
Roles of Bacterial Mechanosensitive Channels in Infection and Antibiotic Susceptibility


Abstract
:1. Introduction
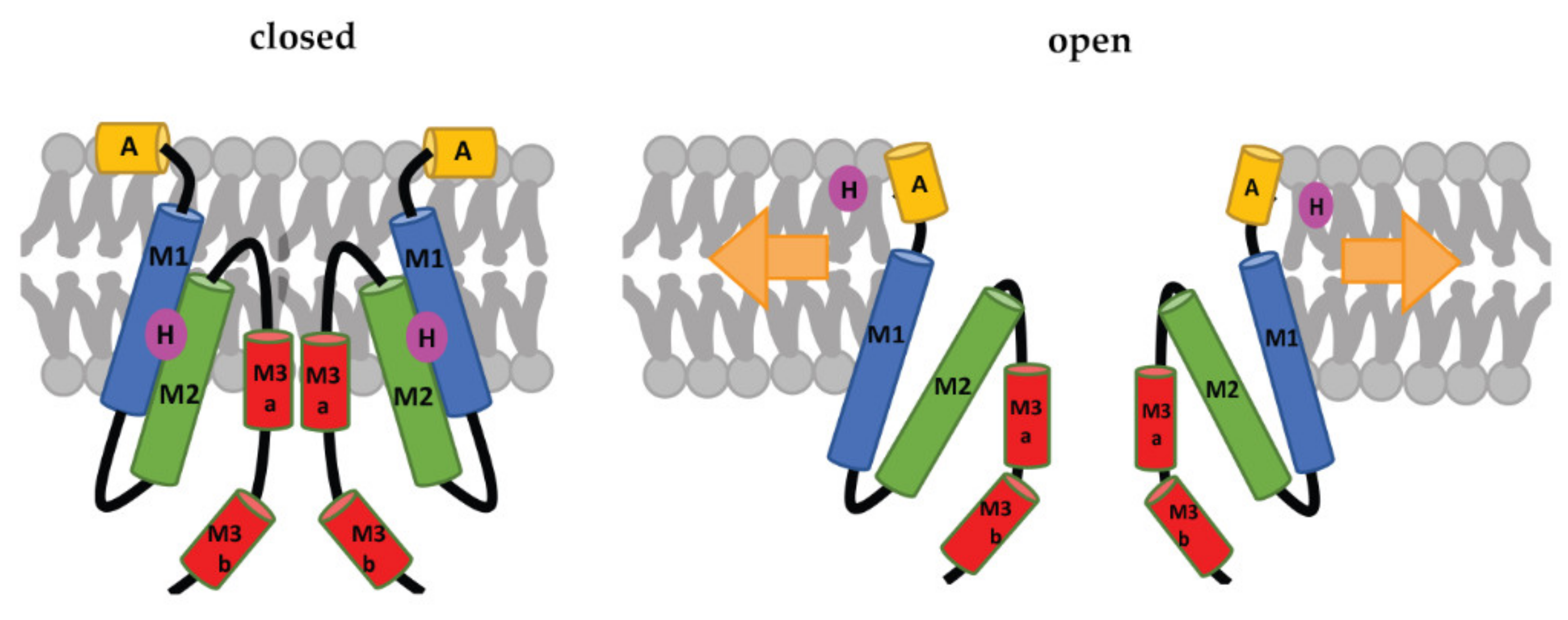
2. Structure and Gating of Bacterial Mechanosensitive Channels
2.1. Structural Determinant of MscL and MscS Gating
2.2. Importance of Membrane Lipids for the Gating Mechanism
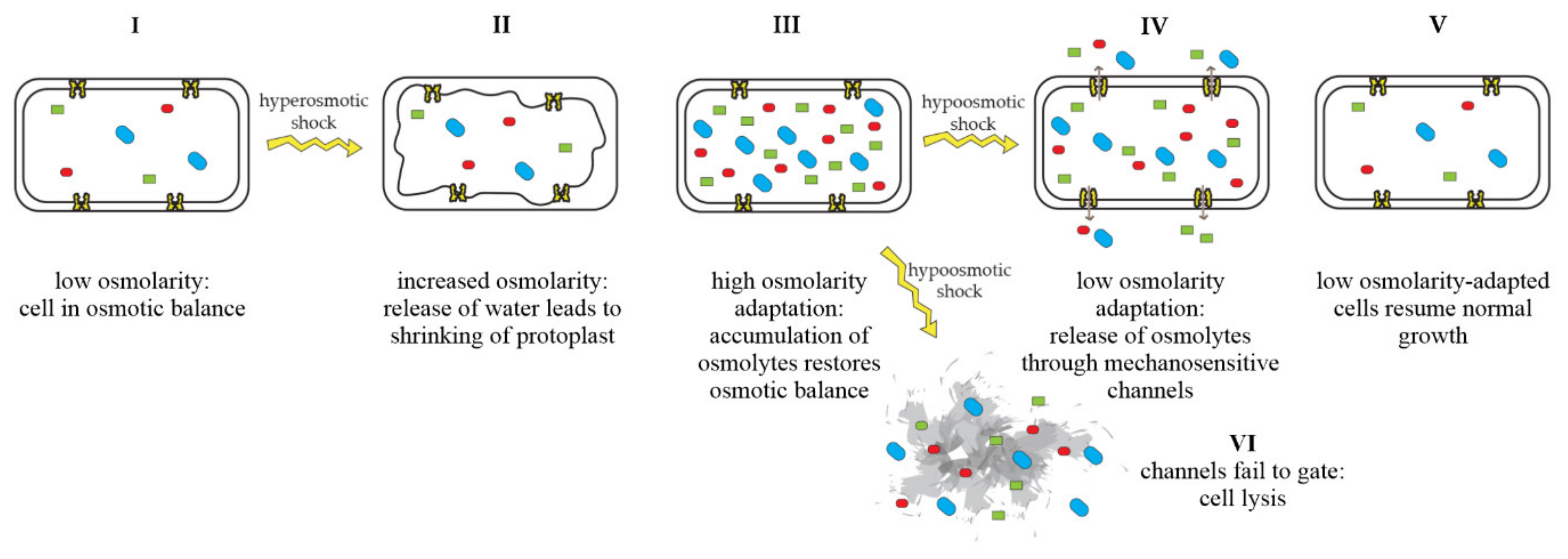
3. Role of Mechanosensitive Channels during Infection
3.1. Transition between Host and Environment
3.2. The Urinary Tract as an Osmotically Challenging Environment in the Human Body
4. Impact on Antibiotic Susceptibility
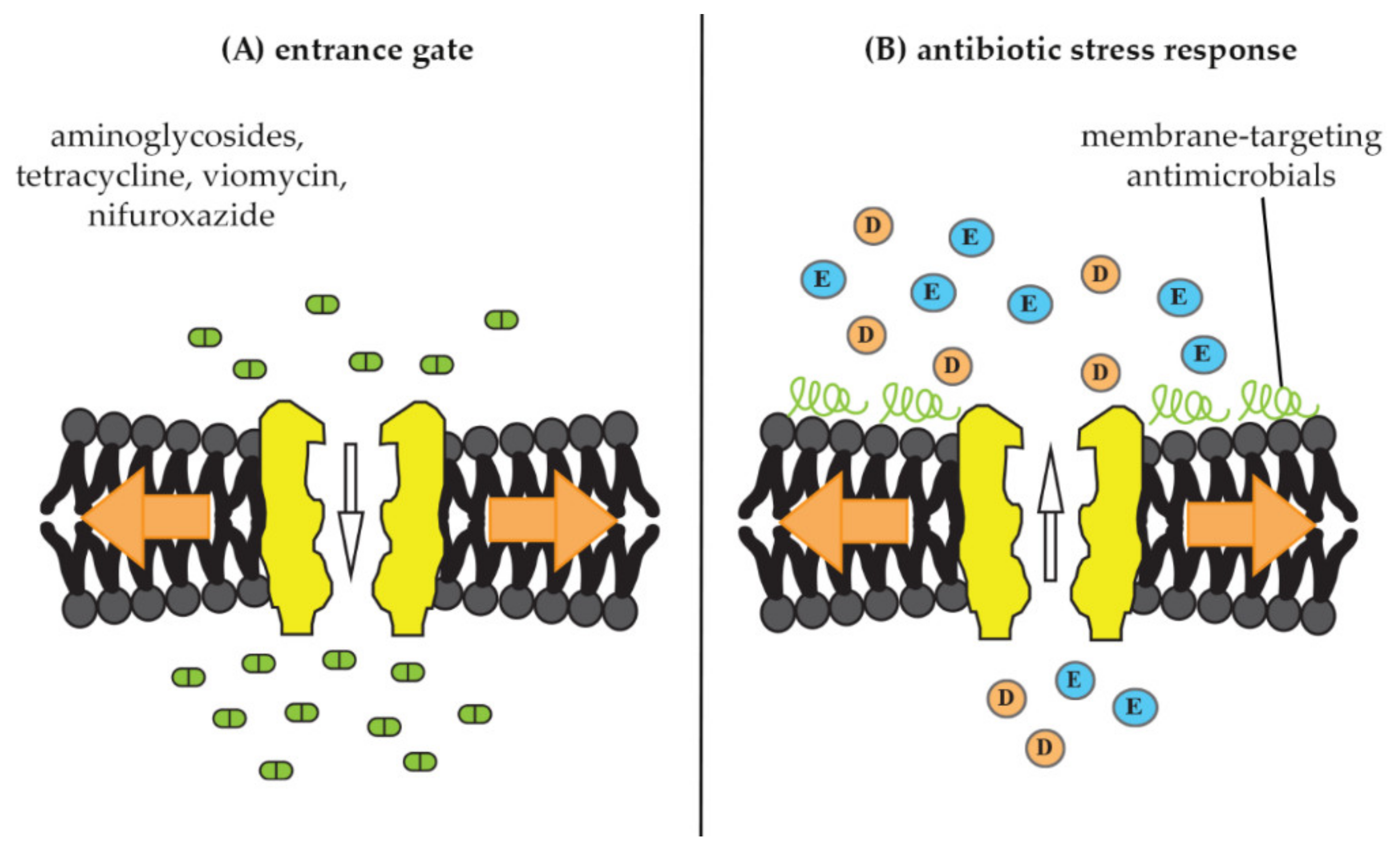
4.1. Mechanosensitive Channels as Antibiotic Entry Point
4.2. Mechanosensitive-Channel Activation as Antibiotic-Stress Response
4.3. Osmolarity and Host-Defense Peptides
5. Mechanosensitive Channels as Novel Antimicrobial-Drug Targets
5.1. Compounds Directly Targeting Mechanosensitive Channels
5.2. Compounds Indirectly Modulating Channel Gating
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pivetti, C.D.; Yen, M.-R.; Miller, S.; Busch, W.; Tseng, Y.-H.; Booth, I.R.; Saier, M.H. Two Families of Mechanosensitive Channel Proteins. Microbiol. Mol. Biol. Rev. 2003, 67, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Booth, I.R.; Miller, S.; Müller, A.; Lehtovirta-Morley, L. The Evolution of Bacterial Mechanosensitive Channels. Cell Calcium 2015, 57, 140–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, C.; Martinac, B.; Sukharev, S. Mechanosensitive Channels in Microbes. Annu. Rev. Microbiol. 2010, 64, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Árnadóttir, J.; Chalfie, M. Eukaryotic Mechanosensitive Channels. Annu. Rev. Biophys. 2010, 39, 111–137. [Google Scholar] [CrossRef]
- Levina, N. Protection of Escherichia coli Cells against Extreme Turgor by Activation of MscS and MscL Mechanosensitive Channels: Identification of Genes Required for MscS Activity. EMBO J. 1999, 18, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Booth, I.R.; Blount, P. The MscS and MscL Families of Mechanosensitive Channels Act as Microbial Emergency Release Valves. J. Bacteriol. 2012, 194, 4802–4809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csonka, L.N. Physiological and Genetic Responses of Bacteria to Osmotic Stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef]
- Bougouffa, S.; Radovanovic, A.; Essack, M.; Bajic, V.B. DEOP: A Database on Osmoprotectants and Associated Pathways. Database 2014, 2014, bau100. [Google Scholar] [CrossRef]
- Hoffmann, T.; Bremer, E. Protection of Bacillus subtilis against Cold Stress via Compatible-Solute Acquisition. J. Bacteriol. 2011, 193, 1552–1562. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.; Boiangiu, C.; Moses, S.; Bremer, E. Responses of Bacillus subtilis to Hypotonic Challenges: Physiological Contributions of Mechanosensitive Channels to Cellular Survival. Appl. Environ. Microbiol. 2008, 74, 2454–2460. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Chiriac, A.I.; Otto, A.; Zweytick, D.; May, C.; Schumacher, C.; Gust, R.; Albada, H.B.; Penkova, M.; Krämer, U.; et al. Small Cationic Antimicrobial Peptides Delocalize Peripheral Membrane Proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E1409–E1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajouz, B.; Berrier, C.; Garrigues, A.; Besnard, M.; Ghazi, A. Release of Thioredoxin via the Mechanosensitive Channel MscL during Osmotic Downshock of Escherichia coli Cells. J. Biol. Chem. 1998, 273, 26670–26674. [Google Scholar] [CrossRef] [Green Version]
- Kakuda, T.; Koide, Y.; Sakamoto, A.; Takai, S. Characterization of Two Putative Mechanosensitive Channel Proteins of Campylobacter Jejuni Involved in Protection against Osmotic Downshock. Vet. Microbiol. 2012, 160, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.R.; Morgan, E.; Peters, S.E.; Pleasance, S.J.; Hudson, D.L.; Davies, H.M.; Wang, J.; van Diemen, P.M.; Buckley, A.M.; Bowen, A.J.; et al. Comprehensive Assignment of Roles for Salmonella Typhimurium Genes in Intestinal Colonization of Food-Producing Animals. PLoS Genet. 2013, 9, e1003456. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, X.; Lu, P.; Ni, C.; Li, Y.; van der Veen, S. Identification and Characterization of the Neisseria gonorrhoeae MscS-Like Mechanosensitive Channel. Infect. Immun. 2018, 86, e00090-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, D.R.; Dewan, K.K.; Patel, T.; Wastella, C.M.; Ning, G.; Kirimanjeswara, G.S. A Single Mechanosensitive Channel Protects Francisella tularensis Subsp. Holarctica from Hypoosmotic Shock and Promotes Survival in the Aquatic Environment. Appl. Environ. Microbiol. 2018, 84, e02203-17. [Google Scholar] [CrossRef] [Green Version]
- Asogwa, M.; Miller, S.; Spano, S.; Stevens, M. Investigating the Role of the Bacterial Mechanosensitive Channel YnaI in Salmonella Pathogenesis. Access Microbiol. 2019, 1, 500. [Google Scholar] [CrossRef]
- Kouwen, T.R.H.M.; Trip, E.N.; Denham, E.L.; Sibbald, M.J.J.B.; Dubois, J.-Y.F.; van Dijl, J.M. The Large Mechanosensitive Channel MscL Determines Bacterial Susceptibility to the Bacteriocin Sublancin 168. Antimicrob. Agents Chemother. 2009, 53, 4702–4711. [Google Scholar] [CrossRef] [Green Version]
- Iscla, I.; Wray, R.; Wei, S.; Posner, B.; Blount, P. Streptomycin Potency Is Dependent on MscL Channel Expression. Nat. Commun. 2014, 5, 4891. [Google Scholar] [CrossRef] [Green Version]
- Wray, R.; Iscla, I.; Gao, Y.; Li, H.; Wang, J.; Blount, P. Dihydrostreptomycin Directly Binds to, Modulates, and Passes through the MscL Channel Pore. PLoS Biol. 2016, 14, e1002473. [Google Scholar] [CrossRef]
- Wenzel, M.; Senges, C.H.R.; Zhang, J.; Suleman, S.; Nguyen, M.; Kumar, P.; Chiriac, A.I.; Stepanek, J.J.; Raatschen, N.; May, C.; et al. Antimicrobial Peptides from the Aurein Family Form Ion-Selective Pores in Bacillus subtilis. ChemBioChem 2015, 16, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Booth, I.R.; Edwards, M.D.; Black, S.; Schumann, U.; Miller, S. Mechanosensitive Channels in Bacteria: Signs of Closure? Nat. Rev. Microbiol. 2007, 5, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Haswell, E.S.; Phillips, R.; Rees, D.C. Mechanosensitive Channels: What Can They Do and How Do They Do It? Structure 2011, 19, 1356–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y. Ionic Regulation of MscK, a Mechanosensitive Channel from Escherichia coli. EMBO J. 2002, 21, 5323–5330. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.; Spencer, R.H.; Lee, A.T.; Barclay, M.T.; Rees, D.C. Structure of the MscL Homolog from Mycobacterium tuberculosis: A Gated Mechanosensitive Ion Channel. Science 1998, 282, 2220–2226. [Google Scholar] [CrossRef] [Green Version]
- Bass, R.B.; Strop, P.; Barclay, M.; Rees, D.C. Crystal Structure of Escherichia coli MscS, a Voltage-Modulated and Mechanosensitive Channel. Science 2002, 298, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Booth, I.R. Bacterial Mechanosensitive Channels: Progress towards an Understanding of Their Roles in Cell Physiology. Curr. Opin. Microbiol. 2014, 18, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, Y.; Hashimoto, K.; Sawada, Y.; Sokabe, M.; Kawasaki, H.; Martinac, B. Corynebacterium Glutamicum Mechanosensitive Channels: Towards Unpuzzling “Glutamate Efflux” for Amino Acid Production. Biophys. Rev. 2018, 10, 1359–1369. [Google Scholar] [CrossRef]
- Kawasaki, H.; Martinac, B. Mechanosensitive Channels of Corynebacterium Glutamicum Functioning as Exporters of L-Glutamate and Other Valuable Metabolites. Curr. Opin. Chem. Biol. 2020, 59, 77–83. [Google Scholar] [CrossRef]
- Cox, C.D.; Bavi, N.; Martinac, B. Bacterial Mechanosensors. Annu. Rev. Physiol. 2018, 80, 71–93. [Google Scholar] [CrossRef]
- Rasmussen, T. How Do Mechanosensitive Channels Sense Membrane Tension? Biochem. Soc. Trans. 2016, 44, 1019–1025. [Google Scholar] [CrossRef]
- Iscla, I.; Blount, P. Sensing and Responding to Membrane Tension: The Bacterial MscL Channel as a Model System. Biophys. J. 2012, 103, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbacher, S.; Bass, R.; Strop, P.; Rees, D.C. Structures of the Prokaryotic Mechanosensitive Channels MscL and MscS. In Current Topics in Membranes; Benos, D.J., Simon, S.A., Eds.; Academic Press: San Diego, CA, USA, 2007; Volume 58, pp. 1–24. [Google Scholar] [CrossRef]
- Reddy, B.; Bavi, N.; Lu, A.; Park, Y.; Perozo, E. Molecular Basis of Force-from-Lipids Gating in the Mechanosensitive Channel MscS. eLife 2019, 8, e50486. [Google Scholar] [CrossRef] [PubMed]
- Blount, P.; Iscla, I. Life with Bacterial Mechanosensitive Channels, from Discovery to Physiology to Pharmacological Target. Microbiol. Mol. Biol. Rev. 2020, 84, e00055-19. [Google Scholar] [CrossRef] [PubMed]
- Kapsalis, C.; Wang, B.; el Mkami, H.; Pitt, S.J.; Schnell, J.R.; Smith, T.K.; Lippiat, J.D.; Bode, B.E.; Pliotas, C. Allosteric Activation of an Ion Channel Triggered by Modification of Mechanosensitive Nano-Pockets. Nat. Commun. 2019, 10, 4619. [Google Scholar] [CrossRef] [Green Version]
- Kapsalis, C.; Ma, Y.; Bode, B.E.; Pliotas, C. In-Lipid Structure of Pressure-Sensitive Domains Hints Mechanosensitive Channel Functional Diversity. Biophys. J. 2020, 119, 448–459. [Google Scholar] [CrossRef]
- Naismith, J.H.; Booth, I.R. Bacterial Mechanosensitive Channels—MscS: Evolution’s Solution to Creating Sensitivity in Function. Annu. Rev. Biophys. 2012, 41, 157–177. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.; Park, S.; Reddy, B.; Fei, J.; Perozo, E. Using Fluorescence Microscopy to Characterize the Role of Mechanosensation in Cell Division. Biophys. J. 2020, 118, 529a. [Google Scholar] [CrossRef]
- Gamini, R.; Sotomayor, M.; Chipot, C.; Schulten, K. Cytoplasmic Domain Filter Function in the Mechanosensitive Channel of Small Conductance. Biophys. J. 2011, 101, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Rowe, I.; Anishkin, A.; Kamaraju, K.; Yoshimura, K.; Sukharev, S. The Cytoplasmic Cage Domain of the Mechanosensitive Channel MscS Is a Sensor of Macromolecular Crowding. J. Gen. Physiol. 2014, 143, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Cox, C.D.; Bavi, N.; Martinac, B. Biophysical Principles of Ion-Channel-Mediated Mechanosensory Transduction. Cell Rep. 2019, 29, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, J.; Loukin, S.; Anishkin, A.; Kung, C. The Force-from-Lipid (FFL) Principle of Mechanosensitivity, at Large and in Elements. Pflügers Arch. Eur. J. Physiol. 2015, 467, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romantsov, T.; Guan, Z.; Wood, J.M. Cardiolipin and the Osmotic Stress Responses of Bacteria. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 2092–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Cox, C.D.; Bavi, N.; Rohde, P.R.; Nakayama, Y.; Martinac, B. Membrane Stiffness Is One of the Key Determinants of E. coli MscS Channel Mechanosensitivity. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Daday, C.; Gu, R.X.; Cox, C.D.; Martinac, B.; de Groot, B.L.; Walz, T. Visualization of the Mechanosensitive Ion Channel MscS under Membrane Tension. Nature 2021, 590, 509–514. [Google Scholar] [CrossRef]
- Anishkin, A.; Loukin, S.H.; Teng, J.; Kung, C. Feeling the Hidden Mechanical Forces in Lipid Bilayer Is an Original Sense. Proc. Natl. Acad. Sci. USA 2014, 111, 7898–7905. [Google Scholar] [CrossRef] [Green Version]
- Perozo, E.; Kloda, A.; Cortes, D.M.; Martinac, B. Physical Principles Underlying the Transduction of Bilayer Deformation Forces during Mechanosensitive Channel Gating. Nat. Struct. Biol. 2002, 9, 696–703. [Google Scholar] [CrossRef]
- Nomura, T.; Cranfield, C.G.; Deplazes, E.; Owen, D.M.; Macmillan, A.; Battle, A.R.; Constantine, M.; Sokabe, M.; Martinac, B. Differential Effects of Lipids and Lyso-Lipids on the Mechanosensitivity of the Mechanosensitive Channels MscL and MscS. Proc. Natl. Acad. Sci. USA 2012, 109, 8770–8775. [Google Scholar] [CrossRef] [Green Version]
- Carniello, V.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Role of Adhesion Forces in Mechanosensitive Channel Gating in Staphylococcus aureus Adhering to Surfaces. NPJ Biofilms Microbiomes 2020, 6, 31. [Google Scholar] [CrossRef]
- Li, J.; Busscher, H.J.; Swartjes, J.J.T.M.; Chen, Y.; Harapanahalli, A.K.; Norde, W.; van der Mei, H.C.; Sjollema, J. Residence-Time Dependent Cell Wall Deformation of Different Staphylococcus aureus Strains on Gold Measured Using Surface-Enhanced-Fluorescence. Soft Matter 2014, 10, 7638–7646. [Google Scholar] [CrossRef]
- Gu, J.; Valdevit, A.; Chou, T.M.; Libera, M. Substrate Effects on Cell-Envelope Deformation during Early-Stage: Staphylococcus aureus Biofilm Formation. Soft Matter 2017, 13, 2967–2976. [Google Scholar] [CrossRef] [PubMed]
- Culham, D.E.; Dalgado, C.; Gyles, C.L.; Mamelak, D.; MacLellan, S.; Wood, J.M. Osmoregulatory Transporter ProP Influences Colonization of the Urinary Tract by Escherichia coli. Microbiology 1998, 144, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Maurin, M.; Gyuranecz, M. Tularaemia: Clinical Aspects in Europe. Lancet Infect. Dis. 2016, 16, 113–124. [Google Scholar] [CrossRef]
- Sjöstedt, A. Tularemia: History, Epidemiology, Pathogen Physiology, and Clinical Manifestations. Ann. N. Y. Acad. Sci. 2007, 1105, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.; McGregor, A.; Cooley, L.; Ng, J.; Brown, M.; Ong, C.W.; Darcy, C.; Sintchenko, V. Francisella tularensis Subspecies Holarctica, Tasmania, Australia, 2011. Emerg. Infect. Dis. 2012, 18, 1484–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwaran, A.; Okoh, A.I. Human Campylobacteriosis: A Public Health Concern of Global Importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Lawe-Davies, O.; Bennett, S.; WHO. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Saudi Med. J. 2017, 38, 444–445. [Google Scholar]
- Ipe, D.S.; Horton, E.; Ulett, G.C. The Basics of Bacteriuria: Strategies of Microbes for Persistence in Urine. Front. Cell. Infect. Microbiol. 2016, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Schwan, W.R. Survival of Uropathogenic Escherichia coli in the Murine Urinary Tract Is Dependent on OmpR. Microbiology 2009, 155, 1832–1839. [Google Scholar] [CrossRef] [Green Version]
- Gargan, R.A.; Hamilton-Miller, J.M.; Brumfitt, W. Effect of PH and Osmolality on in Vitro Phagocytosis and Killing by Neutrophils in Urine. Infect. Immun. 1993, 61, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Sharma, S.; Chhibber, S.; Harjai, K. Effect of Osmolarity on Virulence of Uropathogenic Pseudomonas aeruginosa. Am. J. Biomed. Sci. 2009, 1, 12–26. [Google Scholar] [CrossRef]
- Wray, R.; Iscla, I.; Blount, P. Curcumin Activation of a Bacterial Mechanosensitive Channel Underlies Its Membrane Permeability and Adjuvant Properties. PLoS Pathog. 2021, 17, e1010198. [Google Scholar] [CrossRef] [PubMed]
- Wray, R.; Herrera, N.; Iscla, I.; Wang, J.; Blount, P. An Agonist of the MscL Channel Affects Multiple Bacterial Species and Increases Membrane Permeability and Potency of Common Antibiotics. Mol. Microbiol. 2019, 112, 896–905. [Google Scholar] [CrossRef]
- Wray, R.; Wang, J.; Iscla, I.; Blount, P. Novel MscL Agonists That Allow Multiple Antibiotics Cytoplasmic Access Activate the Channel through a Common Binding Site. PLoS ONE 2020, 15, e0228153. [Google Scholar] [CrossRef] [PubMed]
- Oswald, F.; Varadarajan, A.; Lill, H.; Peterman, E.J.G.; Bollen, Y.J.M. MreB-Dependent Organization of the E. coli Cytoplasmic Membrane Controls Membrane Protein Diffusion. Biophys. J. 2016, 110, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.; Börngen, K.; Nomura, T.; Battle, A.R.; Marin, K.; Martinac, B.; Krämer, R. Glutamate Efflux Mediated by Corynebacterium Glutamicum MscCG, Escherichia coli MscS, and Their Derivatives. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 1230–1240. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, G.; Xu, D.; Fan, L.; Wu, X.; Ni, X.; Zhao, S.; Zheng, P.; Sun, J.; Ma, Y. A Novel Corynebacterium Glutamicum l-Glutamate Exporter. Appl. Environ. Microbiol. 2018, 84, e02691-17. [Google Scholar] [CrossRef] [Green Version]
- Kaldalu, N.; Mei, R.; Lewis, K. Killing by Ampicillin and Ofloxacin Induces Overlapping Changes in Escherichia coli Transcription Profile. Antimicrob. Agents Chemother. 2004, 48, 890–896. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, A.; Fleurier, S.; Frénoy, A.; Dairou, J.; Bredeche, M.-F.; Sanchez-Vizuete, P.; Song, X.; Matic, I. Discovery and Function of a General Core Hormetic Stress Response in E. coli Induced by Sublethal Concentrations of Antibiotics. Cell Rep. 2016, 17, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Jiafeng, L.; Fu, X.; Chang, Z. Hypoionic Shock Treatment Enables Aminoglycosides Antibiotics to Eradicate Bacterial Persisters. Sci. Rep. 2015, 5, 14247. [Google Scholar] [CrossRef]
- Zhao, Y.; Lv, B.; Sun, F.; Liu, J.; Wang, Y.; Gao, Y.; Qi, F.; Chang, Z.; Fu, X. Rapid Freezing Enables Aminoglycosides to Eradicate Bacterial Persisters via Enhancing Mechanosensitive Channel MscL-Mediated Antibiotic Uptake. mBio 2020, 11, e03239-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, J.; Hirano, S.; Ito, H.; Wachi, M. Mutations of the Corynebacterium Glutamicum NCgl1221 Gene, Encoding a Mechanosensitive Channel Homolog, Induce l-Glutamic Acid Production. Appl. Environ. Microbiol. 2007, 73, 4491–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, K.; Murata, J.; Konishi, T.; Yabe, I.; Nakamatsu, T.; Kawasaki, H. Glutamate Is Excreted Across the Cytoplasmic Membrane through the NCgl1221 Channel of Corynebacterium Glutamicum by Passive Diffusion. Biosci. Biotechnol. Biochem. 2012, 76, 1422–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, T.; Saito, M.; Yoshikawa, K.; Furusawa, C.; Shmizu, H. Integrated Analysis of the Transcriptome and Metabolome of Corynebacterium Glutamicum during Penicillin-Induced Glutamic Acid Production. Biotechnol. J. 2018, 13, 1700612. [Google Scholar] [CrossRef] [PubMed]
- Koprowski, P.; Grajkowski, W.; Balcerzak, M.; Filipiuk, I.; Fabczak, H.; Kubalski, A. Cytoplasmic Domain of MscS Interacts with Cell Division Protein FtsZ: A Possible Non-Channel Function of the Mechanosensitive Channel in Escherichia coli. PLoS ONE 2015, 10, e0127029. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Wright, S.N.; Hamblin, M.; McCloskey, D.; Alcantar, M.A.; Schrübbers, L.; Lopatkin, A.J.; Satish, S.; Nili, A.; Palsson, B.O.; et al. A White-Box Machine Learning Approach for Revealing Antibiotic Mechanisms of Action. Cell 2019, 177, 1649–1661.e9. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Harada, M.; Funakoshi, S.; Fujii, N.; Miyajima, K. Physicochemical Determinants for the Interactions of Magainins 1 and 2 with Acidic Lipid Bilayers. Biochim. Biophys. Acta-Biomembr. 1991, 1063, 162–170. [Google Scholar] [CrossRef]
- Dürr, U.H.N.; Sudheendra, U.S.; Ramamoorthy, A. LL-37, the Only Human Member of the Cathelicidin Family of Antimicrobial Peptides. Biochim. Biophys. Acta-Biomembr. 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, S.K.; Larson, R.G. Effect of Salt on the Interactions of Antimicrobial Peptides with Zwitterionic Lipid Bilayers. Biochim. Biophys. Acta-Biomembr. 2006, 1758, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Ding, J.; Li, H.; Li, L.; Zhao, R.; Shen, Z.; Fan, X.; Xi, T. Effects of Cations and PH on Antimicrobial Activity of Thanatin and S-Thanatin against Escherichia coli ATCC25922 and B. subtilis ATCC 21332. Curr. Microbiol. 2008, 57, 552–557. [Google Scholar] [CrossRef] [Green Version]
- Aoki, W.; Ueda, M. Characterization of Antimicrobial Peptides toward the Development of Novel Antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef] [Green Version]
- Wahid, M.H.; Stroeher, U.H.; Eroglu, E.; Chen, X.; Vimalanathan, K.; Raston, C.L.; Boulos, R.A. Aqueous Based Synthesis of Antimicrobial-Decorated Graphene. J. Colloid Interface Sci. 2015, 443, 88–96. [Google Scholar] [CrossRef]
- Wright, L.; Rao, S.; Thomas, N.; Boulos, R.A.; Prestidge, C.A. Ramizol ® Encapsulation into Extended Release PLGA Micro- and Nanoparticle Systems for Subcutaneous and Intramuscular Administration: In Vitro and in Vivo Evaluation. Drug Dev. Ind. Pharm. 2018, 44, 1451–1457. [Google Scholar] [CrossRef]
- Sibley, K.; Chen, J.; Koetzner, L.; Mendes, O.; Kimzey, A.; Lansita, J.; Boulos, R.A. A 14-Day Repeat Dose Oral Gavage Range-Finding Study of a First-in-Class CDI Investigational Antibiotic, in Rats. Sci. Rep. 2019, 9, 158. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, C.; Pagano, P.; Pillar, C.M.; Shinabarger, D.L.; Boulos, R.A. Comparison of the in Vitro Antibacterial Activity of Ramizol, Fidaxomicin, Vancomycin, and Metronidazole against 100 Clinical Isolates of Clostridium Difficile by Broth Microdilution. Diagn. Microbiol. Infect. Dis. 2018, 92, 250–252. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Prestidge, C.A.; Miesel, L.; Sweeney, D.; Shinabarger, D.L.; Boulos, R.A. Preclinical Development of Ramizol, an Antibiotic Belonging to a New Class, for the Treatment of Clostridium difficile Colitis. J. Antibiot. 2016, 69, 879–884. [Google Scholar] [CrossRef] [Green Version]
- Iscla, I.; Wray, R.; Blount, P.; Larkins-Ford, J.; Conery, A.L.; Ausubel, F.M.; Ramu, S.; Kavanagh, A.; Huang, J.X.; Blaskovich, M.A.; et al. A New Antibiotic with Potent Activity Targets MscL. J. Antibiot. 2015, 68, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Wray, R.; Iscla, I.; Kovacs, Z.; Wang, J.; Blount, P. Novel Compounds That Specifically Bind and Modulate MscL: Insights into Channel Gating Mechanisms. FASEB J. 2019, 33, 3180–3189. [Google Scholar] [CrossRef] [Green Version]
- Wray, R.; Blount, P.; Wang, J.; Iscla, I. In Silico Screen Identifies a New Family of Agonists for the Bacterial Mechanosensitive Channel MscL. Antibiotics 2022, 11, 433. [Google Scholar] [CrossRef]
- Nguyen, T.; Clare, B.; Guo, W.; Martinac, B. The Effects of Parabens on the Mechanosensitive Channels of E. coli. Eur. Biophys. J. 2005, 34, 389–395. [Google Scholar] [CrossRef]
- Kamaraju, K.; Sukharev, S. The Membrane Lateral Pressure-Perturbing Capacity of Parabens and Their Effects on the Mechanosensitive Channel Directly Correlate with Hydrophobicity. Biochemistry 2008, 47, 10540–10550. [Google Scholar] [CrossRef] [PubMed]
- Comert, F.; Greenwood, A.; Maramba, J.; Acevedo, R.; Lucas, L.; Kulasinghe, T.; Cairns, L.S.; Wen, Y.; Fu, R.; Hammer, J.; et al. The Host-Defense Peptide Piscidin P1 Reorganizes Lipid Domains in Membranes and Decreases Activation Energies in Mechanosensitive Ion Channels. J. Biol. Chem. 2019, 294, 18557–18570. [Google Scholar] [CrossRef] [PubMed]
- Cetuk, H.; Maramba, J.; Britt, M.; Scott, A.J.; Ernst, R.K.; Mihailescu, M.; Cotten, M.L.; Sukharev, S. Differential Interactions of Piscidins with Phospholipids and Lipopolysaccharides at Membrane Interfaces. Langmuir 2020, 36, 5065–5077. [Google Scholar] [CrossRef] [PubMed]
- Hurst, A.C.; Gottlieb, P.A.; Martinac, B. Concentration Dependent Effect of GsMTx4 on Mechanosensitive Channels of Small Conductance in E. coli Spheroplasts. Eur. Biophys. J. 2009, 38, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Kamaraju, K.; Gottlieb, P.A.; Sachs, F.; Sukharev, S. Effects of GsMTx4 on Bacterial Mechanosensitive Channels in Inside-Out Patches from Giant Spheroplasts. Biophys. J. 2010, 99, 2870–2878. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Kim, P.I.; Lee, S.K.; Lee, C.W.; Eu, Y.J.; Lee, D.G.; Earm, Y.E.; Kim, J.I. Lipid Membrane Interaction and Antimicrobial Activity of GsMTx-4, an Inhibitor of Mechanosensitive Channel. Biochem. Biophys. Res. Commun. 2006, 340, 633–638. [Google Scholar] [CrossRef]
- Berrier, C.; Coulombe, A.; Szabo, I.; Zoratti, M.; Ghazi, A. Gadolinium Ion Inhibits Loss of Metabolites Induced by Osmotic Shock and Large Stretch-activated Channels in Bacteria. Eur. J. Biochem. 1992, 206, 559–565. [Google Scholar] [CrossRef]
- Ermakov, Y.A.; Kamaraju, K.; Sengupta, K.; Sukharev, S. Gadolinium Ions Block Mechanosensitive Channels by Altering the Packing and Lateral Pressure of Anionic Lipids. Biophys. J. 2010, 98, 1018–1027. [Google Scholar] [CrossRef] [Green Version]
- Martinac, B.; Adler, J.; Kung, C. Mechanosensitive Ion Channels of E. coli Activated by Amphipaths. Nature 1990, 348, 261–263. [Google Scholar] [CrossRef]
- Atcha, H.; Jairaman, A.; Holt, J.R.; Meli, V.S.; Nagalla, R.R.; Veerasubramanian, P.K.; Brumm, K.T.; Lim, H.E.; Othy, S.; Cahalan, M.D.; et al. Mechanically Activated Ion Channel Piezo1 Modulates Macrophage Polarization and Stiffness Sensing. Nat. Commun. 2021, 12, 3256. [Google Scholar] [CrossRef]
- Beaulieu-Laroche, L.; Christin, M.; Donoghue, A.; Agosti, F.; Yousefpour, N.; Petitjean, H.; Davidova, A.; Stanton, C.; Khan, U.; Dietz, C.; et al. TACAN Is an Ion Channel Involved in Sensing Mechanical Pain. Cell 2020, 180, 956–967.e17. [Google Scholar] [CrossRef]
- Tran, D.; Petitjean, H.; Chebli, Y.; Geitmann, A.; Sharif-Naeini, R. Mechanosensitive Ion Channels Contribute to Mechanically Evoked Rapid Leaflet Movement in Mimosa pudica. Plant Physiol. 2021, 187, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Parisi, C.; Chandaria, V.V.; Nowlan, N.C. Blocking Mechanosensitive Ion Channels Eliminates the Effects of Applied Mechanical Loading on Chick Joint Morphogenesis. Philos. Trans. R. Soc. B 2018, 373, 20170317. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Gu, C. Physiological and Pathological Functions of Mechanosensitive Ion Channels. Mol. Neurobiol. 2014, 50, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Martinac, B.; Nomura, T.; Chi, G.; Petrov, E.; Rohde, P.R.; Battle, A.R.; Foo, A.; Constantine, M.; Rothnagel, R.; Carne, S.; et al. Bacterial Mechanosensitive Channels: Models for Studying Mechanosensory Transduction. Antioxid. Redox Signal. 2014, 20, 952–969. [Google Scholar] [CrossRef] [Green Version]
- Herrera, N.; Maksaev, G.; Haswell, E.S.; Rees, D.C. Elucidating a Role for the Cytoplasmic Domain in the Mycobacterium Tuberculosis Mechanosensitive Channel of Large Conductance. Sci. Rep. 2018, 8, 14566. [Google Scholar] [CrossRef]
- Wang, W.; Black, S.S.; Edwards, M.D.; Miller, S.; Morrison, E.L.; Bartlett, W.; Dong, C.; Naismith, J.H.; Booth, I.R. The Structure of an Open Form of an E. coli Mechanosensitive Channel at 3.45 å Resolution. Science 2008, 321, 1179–1183. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, T.; Flegler, V.J.; Rasmussen, A.; Böttcher, B. Structure of the Mechanosensitive Channel MscS Embedded in the Membrane Bilayer. J. Mol. Biol. 2019, 431, 3081–3090. [Google Scholar] [CrossRef]
- Pliotas, C.; Ward, R.; Branigan, E.; Rasmussen, A.; Hagelueken, G.; Huang, H.; Black, S.S.; Booth, I.R.; Schiemann, O.; Naismith, J.H. Conformational State of the MscS Mechanosensitive Channel in Solution Revealed by Pulsed Electron-Electron Double Resonance (PELDOR) Spectroscopy. Proc. Natl. Acad. Sci. USA 2012, 109, E2675–E2682. [Google Scholar] [CrossRef] [Green Version]
- Dorwart, M.R.; Wray, R.; Brautigam, C.A.; Jiang, Y.; Blount, P.S. Aureus MscL Is a Pentamer In Vivo but of Variable Stoichiometries In Vitro: Implications for Detergent-Solubilized Membrane Proteins. PLoS Biol. 2010, 8, e1000555. [Google Scholar] [CrossRef]
- Liu, Z.; Gandhi, C.S.; Rees, D.C. Structure of a Tetrameric MscL in an Expanded Intermediate State. Nature 2009, 461, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pliotas, C.; Dahl, A.C.E.; Rasmussen, T.; Mahendran, K.R.; Smith, T.K.; Marius, P.; Gault, J.; Banda, T.; Rasmussen, A.; Miller, S.; et al. The Role of Lipids in Mechanosensation. Nat. Struct. Mol. Biol. 2015, 22, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angiulli, G.; Dhupar, H.S.; Suzuki, H.; Wason, I.S.; Duong Van Hoa, F.; Walz, T. New Approach for Membrane Protein Reconstitution into Peptidiscs and Basis for Their Adaptability to Different Proteins. eLife 2020, 9, e53530. [Google Scholar] [CrossRef] [PubMed]
- Flegler, V.J.; Rasmussen, A.; Borbil, K.; Boten, L.; Chen, H.A.; Deinlein, H.; Halang, J.; Hellmanzik, K.; Löffler, J.; Schmidt, V.; et al. Mechanosensitive Channel Gating by Delipidation. Proc. Natl. Acad. Sci. USA 2021, 118, e2107095118. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, B.; Zhang, Y.; Xu, C.-Q.; Zhuo, W.; Ge, J.; Li, J.; Gao, N.; Li, Y.; Yang, M. A Binding-Block Ion Selective Mechanism Revealed by a Na/K Selective Channel. Protein Cell 2018, 9, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, J.; Feng, Y.; Ge, J.; Li, W.; Sun, W.; Iscla, I.; Yu, J.; Blount, P.; Li, Y.; et al. Structure and Molecular Mechanism of an Anion-Selective Mechanosensitive Channel of Small Conductance. Proc. Natl. Acad. Sci. USA 2012, 109, 18180–18185. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.Y.; Poon, Y.S.; Kaiser, J.T.; Rees, D.C. Open and Shut: Crystal Structures of the Dodecylmaltoside Solubilized Mechanosensitive Channel of Small Conductance from Escherichia coli and Helicobacter pylori at 4.4 Å and 4.1 Å Resolutions. Protein Sci. 2013, 22, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Flegler, V.J.; Rasmussen, A.; Rao, S.; Wu, N.; Zenobi, R.; Sansom, M.S.P.; Hedrich, R.; Rasmussen, T.; Böttcher, B. The MscS-like Channel YnaI Has a Gating Mechanism Based on Flexible Pore Helices. Proc. Natl. Acad. Sci. USA 2020, 117, 28754–28762. [Google Scholar] [CrossRef]
- Catalano, C.; Ben-Hail, D.; Qiu, W.; Blount, P.; des Georges, A.; Guo, Y. Cryo-EM Structure of Mechanosensitive Channel YnaI Using SMA2000: Challenges and Opportunities. Membranes 2021, 11, 849. [Google Scholar] [CrossRef]
- Hu, W.; Wang, Z.; Zheng, H. Extended Sensor Paddles with Bound Lipids Revealed in Mechanosensitive Channel YnaI. Available online: https://doi.org/10.2210/pdb6URT/pdb (accessed on 11 May 2022).
- Walton, T.A.; Rees, D.C. Structure and Stability of the C-Terminal Helical Bundle of the E. coli Mechanosensitive Channel of Large Conductance. Protein Sci. 2013, 22, 1592–1601. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ke, M. Mechanosensitive Channel MscS K180R Mutant. Available online: https://www.rcsb.org/structure/7DLU (accessed on 11 May 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds Entering Cells through Mechanosensitive Channels | Compounds Triggering Amino-Acid Release from Mechanosensitive Channels |
|---|---|
| dihydrostreptomycin [19,20] | MP196 [11] |
| spectinomycin [19] | gramicidin A [11] |
| viomycin [19] | gramicidin S [11] |
| nifuroxazide [19] | nisin [11] |
| curcumin [63] | aurein 2.2 [11,21] |
| tetracycline [64,65] | aurein 2.3 [21] |
| * sublancin 168 [18] | aurein 2.2Δ3 [21] |
| * penicillin [28,66,67,68,69,70] | |
| * ampicillin [28,66,67,68,69,70] |
| Compound | Target | Mechanism | Structural Class | Activity Shown against |
|---|---|---|---|---|
| compounds directly targeting mechanosensitive channels | ||||
| ramizol [83,84,85,86,87,88] | MscL MscS | reduces gating threshold | styrylbenzene | S. aureus, Streptococcus pneumoniae, Clostridium difficile |
| 011A [64,65,89] | MscL | stabilizes open state, increases permeability for antibiotics | small organic molecule | E. coli, S. aureus, Mycolicibacterium smegmatis |
| K05 [64,65,89] | MscL | stabilizes open state, increases permeability for antibiotics | small organic molecule | E. coli, S. aureus, M. smegmatis |
| compound 262 [90] | MscL | stabilizes open state, increases permeability for antibiotics | small organic molecule | E. coli, M. tuberculosis |
| compounds indirectly targeting mechanosensitive channels | ||||
| parabens [91,92] | MscL MscS | modulates gating | 4-hydroxylbenzoic acid ester | E. coli |
| piscidins (P1 and P3) [93,94] | MscL MscS | sensitizes channel gating | alpha-helical peptide | E. coli |
| GsMTx4 [95,96,97] | MscL MscS | promotes or inhibits channel gating; concentration-dependent | globular peptide | E. coli |
| gadolinium chloride (GdCl3) [98,99] | MscL MscS | inhibits channel gating | inorganic salt | E. coli, B. subtilis, Enterococcus faecalis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidarta, M.; Baruah, L.; Wenzel, M. Roles of Bacterial Mechanosensitive Channels in Infection and Antibiotic Susceptibility. Pharmaceuticals 2022, 15, 770. https://doi.org/10.3390/ph15070770
Sidarta M, Baruah L, Wenzel M. Roles of Bacterial Mechanosensitive Channels in Infection and Antibiotic Susceptibility. Pharmaceuticals. 2022; 15(7):770. https://doi.org/10.3390/ph15070770
Chicago/Turabian StyleSidarta, Margareth, Luna Baruah, and Michaela Wenzel. 2022. "Roles of Bacterial Mechanosensitive Channels in Infection and Antibiotic Susceptibility" Pharmaceuticals 15, no. 7: 770. https://doi.org/10.3390/ph15070770
APA StyleSidarta, M., Baruah, L., & Wenzel, M. (2022). Roles of Bacterial Mechanosensitive Channels in Infection and Antibiotic Susceptibility. Pharmaceuticals, 15(7), 770. https://doi.org/10.3390/ph15070770