
Wheat Water-Soluble Carbohydrate Remobilisation under Water Deficit by 1-FEH w3


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Backcrossing for 90K SNP Analysis
2.3. Genotypic DNA Extraction
2.4. The 90K SNP Chip Analysis
2.5. The 90K SNP Data Analysis
2.6. Glasshouse Experimental Setup
2.7. Imposing Water Deficit
2.8. Sampling
2.9. Carbohydrate Analysis
2.10. Calculation of WSC Remobilisation
2.11. Data Analysis
3. Results
3.1. Detecting the Mutation Region
3.2. Mapping of Mutation Regions in Chromosomes
3.3. Gene Annotation at the Mutated Region and Functional Analysis
3.4. Observation of the Water-Deficit Regime in Mutated Lines
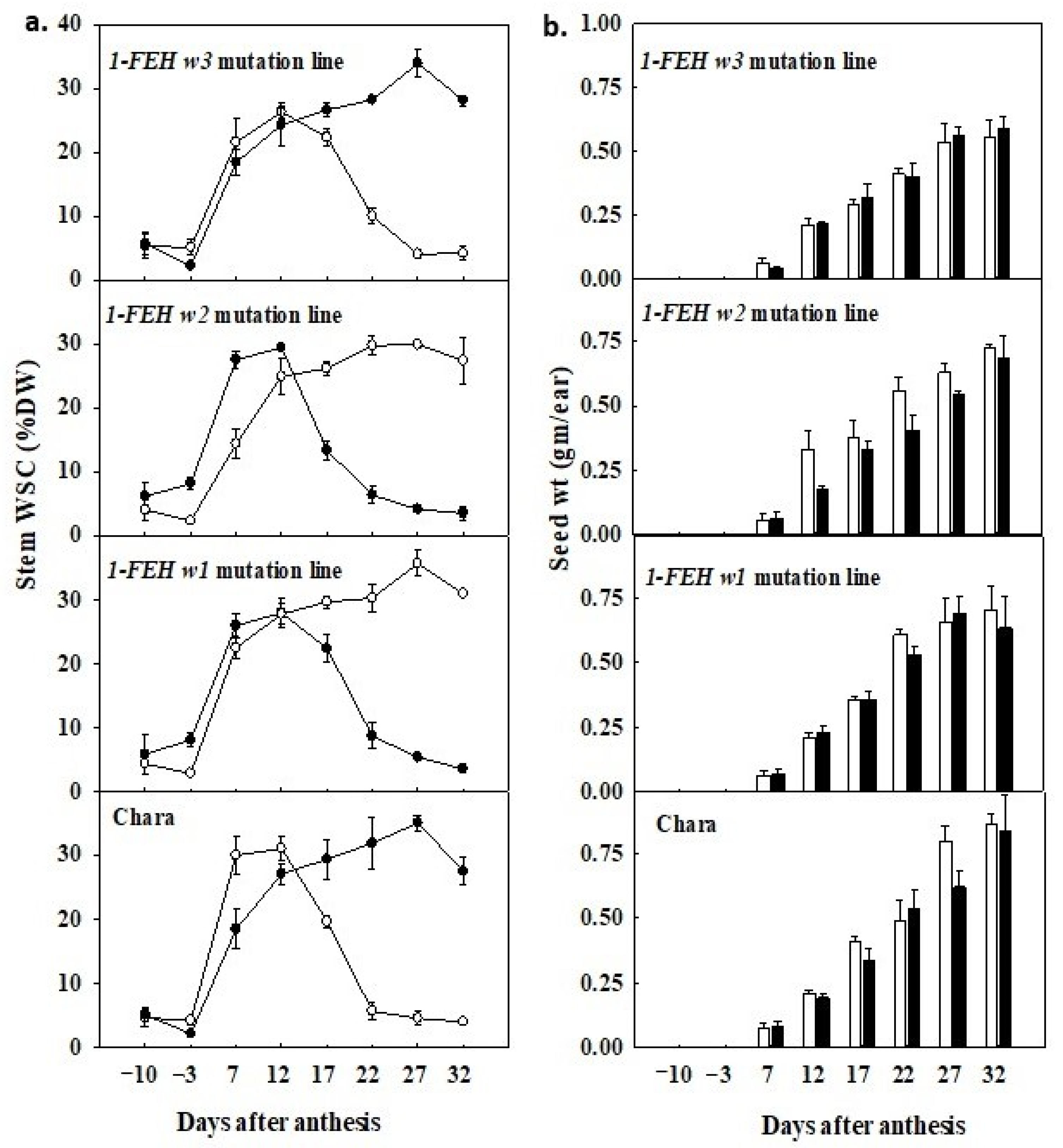
3.5. Water Deficit Promoted Early Remobilisation of WSC
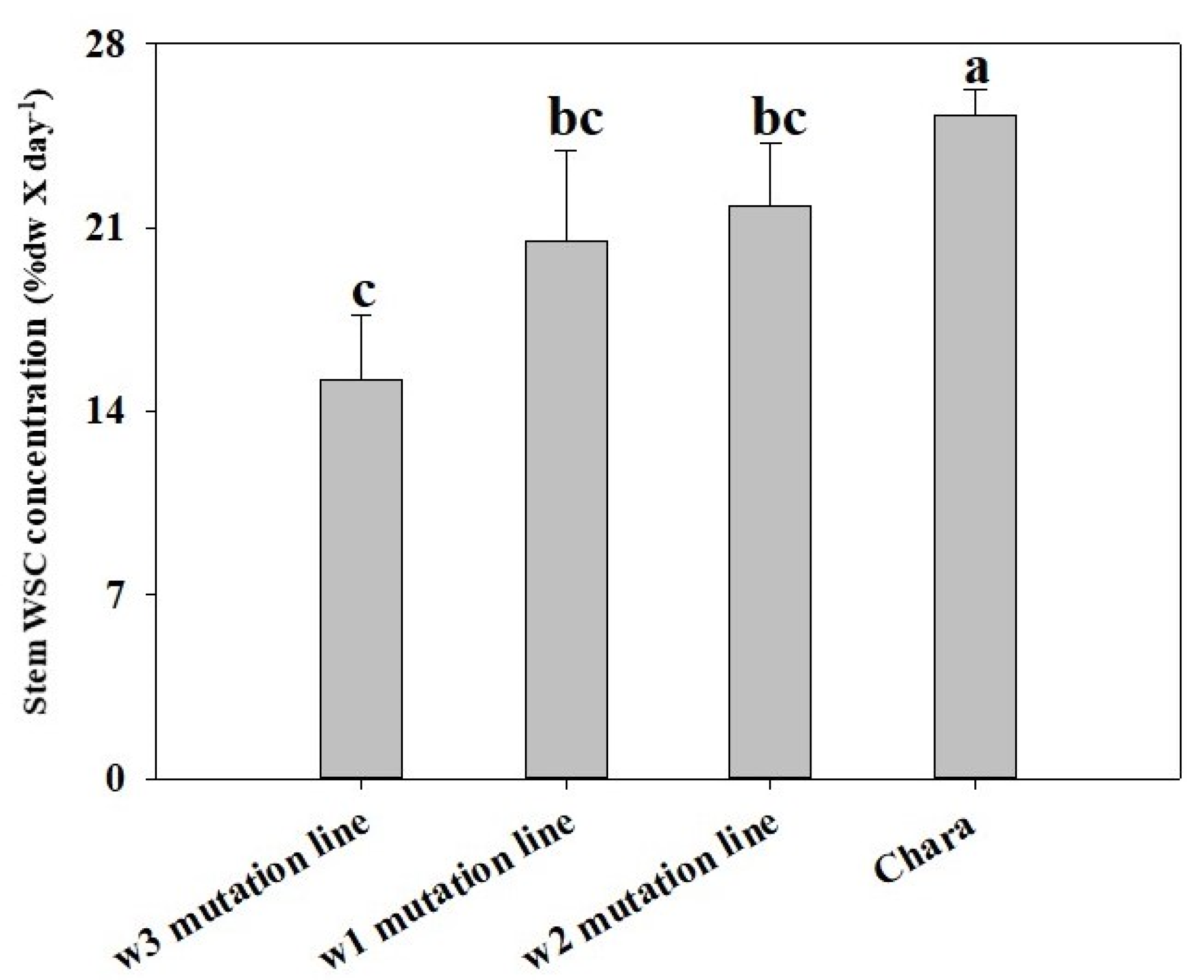
3.6. Timing of Decline in WSC Differed among Mutation Lines
3.7. WSC Concentration and Grain Development
3.8. Grain Weight at Maturity Was Influenced by 1-FEH Isoforms under Water-Deficit Condition
3.9. Correlation Analysis of Grain Weight with WSC Level and Remobilisation Rate
4. Discussion
4.1. Characterisation of 1-FEH Gene Mutations
4.2. Chromosomal Mapping of Mutation Regions and Candidate Genes
4.3. Annotation of the Deleted Region
4.4. Function Prediction for Non-Target Genes in Deleted Regions
4.5. Influence of 1-FEH Gene Mutations on WSC Remobilisation
4.6. The 1-FEH Gene Mutations Showed Functional Variation Only under Water-Deficit Condition Not under Well-Watered Condition
4.7. Under Water-Deficit Conditions, WSC Remobilisation Rate and Grain Filling Were Slowed Down by the Deletion of 1-FEH w3
4.8. Components of Grain Yield Were Reduced in the 1-FEH w3 Mutation Line but Not in 1-FEH w2 or 1-FEH w1 Mutation Lines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yáñez, A.; Tapia, G.; Guerra, F.; Del Pozo, A. Stem carbohydrate dynamics and expression of genes involved in fructan accumulation and remobili-zation during grain growth in wheat (Triticum aestivum L.) genotypes with contrasting tolerance to water stress. PLoS ONE 2017, 12, e0177667. [Google Scholar] [CrossRef] [Green Version]
- Ende, W.V.D.; Clerens, S.; Vergauwen, R.; Van Riet, L.; Van Laere, A.; Yoshida, M.; Kawakami, A. Fructan 1-Exohydrolases. β-(2,1)-Trimmers during Graminan Biosynthesis in Stems of Wheat? Purification, Characterization, Mass Mapping, and Cloning of Two Fructan 1-Exohydrolase Isoforms. Plant Physiol. 2003, 131, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ende, W.V.D.; Michiels, A.; Van Wonterghem, D.; Clerens, S.P.; De Roover, J.; Van Laere, A.J. Defoliation Induces Fructan 1-Exohydrolase II in Witloof Chicory Roots. Cloning and Purification of Two Isoforms, Fructan 1-Exohydrolase IIa and Fructan 1-Exohydrolase IIb. Mass Fingerprint of the Fructan 1-Exohydrolase II Enzymes. Plant Physiol. 2001, 126, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Doyle, A.; Fischer, R. Dry matter accumulation and water use relationships in wheat crops. Aust. J. Agric. Res. 1979, 30, 815–829. [Google Scholar] [CrossRef]
- Van Riet, L.; Nagaraj, V.; Ende, W.V.D.; Clerens, S.; Wiemken, A.; Van Laere, A. Purification, cloning and functional characterization of a fructan 6-exohydrolase from wheat (Triticum aestivum L.). J. Exp. Bot. 2006, 57, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Yoshida, M. Fructan: Fructan 1-fructosyltransferase, a key enzyme for biosynthesis of graminan oli-gomers in hardened wheat. Planta 2005, 223, 90–104. [Google Scholar] [CrossRef]
- Valluru, R.; Van den Ende, W. Plant fructans in stress environments: Emerging concepts and future prospects. J. Exp. Bot. 2008, 59, 2905–2916. [Google Scholar] [CrossRef] [Green Version]
- Van Riet, L.; Altenbach, D.; Vergauwen, R.; Clerens, S.; Kawakami, A.; Yoshida, M.; Ende, W.V.D.; Wiemken, A.; Van Laere, A. Purification, cloning and functional differences of a third fructan 1-exohydrolase (1-FEHw3) from wheat (Triticum aestivum). Physiol. Plant. 2008, 133, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Liu, L. Activities of fructan- and sucrose-metabolizing enzymes in wheat stems subjected to water stress during grain filling. Planta 2004, 220, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dell, B.; Conocono, E.; Waters, I.; Setter, T.; Appels, R. Water deficits in wheat: Fructan exohydrolase (1-FEH) mRNA expression and relationship to soluble carbohydrate concentrations in two varieties. New Phytol. 2009, 181, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, T.L.; Kazan, K.; Li, Z.; Morell, M.K.; Manners, J.M. A high-throughput method for the detection of homoeologous gene deletions in hexaploid wheat. BMC Plant Biol. 2010, 10, 264. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.; Meinke, D. A Comprehensive Dataset of Genes with a Loss-of-Function Mutant Phenotype in Arabidopsis. Plant Physiol. 2012, 158, 1115–1129. [Google Scholar] [CrossRef] [Green Version]
- Knoll, J.E.; Ramos, M.L.; Zeng, Y.; Holbrook, C.C.; Chow, M.; Chen, S.; Maleki, S.; Bhattacharya, A.; Ozias-Akins, P. TILLING for allergen reduction and improvement of quality traits in peanut (Arachis hypogaea L.). BMC Plant Biol. 2011, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- The International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Stein, N.; Choulet, F.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, T.L.; Kazan, K.; Manners, J.M. The Application of Reverse Genetics to Polyploid Plant Species. Crit. Rev. Plant Sci. 2012, 31, 181–200. [Google Scholar] [CrossRef]
- Šafář, J.; Šimková, H.; Kubaláková, M.; Číhalíková, J.; Suchánková, P.; Bartoš, J.; Doležel, J. Development of Chromosome-Specific BAC Resources for Genomics of Bread Wheat. Cytogenet. Genome Res. 2010, 129, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Wanjugi, H.; Coleman-Derr, D.; Huo, N.; Kianian, S.F.; Luo, M.-C.; Wu, J.; Anderson, O.; Gu, Y.Q. Rapid development of PCR-based genome-specific repetitive DNA junction markers in wheat. Genome 2009, 52, 576–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.; Duran, C.; Berkman, P.J.; Lorenc, M.T.; Stiller, J.; Manoli, S.; Hayden, M.J.; Forrest, K.L.; Fleury, D.; Baumann, U.; et al. Single nucleotide polymorphism discovery from wheat next-generation sequence data. Plant Biotechnol. J. 2012, 10, 73–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.L.; Uauy, C.; Robson, F.; Till, B. TILLING in extremis. Plant Biotechnol. J. 2012, 10, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, E.; Mulligan, B.J.; Covey, S.N.; Milner, J.J. Characterization of gamma irradiation-induced deletion mutations at a selectable locus in Arabidopsis. Mutat. Res. Mol. Mech. Mutagen. 1998, 401, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.; Kusaba, M.; Iida, S.; Yamaguchi, H.; Nishio, T.; Nishimura, M. Molecular characterization of mutations induced by gamma irradiation in rice. Genes Genet. Syst. 2009, 84, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordström, K.J.V.; Albani, M.C.; James, G.V.; Gutjahr, C.; Hartwig, B.; Turck, F.; Paszkowski, U.; Coupland, G.; Schneeberger, K. Mutation identification by direct comparison of whole-genome sequencing data from mutant and wild-type individuals using k-mers. Nat. Biotechnol. 2013, 31, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Akond, M.; Liu, S.; Schoener, L.; Anderson, J.A.; Kantartzi, S.K.; Meksem, K.; Song, O.; Wang, D.; Wen, Z.; Lightfoot, D.A.; et al. A SNP-based genetic linkage map of soybean using the SoySNP6K Illumina Infinium BeadChip geno-typing array. Plant Genet. Genom. Biotechnol. 2013, 1, 80–89. [Google Scholar] [CrossRef]
- Cook, J.P.; McMullen, M.D.; Holland, J.B.; Tian, F.; Bradbury, P.; Ross-Ibarra, J.; Buckler, E.S.; Flint-Garcia, S.A. Genetic Architecture of Maize Kernel Composition in the Nested Association Mapping and Inbred Association Panels. Plant Physiol. 2012, 158, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganal, M.W.; Durstewitz, G.; Polley, A.; Bérard, A.; Buckler, E.S.; Charcosset, A.; Clarke, J.D.; Graner, E.-M.; Hansen, M.; Joets, J.; et al. A Large Maize (Zea mays L.) SNP Genotyping Array: Development and Germplasm Genotyping, and Genetic Mapping to Compare with the B73 Reference Genome. PLoS ONE 2011, 6, e28334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, S.-C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; Van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a Large SNP Genotyping Array and Generation of High-Density Genetic Maps in Tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef] [PubMed]
- Wiedmann, R.T.; Smith, T.P.; Nonneman, D.J. SNP discovery in swine by reduced representation and high throughput pyrosequencing. BMC Genet. 2008, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Tung, C.-W.; Eizenga, G.C.; Wright, M.H.; Ali, M.L.; Price, A.H.; Norton, G.J.; Islam, M.R.; Reynolds, A.; Mezey, J.; et al. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun. 2011, 2, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, M.; Wenzl, P.; Caig, V.; Carling, J.; Xia, L.; Yang, S.; Uszynski, G.; Mohler, V.; Lehmensiek, A.; Kuchel, H.; et al. Diversity arrays technology (DArT) for high-throughput profiling of the hexaploid wheat genome. Theor. Appl. Genet. 2006, 113, 1409–1420. [Google Scholar] [CrossRef]
- Trebbi, D.; Maccaferri, M.; de Heer, P.; Sørensen, A.; Giuliani, S.; Salvi, S.; Sanguineti, M.C.; Massi, A.; van der Vossen, E.A.G.; Tuberosa, R. High-throughput SNP discovery and genotyping in durum wheat (Triticum durum Desf.). Theor. Appl. Genet. 2011, 123, 555–569. [Google Scholar] [CrossRef]
- Allen, A.M.; Barker, G.L.; Berry, S.T.; Coghill, J.A.; Gwilliam, R.; Kirby, S.; Robinson, P.; Brenchley, R.C.; D’Amore, R.; McKenzie, N.; et al. Transcript-specific, single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L.). Plant Biotechnol. J. 2011, 9, 1086–1099. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef]
- Keeble-Gagnère, G.; Pasam, R.; Forrest, K.L.; Wong, D.; Robinson, H.; Godoy, J.; Rattey, A.; Moody, D.; Mullan, D.; Walmsley, T.; et al. Novel Design of Imputation-Enabled SNP Arrays for Breeding and Research Applications Supporting Multi-Species Hybridization. Front. Plant Sci. 2021, 12, 756877. [Google Scholar] [CrossRef]
- Akhunov, E.; Nicolet, C.; Dvorak, J. Single nucleotide polymorphism genotyping in polyploid wheat with the Illumina GoldenGate assay. Theor. Appl. Genet. 2009, 119, 507–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.; Wang, P.; Vincent, D.; Khan, J.M.; Juhasz, A.; Diepeveen, D.; Lipscombe, R.J.; Ma, W.; Appels, R. Proteomic profiling of developing wheat heads under water-stress. Funct. Integr. Genom. 2020, 20, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Fales, F. The assimilation and degradation of carbohydrates by yeast cells. J. Biol. Chem. 1951, 193, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Zuryn, S.; Le Gras, S.; Jamet, K.; Jarriault, S. A Strategy for Direct Mapping and Identification of Mutations by Whole-Genome Sequencing. Genetics 2010, 186, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of Carbon Reserves Is Improved by Controlled Soil-Drying during Grain Filling of Wheat. Crop. Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Cruz-Aguado, J.A.; Rodés, R.; Pérez, I.P.; Dorado, M. Morphological characteristics and yield components associated with accumulation and loss of dry mass in the internodes of wheat. Field Crop. Res. 2000, 66, 129–139. [Google Scholar] [CrossRef]
- Simmons, S.; Oelke, E.; Anderson, P. Growth and Development Guide for Spring Wheat; University of Minnesota Agricultural Extension Service: St. Paul, MN, USA, 1985. [Google Scholar]
- Zhang, J.; Dell, B.; Biddulph, B.; Khan, N.; Xu, Y.; Luo, H.; Appels, R. Vernalization gene combination to maximize grain yield in bread wheat (Triticum aestivum L.) in diverse environments. Euphytica 2014, 198, 439–454. [Google Scholar] [CrossRef]
- Khoshro, H.H.; Taleei, A.; Bihamta, M.R.; Shahbazi, M.; Abbasi, A.; Ramezanpour, S.S. Expression analysis of the genes involved in accumulation and remobilization of assimilates in wheat stem under terminal drought stress. Plant Growth Regul. 2014, 74, 165–176. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Chen, W.; Dell, B.; Vergauwen, R.; Biddulph, B.; Khan, N.; Luo, H.; Appels, R.; Van den Ende, W.; et al. A wheat 1-FEH w3 variant underlies enzyme activity for stem WSC remobilization to grain under drought. New Phytol. 2015, 205, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | w1 mutant line -BW22-M3-22-w1 |
| w3 mutant line -BW18b-M3-426-w3 | |
| w2 mutant line -BW21-M3-433-w2 | |
| Parent: Chara | |
| Treatments | Terminal water deficit (water deficit imposed from anthesis (50% plants at the flowering stage) until maturity) |
| Wellwatered | |
| Sample collection growth stages | AR5 (Auricle distance 5 cm) |
| Fully emerged spike | |
| 7 days after anthesis | |
| 12 days after anthesis | |
| 17 days after anthesis | |
| 22 days after anthesis | |
| 27 days after anthesis | |
| 32 days after anthesis | |
| At grain maturity |
| 6A | 6A x C | 6B | 6B x C | 6D | 6D x C | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chr | Size of the Deletion (bp) | Region Containing FEH-like Sequence | 22 | C X22 | 426 | Cx 426 | 433 | Cx 433 | SNP Index | SNP Name | Position | Description of Observed Poly | Comment |
| 6AS | 28,246,961 | D | H | 76,092 | wsnp_CAP8_c6680_3136899 | 1,28,36,428---1,28,36,621 | Poly | SNP near top of del in 22 | |||||
| D | H | 3722 | BobWhite_c5092_422 | 1,30,71,495-----1,30,71,594 | Null | Null SNP near top of del in 22 | |||||||
| FEH_6A | D | H | |||||||||||
| D | H | 71,378 | Tdurum_contig42906_732 | 3,26,58,889----3,26,58,989 | Null | Example of SNP located in FEH | |||||||
| D | H | d | h | d | h | 55,811 | RAC875_c25556_1250 | 3,26,60,703---3,26,60,803 | Poly | Example of SNP located in FEH | |||
| D | H | 1675 | BobWhite_c23193_170 | 4,02,21,884---4,02,21,984 | Poly | ||||||||
| D | H | 41,259 | Kukri_c14765_1655 | 4,10,83,842---4,10,83,933 | Poly | ||||||||
| D | H | 3378 | BobWhite_c44549_83 | 4,10,83,289---4,10,83,389 | Poly | ||||||||
| 6BS | 4,876,122 | D | H | 10,716 | BS00074183_51 | 5,44,19,374---5,44,19,474 | Null | Null SNP near top of del in 426 | |||||
| FEH_6B | D | H | |||||||||||
| d | h | D | H | d | h | 55,811 | RAC875_c25556_1250 | 5,72,96,208---5,72,86,308 | Poly | Example of SNP located in FEH | |||
| D | H | . | . | 75,083 | tplb0055h14_483 | 5,92,95,396---5,92,95,496 | Poly | SNP near bottom of del in 426 | |||||
| 6DS | 1,600,754 | d | h | D | H | 18,485 | D_GBQ4KXB01B5NHZ_336 | 2,86,82,554---2,86,82,785 | Poly | SNP near top of del in 433 | |||
| FEH_6D | |||||||||||||
| d | h | d | h | D | H | 55,811 | RAC875_c25556_1250 | 3,02,82,758---3,02,82,858 | Poly | Example of SNP located in FEH | |||
| s | d | h | d | h | D | H | 46,499 | Kukri_c5531_358 | 3,02,83,208---3,02,83,308 | Poly | SNP near end of del in 433. Located in FEH |
| Major Classification | Chromosome 6A | Chromosome 6B | Chromosome 6D |
|---|---|---|---|
| Hydrolase | TraesCS6A02G048200 TraesCS6A02G042300 TraesCS6A02G042400 TraesCS6A02G060700 (Fructan 1-exohydrolase w1 (Source: UniProtKB/Swiss Prot; Acc: Q84PN8)) TraesCS6A02G060800 TraesCS6A02G069000 | TraesCS6B02G080800 TraesCS6B02G080700 Fructan 1-exohydrolase w3 (Source: UniProtKB/Swiss-Prot; Acc: B6DZC8) | TraesCS6D02G066800 TraesCS6D02G064100 TraesCS6D02G064200 TraesCS6D02G064300 TraesCS6D02G064400 Fructan 1-exohydrolase w2 (Source: UniProtKB/Swiss-Prot; Acc: Q84LA1) |
| Ligase | TraesCS6A02G067500 TraesCS6A02G067600 TraesCS6A02G067700 TraesCS6A02G067800 | TraesCS6D02G065100 TraesCS6D02G065200 | |
| Oxidoreductase | TraesCS6A02G034100 TraesCS6A02G041900 TraesCS6A02G055700 | TraesCS6D02G072600 | |
| Transferase | TraesCS6A02G059300 TraesCS6A02G059700 TraesCS6A02G066000 TraesCS6A02G026800 TraesCS6A02G037400 TraesCS6A02G039500 TraesCS6A02G051200 TraesCS6A02G051300 TraesCS6A02G059700 TraesCS6A02G066000 | TraesCS6B02G079600 TraesCS6B02G080100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, N.; Zhang, J.; Islam, S.; Appels, R.; Dell, B. Wheat Water-Soluble Carbohydrate Remobilisation under Water Deficit by 1-FEH w3. Curr. Issues Mol. Biol. 2023, 45, 6634-6650. https://doi.org/10.3390/cimb45080419
Khan N, Zhang J, Islam S, Appels R, Dell B. Wheat Water-Soluble Carbohydrate Remobilisation under Water Deficit by 1-FEH w3. Current Issues in Molecular Biology. 2023; 45(8):6634-6650. https://doi.org/10.3390/cimb45080419
Chicago/Turabian StyleKhan, Nusrat, Jingjuan Zhang, Shahidul Islam, Rudi Appels, and Bernard Dell. 2023. "Wheat Water-Soluble Carbohydrate Remobilisation under Water Deficit by 1-FEH w3" Current Issues in Molecular Biology 45, no. 8: 6634-6650. https://doi.org/10.3390/cimb45080419
APA StyleKhan, N., Zhang, J., Islam, S., Appels, R., & Dell, B. (2023). Wheat Water-Soluble Carbohydrate Remobilisation under Water Deficit by 1-FEH w3. Current Issues in Molecular Biology, 45(8), 6634-6650. https://doi.org/10.3390/cimb45080419