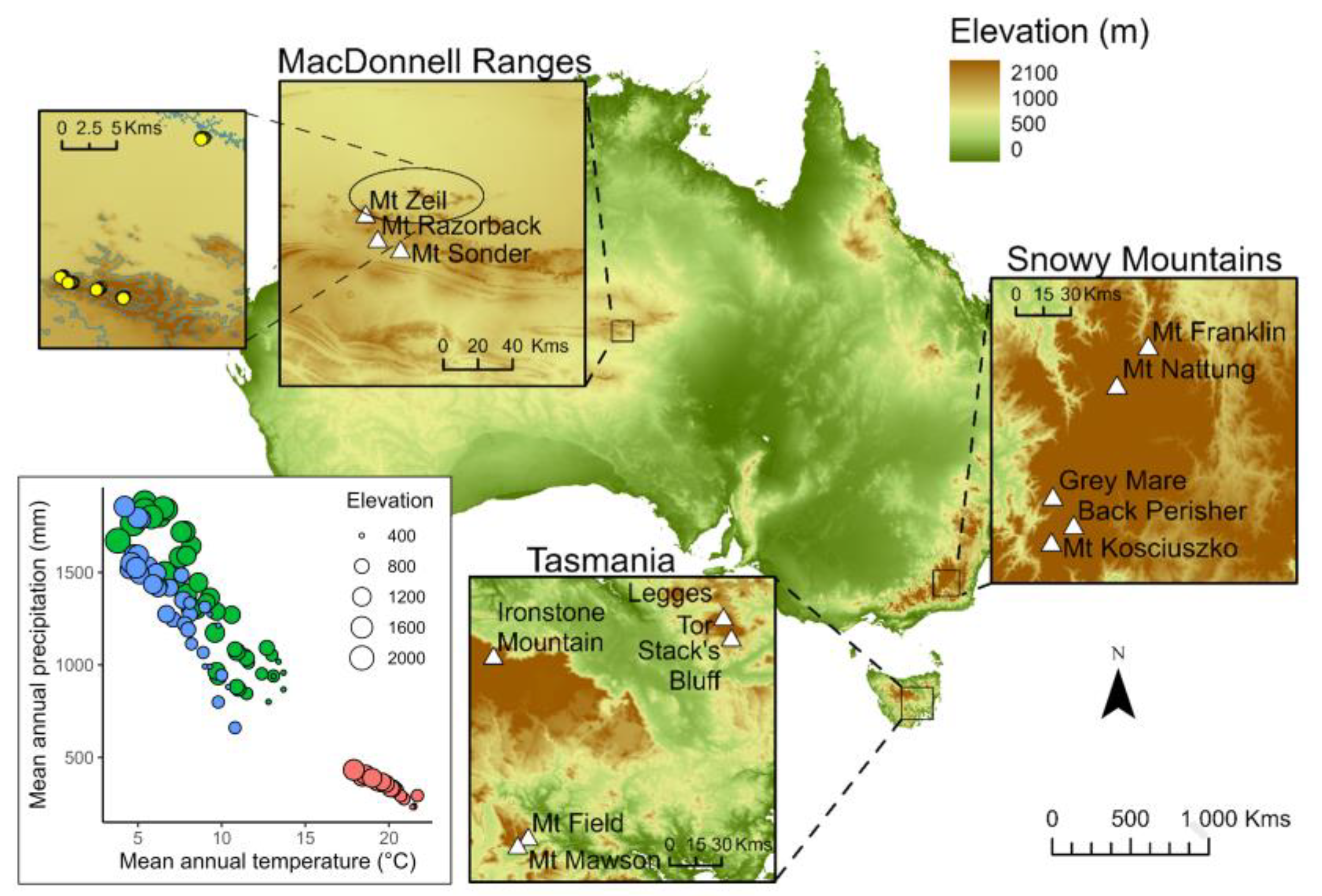
2.1. Study Sites
2.2. Sampling Methods
2.3. Morphological Measurements
2.4. Statistical Analysis
3. Results
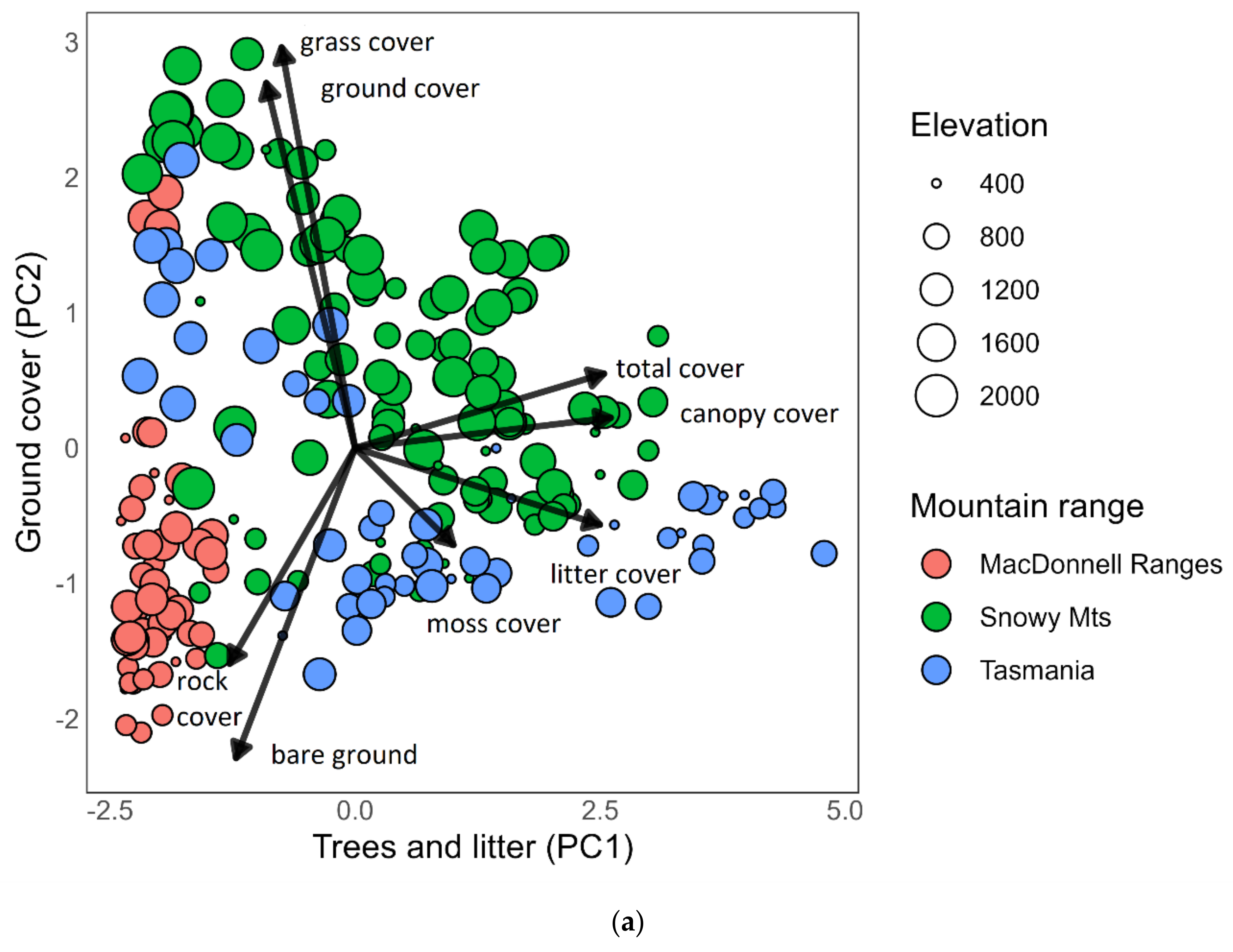
3.1. Microhabitat Composition among Mountain Ranges and Elevations
3.2. Effects of Elevation and Microhabitat on Ant Assemblage Composition
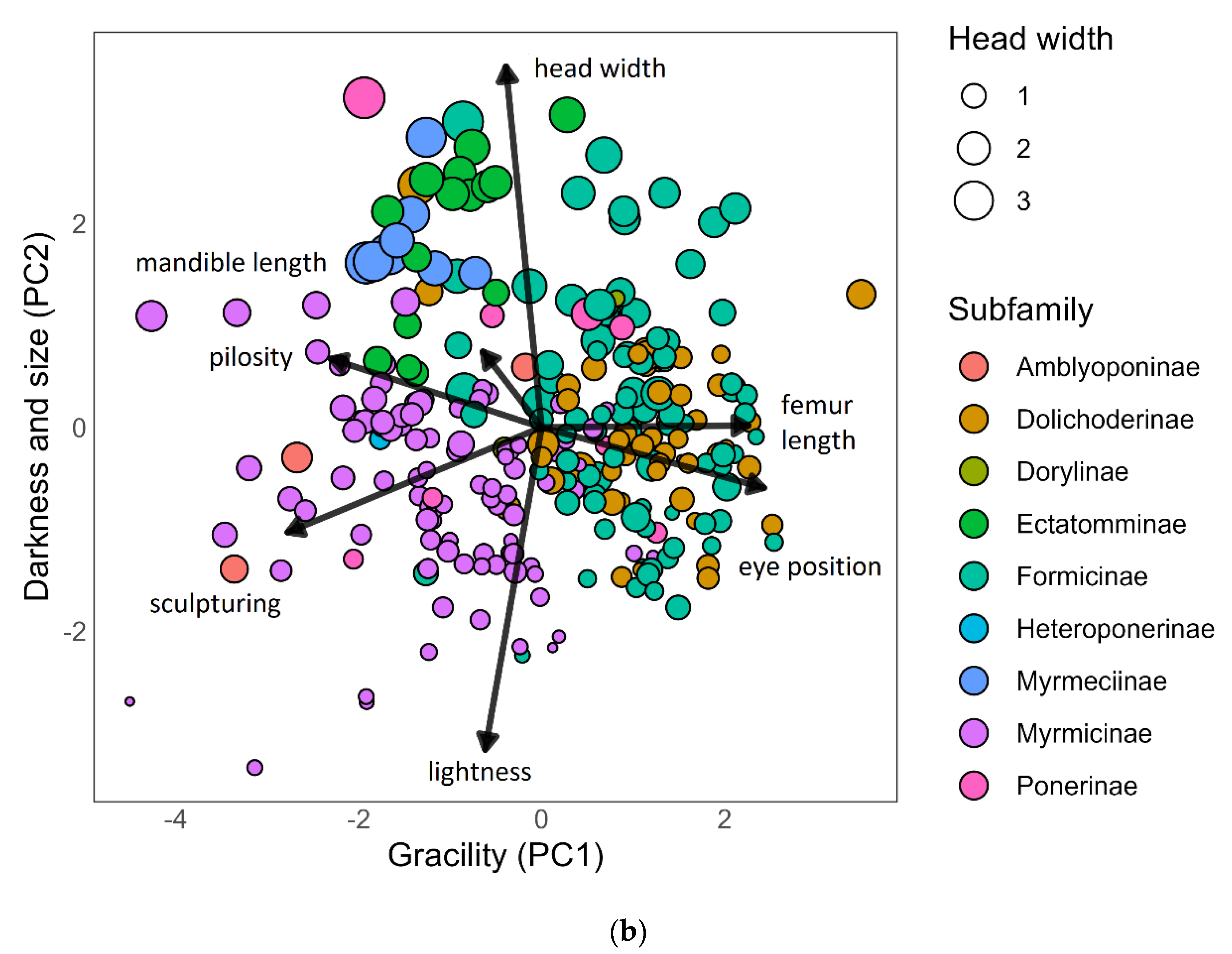
3.3. Morphological Trait Variation among Subfamilies
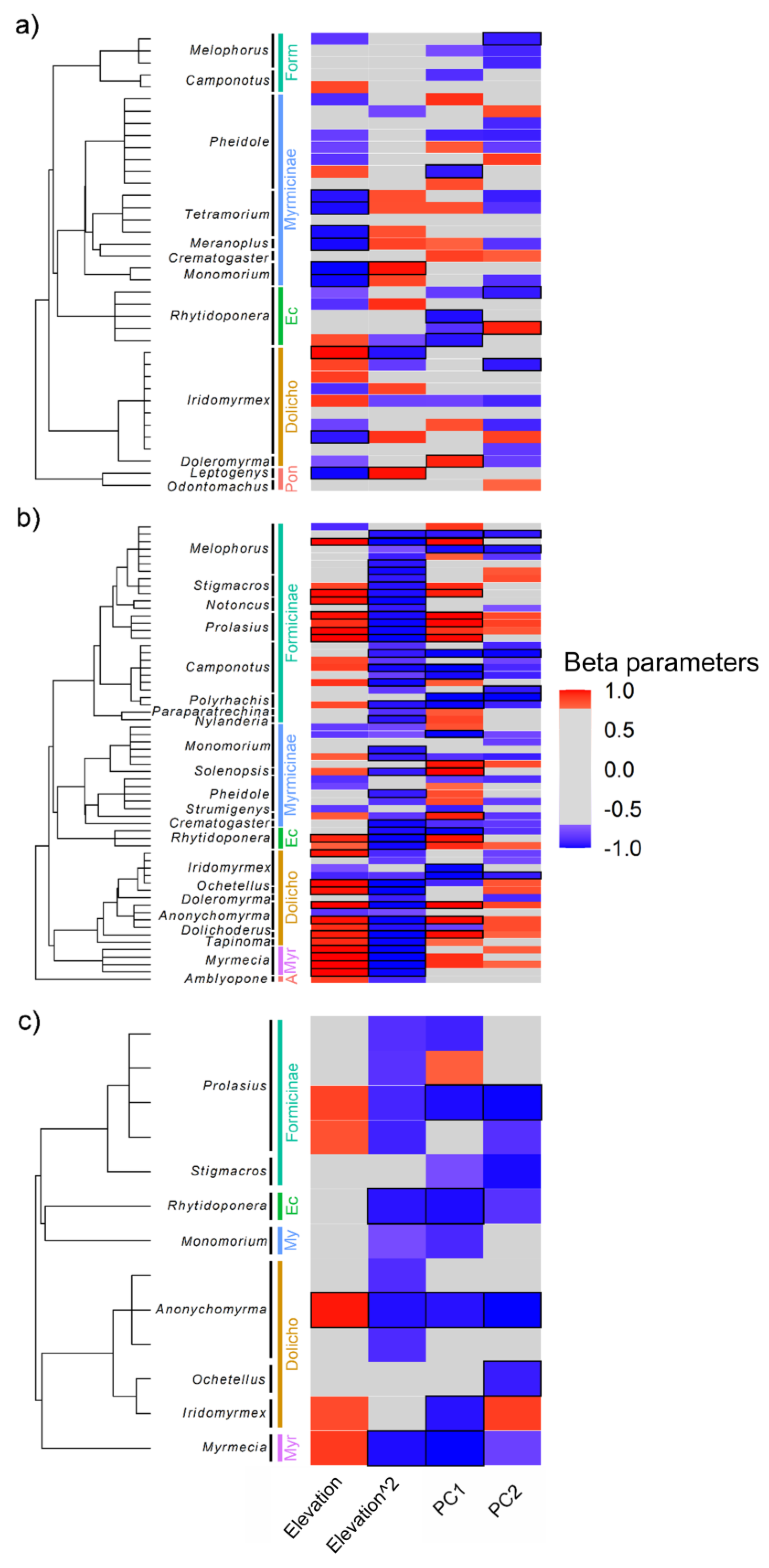
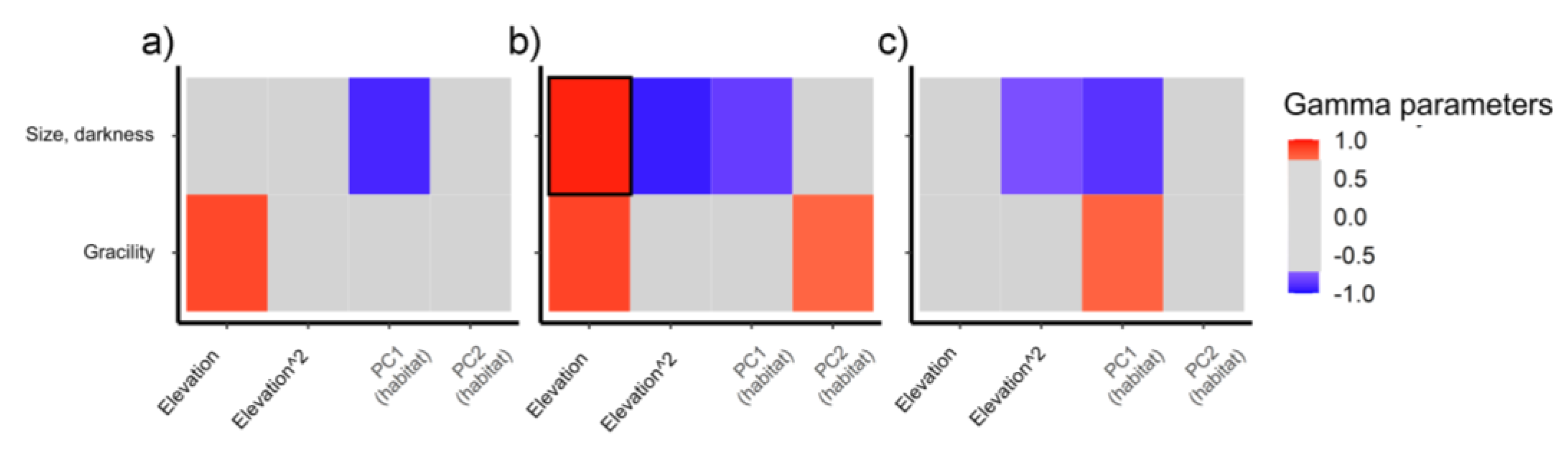
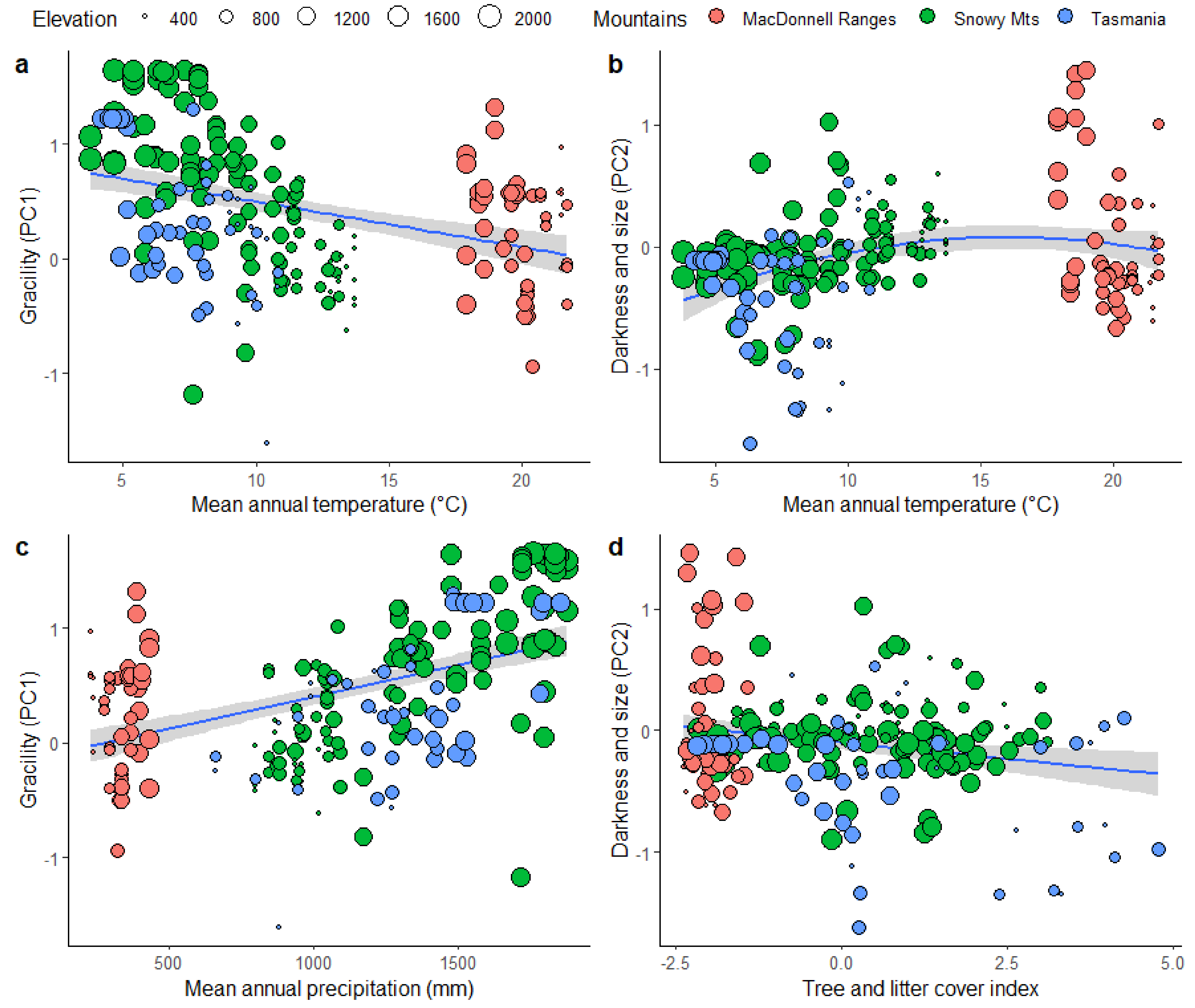
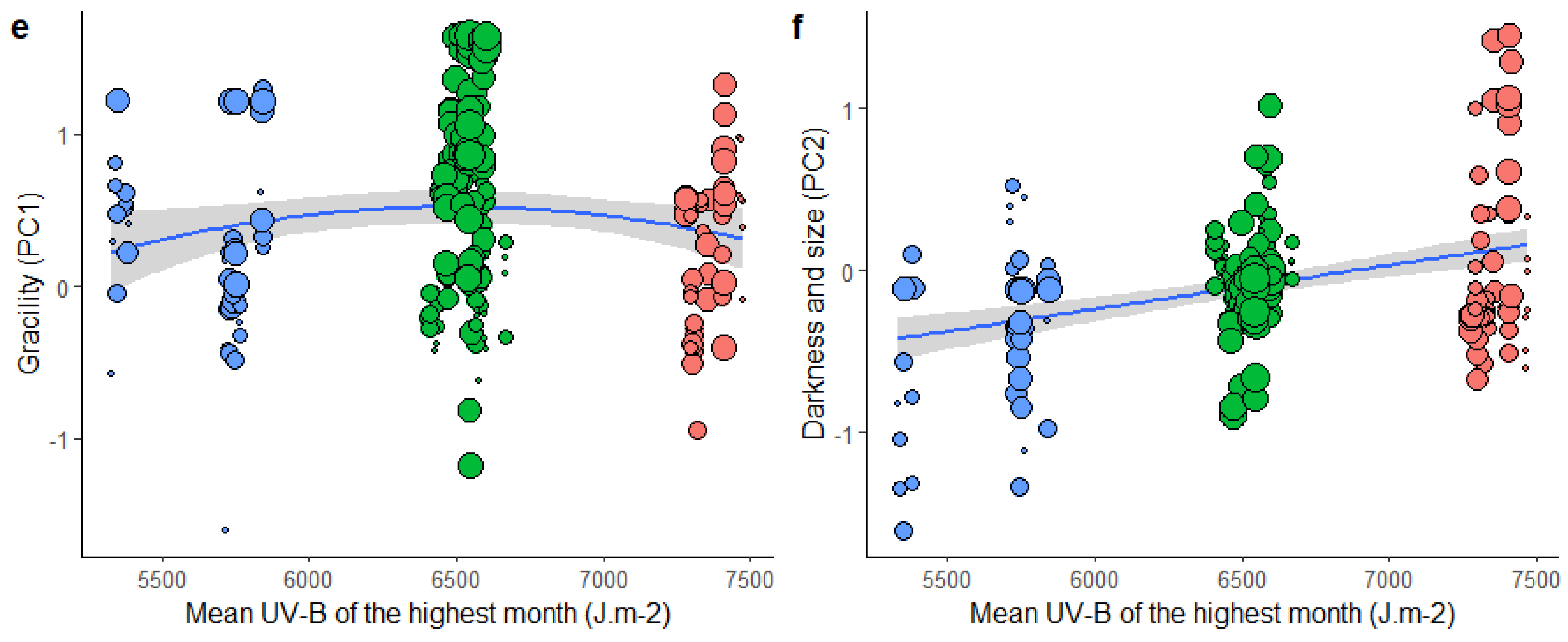
3.4. Effect of Climate and Microhabitat on Morphological Strategies across Mountain Ranges
4. Discussion
4.1. Assemblage Composition along Elevational Gradients
4.2. Changes in Morphological Strategy in Relation to the Environment
4.2.1. Gracility
4.2.2. Size and Darkness
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Gibb, H.; Bishop, T.R.; Leahy, L.; Parr, C.L.; Lessard, J.P.; Sanders, N.J.; Shik, J.Z.; Ibarra-Isassi, J.; Narendra, A.; Dunn, R.R. Ecological strategies of (pl) ants: Towards a world-wide worker economic spectrum for ants. Funct. Ecol. 2023, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Kaspari, M.; Clay, N.A.; Lucas, J.; Yanoviak, S.P.; Kay, A. Thermal adaptation generates a diversity of thermal limits in a rainforest ant community. Glob. Change Biol. 2015, 21, 1092–1102. [Google Scholar] [CrossRef]
- Brown, J.H. Macroecology; University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Chown, S.L.; Gaston, K.J. Body size variation in insects: A macroecological perspective. Biol. Rev. 2010, 85, 139–169. [Google Scholar] [CrossRef]
- Bergmann, C. Uber die verhaltnisse der warmeokonomie der thiere zu ihrer grosse. Gott. Stud. 1847, 1, 595–708. [Google Scholar]
- Rensch, B. Some problems of geographical variation and species-formation. Proc. Linn. Soc. Lond. 1936, 150, 275–285. [Google Scholar] [CrossRef]
- Meiri, S.; Thomas, G.H. The geography of body size–challenges of the interspecific approach. Glob. Ecol. Biogeogr. 2007, 16, 689–693. [Google Scholar] [CrossRef]
- Watt, C.; Mitchell, S.; Salewski, V. Bergmann’s rule; a concept cluster? Oikos 2010, 119, 89–100. [Google Scholar] [CrossRef]
- Huston, M.A.; Wolverton, S. Regulation of animal size by eNPP, Bergmann’s rule and related phenomena. Ecol. Monogr. 2011, 81, 349–405. [Google Scholar] [CrossRef]
- Gibb, H.; Sanders, N.J.; Dunn, R.R.; Arnan, X.; Vasconcelos, H.L.; Donoso, D.A.; Andersen, A.N.; Silva, R.R.; Bishop, T.R.; Gomez, C.; et al. Habitat disturbance selects against both small and large species across varying climates. Ecography 2018, 41, 1184–1193. [Google Scholar] [CrossRef]
- Law, S.J.; Bishop, T.R.; Eggleton, P.; Griffiths, H.; Ashton, L.; Parr, C. Darker ants dominate the canopy: Testing macroecological hypotheses for patterns in colour along a microclimatic gradient. J. Anim. Ecol. 2020, 89, 347–359. [Google Scholar] [CrossRef]
- Delhey, K. A review of Gloger’s rule, an ecogeographical rule of colour: Definitions, interpretations and evidence. Biol. Rev. 2019, 94, 1294–1316. [Google Scholar] [CrossRef] [PubMed]
- Gates, D.M. Biophysical Ecology; Springer: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Bishop, T.R.; Robertson, M.P.; Gibb, H.; Van Rensburg, B.J.; Braschler, B.; Chown, S.L.; Foord, S.H.; Munyai, T.C.; Okey, I.; Tshivhandekano, P.G. Ant assemblages have darker and larger members in cold environments. Glob. Ecol. Biogeogr. 2016, 25, 1489–1499. [Google Scholar] [CrossRef]
- Schweiger, A.H.; Beierkuhnlein, C. Size dependency in colour patterns of Western Palearctic carabids. Ecography 2016, 39, 846–857. [Google Scholar] [CrossRef]
- Moreno Azócar, D.L.; Perotti, M.G.; Bonino, M.F.; Schulte, J.; Abdala, C.S.; Cruz, F.B. Variation in body size and degree of melanism within a lizards clade: Is it driven by latitudinal and climatic gradients? J. Zool. 2015, 295, 243–253. [Google Scholar] [CrossRef]
- Elsen, P.R.; Saxon, E.C.; Simmons, B.A.; Ward, M.; Williams, B.A.; Grantham, H.S.; Kark, S.; Levin, N.; Perez-Hammerle, K.V.; Reside, A.E. Accelerated shifts in terrestrial life zones under rapid climate change. Glob. Change Biol. 2022, 28, 918–935. [Google Scholar] [CrossRef]
- Graham, C.H.; Carnaval, A.C.; Cadena, C.D.; Zamudio, K.R.; Roberts, T.E.; Parra, J.L.; McCain, C.M.; Bowie, R.C.; Moritz, C.; Baines, S.B. The origin and maintenance of montane diversity: Integrating evolutionary and ecological processes. Ecography 2014, 37, 711–719. [Google Scholar] [CrossRef]
- Chown, S.L.; Gaston, K.J. Exploring links between physiology and ecology at macro-scales: The role of respiratory metabolism in insects. Biol. Rev. Camb. Philos. Soc. 1999, 74, 87–120. [Google Scholar] [CrossRef]
- Shah, A.A.; Gill, B.A.; Encalada, A.C.; Flecker, A.S.; Funk, W.C.; Guayasamin, J.M.; Kondratieff, B.C.; Poff, N.L.; Thomas, S.A.; Zamudio, K.R. Climate variability predicts thermal limits of aquatic insects across elevation and latitude. Funct. Ecol. 2017, 31, 2118–2127. [Google Scholar] [CrossRef]
- DeMarche, M.L.; Doak, D.F.; Morris, W.F. Incorporating local adaptation into forecasts of species’ distribution and abundance under climate change. Glob. Change Biol. 2019, 25, 775–793. [Google Scholar] [CrossRef] [PubMed]
- Midolo, G.; De Frenne, P.; Hölzel, N.; Wellstein, C. Global patterns of intraspecific leaf trait responses to elevation. Glob. Change Biol. 2019, 25, 2485–2498. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Valverde, A.; Lobo, J.M. Determinants of local spider (Araneidae and Thomisidae) species richness on a regional scale: Climate and altitude vs. habitat structure. Ecol. Entomol. 2007, 32, 113–122. [Google Scholar] [CrossRef]
- Neel, L.K.; Logan, M.L.; Nicholson, D.J.; Miller, C.; Chung, A.K.; Maayan, I.; Degon, Z.; DuBois, M.; Curlis, J.D.; Taylor, Q. Habitat structure mediates vulnerability to climate change through its effects on thermoregulatory behavior. Biotropica 2021, 53, 1121–1133. [Google Scholar] [CrossRef]
- Gibb, H. The effect of a dominant ant, Iridomyrmex purpureus, on resource use by ant assemblages depends on microhabitat and resource type. Austral Ecol. 2005, 30, 856–867. [Google Scholar] [CrossRef]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef]
- Burger, J.R.; Hou, C.; Brown, J.H. Toward a metabolic theory of life history. Proc. Natl. Acad. Sci. USA 2019, 116, 26653–26661. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Wikelski, M. The physiology/life-history nexus. Trends Ecol. Evol. 2002, 17, 462–468. [Google Scholar] [CrossRef]
- Andersen, A.N. A classification of Australian ant communities, based on functional groups which parallel plant life-forms in relation to stress and disturbance. J. Biogeogr. 1995, 22, 15–29. [Google Scholar] [CrossRef]
- Wilson, E.O. The little things that run the world*(The Importance and Conservation of Invertebrates). Conserv. Biol. 1987, 1, 344–346. [Google Scholar] [CrossRef]
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The little things that run the world revisited: A review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol. News 2012, 17, 133–146. [Google Scholar]
- King, J.R.; Warren, R.J.; Bradford, M.A. Social insects dominate eastern US temperate hardwood forest macroinvertebrate communities in warmer regions. PLoS ONE 2013, 8, e75843. [Google Scholar] [CrossRef]
- Schultheiss, P.; Nooten, S.S.; Wang, R.; Wong, M.K.; Brassard, F.; Guénard, B. The abundance, biomass, and distribution of ants on Earth. Proc. Natl. Acad. Sci. USA 2022, 119, e2201550119. [Google Scholar] [CrossRef] [PubMed]
- Gibb, H.; Parr, C.L. How does habitat complexity affect ant foraging success? A test of functional responses on three continents. Oecologia 2010, 164, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.; Wehner, R. Leg allometry in ants: Extreme long-leggedness in thermophilic species. Arthropod Struct. Dev. 2012, 41, 71–77. [Google Scholar] [CrossRef]
- Wiescher, P.T.; Pearce-Duvet, J.M.C.; Feener, D.H. Assembling an ant community: Species functional traits reflect environmental filtering. Oecologia 2012, 169, 1063–1074. [Google Scholar] [CrossRef]
- Vincent, J.F.; Wegst, U.G. Design and mechanical properties of insect cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef]
- Buxton, J.T.; Robert, K.A.; Marshall, A.T.; Dutka, T.L.; Gibb, H. A cross-species test of the function of cuticular traits in ants (Hymenoptera: Formicidae). Myrmecol. News 2021, 31, 31–46. [Google Scholar] [CrossRef]
- Silva, R.R.; Brandão, C.R.F. Morphological patterns and community organization in leaf-litter ant assemblages. Ecol. Monogr. 2010, 80, 107–124. [Google Scholar] [CrossRef]
- Sosiak, C.E.; Barden, P. Multidimensional trait morphology predicts ecology across ant lineages. Funct. Ecol. 2021, 35, 139–152. [Google Scholar] [CrossRef]
- Gibb, H.; Parr, C.L. Does Structural Complexity Determine the Morphology of Assemblages? An Experimental Test on Three Continents. PLoS ONE 2013, 8, e0064005. [Google Scholar] [CrossRef]
- Parr, C.L.; Dunn, R.R.; Sanders, N.J.; Weiser, M.D.; Photakis, M.; Bishop, T.R.; Fitzpatrick, M.C.; Arnan, X.; Baccaro, F.; Brandao, C.R.F.; et al. GlobalAnts: A new database on the geography of ant traits (Hymenoptera: Formicidae). Insect Conserv. Diver. 2017, 10, 5–20. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing, Version 3.03; R Foundation for Statistical Computing: Vienna, Austria, 2017.
- Tikhonov, G.; Opedal, Ø.H.; Abrego, N.; Lehikoinen, A.; de Jonge, M.M.; Oksanen, J.; Ovaskainen, O. Joint species distribution modelling with the R-package Hmsc. Methods Ecol. Evol. 2020, 11, 442–447. [Google Scholar] [CrossRef]
- Ovaskainen, O.; Abrego, N. Joint Species Distribution Modelling: With Applications in R; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Ovaskainen, O.; Tikhonov, G.; Norberg, A.; Guillaume Blanchet, F.; Duan, L.; Dunson, D.; Roslin, T.; Abrego, N. How to make more out of community data? A conceptual framework and its implementation as models and software. Ecol. Lett. 2017, 20, 561–576. [Google Scholar] [CrossRef]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.S.; Nemergut, D.; Seyednasrollah, B.; Turner, P.J.; Zhang, S. Generalized joint attribute modeling for biodiversity analysis: Median-zero, multivariate, multifarious data. Ecol. Monogr. 2017, 87, 34–56. [Google Scholar] [CrossRef]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-6. 2014. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 1 January 2024).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Beckmann, M.; Václavík, T.; Manceur, A.M.; Šprtová, L.; von Wehrden, H.; Welk, E.; Cord, A.F. gl UV: A global UV-B radiation data set for macroecological studies. Methods Ecol. Evol. 2014, 5, 372–383. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference, R Package Version 1.0.0; R Foundation for Statistical Computing: Vienna, Austria, 2011. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 1 January 2024).
- Andersen, A.N. Ant diversity in arid Australia: A systematic overview. Mem. Am. Entomol. Soc. 2007, 80, 20. [Google Scholar]
- Dunn, R.R.; Agosti, D.; Andersen, A.N.; Arnan, X.; Bruhl, C.A.; Cerda, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Kühsel, S.; Brückner, A.; Schmelzle, S.; Heethoff, M.; Blüthgen, N. Surface area–volume ratios in insects. Insect Sci. 2017, 24, 829–841. [Google Scholar] [CrossRef]
- Wolpert, A. Heat transfer analysis of factors affecting plant leaf temperature. Significance of leaf hair. Plant Physiol. 1962, 37, 113. [Google Scholar] [CrossRef] [PubMed]
- Wuenscher, J.E. The effect of leaf hairs of Verbascum thapsus on leaf energy exchange. New Phytol. 1970, 69, 65–73. [Google Scholar] [CrossRef]
- Casey, T.M.; Hegel, J.R. Caterpillar setae: Insulation for an ectotherm. Science 1981, 214, 1131–1133. [Google Scholar] [CrossRef]
- Kevan, P.G.; Jensen, T.S.; Shorthouse, J.D. Body temperatures and behavioral thermoregulation of high arctic woolly-bear caterpillars and pupae (Gynaephora rossii, Lymantriidae: Lepidoptera) and the importance of sunshine. Arct. Alp. Res. 1982, 14, 125–136. [Google Scholar] [CrossRef]
- Seago, A.E.; Brady, P.; Vigneron, J.-P.; Schultz, T.D. Gold bugs and beyond: A review of iridescence and structural colour mechanisms in beetles (Coleoptera). J. R. Soc. Interface 2009, 6, S165–S184. [Google Scholar] [CrossRef]
- Shi, N.N.; Tsai, C.-C.; Camino, F.; Bernard, G.D.; Yu, N.; Wehner, R. Keeping cool: Enhanced optical reflection and radiative heat dissipation in Saharan silver ants. Science 2015, 349, 298–301. [Google Scholar] [CrossRef]
- Willot, Q.; Simonis, P.; Vigneron, J.-P.; Aron, S. Total internal reflection accounts for the bright color of the Saharan silver ant. PLoS ONE 2016, 11, e0152325. [Google Scholar] [CrossRef]
- Schultz, T.D.; Hadley, N.F. Structural colors of tiger beetles and their role in heat transfer through the integument. Physiol. Zool. 1987, 60, 737–745. [Google Scholar] [CrossRef]
- Matute, D.R.; Harris, A. The influence of abdominal pigmentation on desiccation and ultraviolet resistance in two species of Drosophila. Evolution 2013, 67, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- Schofield, S.F.; Bishop, T.R.; Parr, C.L. Morphological characteristics of ant assemblages (Hymenoptera: Formicidae) differ among contrasting biomes. Myrmecol. News 2016, 23, 129–137. [Google Scholar]
- Idec, J.H.; Bishop, T.R.; Fisher, B.L. Using computer vision to understand the global biogeography of ant color. Ecography 2023, 2023, e06279. [Google Scholar] [CrossRef]
- Trullas, S.C.; van Wyk, J.H.; Spotila, J.R. Thermal melanism in ectotherms. J. Therm. Biol. 2007, 32, 235–245. [Google Scholar] [CrossRef]
- Watt, W.B. Adaptive significance of pigment polymorphisms in Colias butterflies. I. Variation of melanin pigment in relation to thermoregulation. Evolution 1968, 437–458. [Google Scholar] [CrossRef]
- Feild, T.S.; Brodribb, T. Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath. Oecologia 2001, 127, 314–320. [Google Scholar] [CrossRef]
- Green, K.; Pickering, C.M. The decline of snowpatches in the Snowy Mountains of Australia: Importance of climate warming, variable snow, and wind. Arct. Antarct. Alp. Res. 2009, 41, 212–218. [Google Scholar] [CrossRef]
- Woods, H.A.; Dillon, M.E.; Pincebourde, S. The roles of microclimatic diversity and of behavior in mediating the responses of ectotherms to climate change. J. Therm. Biol. 2015, 54, 86–97. [Google Scholar] [CrossRef]
- Ma, C.-S.; Ma, G.; Pincebourde, S. Survive a warming climate: Insect responses to extreme high temperatures. Annu. Rev. Entomol. 2021, 66, 163–184. [Google Scholar] [CrossRef]
- Kaspari, M.; Weiser, M. The size–grain hypothesis and interspecific scaling in ants. Funct. Ecol. 1999, 13, 530–538. [Google Scholar] [CrossRef]
- Andersen, A.N. The use of ant communities to evaluate change in Australian terrestrial ecosystems: A review and a recipe. Proc. Ecol. Soc. Aust. 1990, 16, 347–357. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Best Models | Intercept | poly(MAP,2) | poly(MAT,2) | poly(UVB,2) | MAP | MAT | UV-B | Habitat PC1 | Habitat PC2 | df | logLik | AICc | Delta | Weight |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PC1 “gracility” | ||||||||||||||
| Model 1 | 0.43 | + | 0.27 | −0.60 | 8.00 | −128.42 | 273.5 | 0.00 | 0.064 | |||||
| Model 2 | 0.43 | + | −0.93 | 7.00 | −130.14 | 274.8 | 1.28 | 0.034 | ||||||
| Model 3 | 0.43 | + | 0.26 | −0.63 | 0.02 | 9.00 | −128.26 | 275.4 | 1.85 | 0.025 | ||||
| PC2 “darkness and size” | ||||||||||||||
| Model 1 | −0.09 | + | 0.21 | −0.14 | 8.00 | −99.72 | 216.1 | 0.00 | 0.079 | |||||
| Model 2 | −0.09 | + | −0.19 | 0.27 | −0.12 | 9.00 | −98.69 | 216.2 | 0.11 | 0.074 | ||||
| Source | χ2 | df | p-Value |
|---|---|---|---|
| PC1 “gracility” | |||
| Mean Annual Temperature (MAT) | 10.3 | 1 | 0.0013 |
| Mean Annual Precipitation (MAP) | 3.8 | 1 | 0.0516 |
| UVB (Polynomial, 2) | 67.5 | 2 | <0.0001 |
| PC2 “darkness and size” | |||
| Mean Annual Temperature (MAT) | 24.1 | 2 | <0.0001 |
| PC1 (trees and litter cover) | 11.1 | 1 | 0.0009 |
| UVB | 11.8 | 1 | 0.0006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibb, H.; Contos, P.; Photakis, M.; Okey, I.; Dunn, R.R.; Sanders, N.J.; Jones, M.M. Morphological Strategies in Ant Communities along Elevational Gradients in Three Mountain Ranges. Diversity 2024, 16, 48. https://doi.org/10.3390/d16010048
Gibb H, Contos P, Photakis M, Okey I, Dunn RR, Sanders NJ, Jones MM. Morphological Strategies in Ant Communities along Elevational Gradients in Three Mountain Ranges. Diversity. 2024; 16(1):48. https://doi.org/10.3390/d16010048
Chicago/Turabian StyleGibb, Heloise, Peter Contos, Manoli Photakis, Iona Okey, Robert R. Dunn, Nathan J. Sanders, and Mirkka M. Jones. 2024. "Morphological Strategies in Ant Communities along Elevational Gradients in Three Mountain Ranges" Diversity 16, no. 1: 48. https://doi.org/10.3390/d16010048
APA StyleGibb, H., Contos, P., Photakis, M., Okey, I., Dunn, R. R., Sanders, N. J., & Jones, M. M. (2024). Morphological Strategies in Ant Communities along Elevational Gradients in Three Mountain Ranges. Diversity, 16(1), 48. https://doi.org/10.3390/d16010048