
Diet Supplementation with Fish-Derived Extracts Suppresses Diabetes and Modulates Intestinal Microbiome in a Murine Model of Diet-Induced Obesity


 , , ,
, , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation of Fish Extracts and Nutritional Supplement Preparation
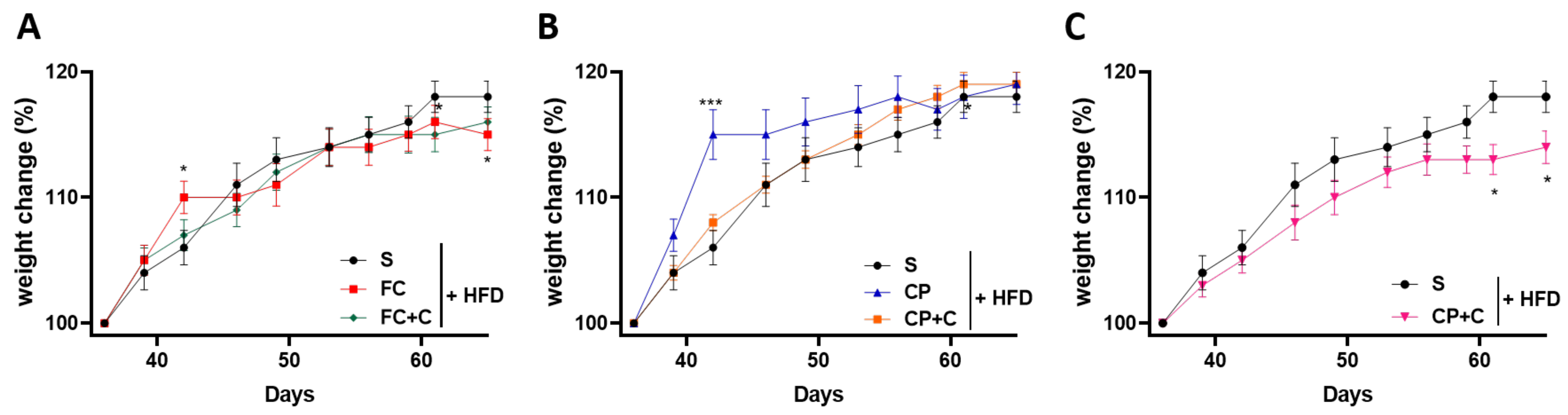
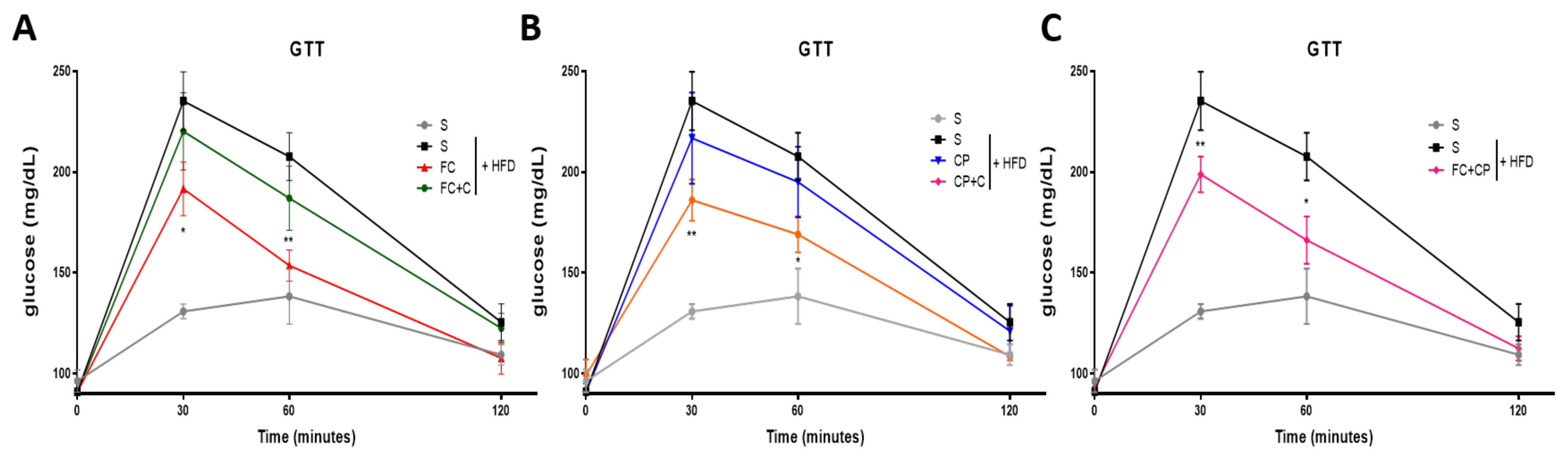
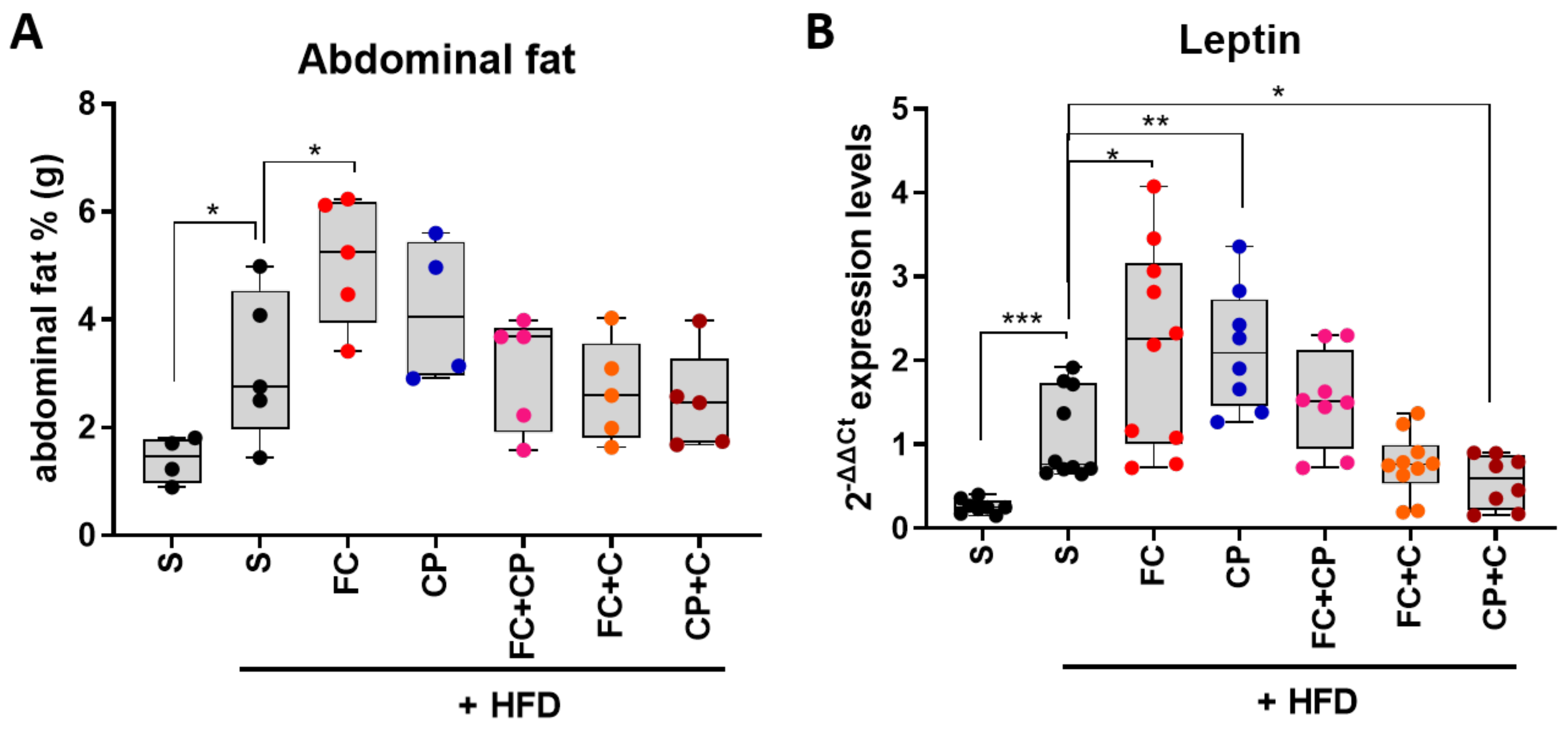
2.2. Fish Supplements Suppress Insulin Resistance in High-Fat Diet Mice Independently of Weight Gain
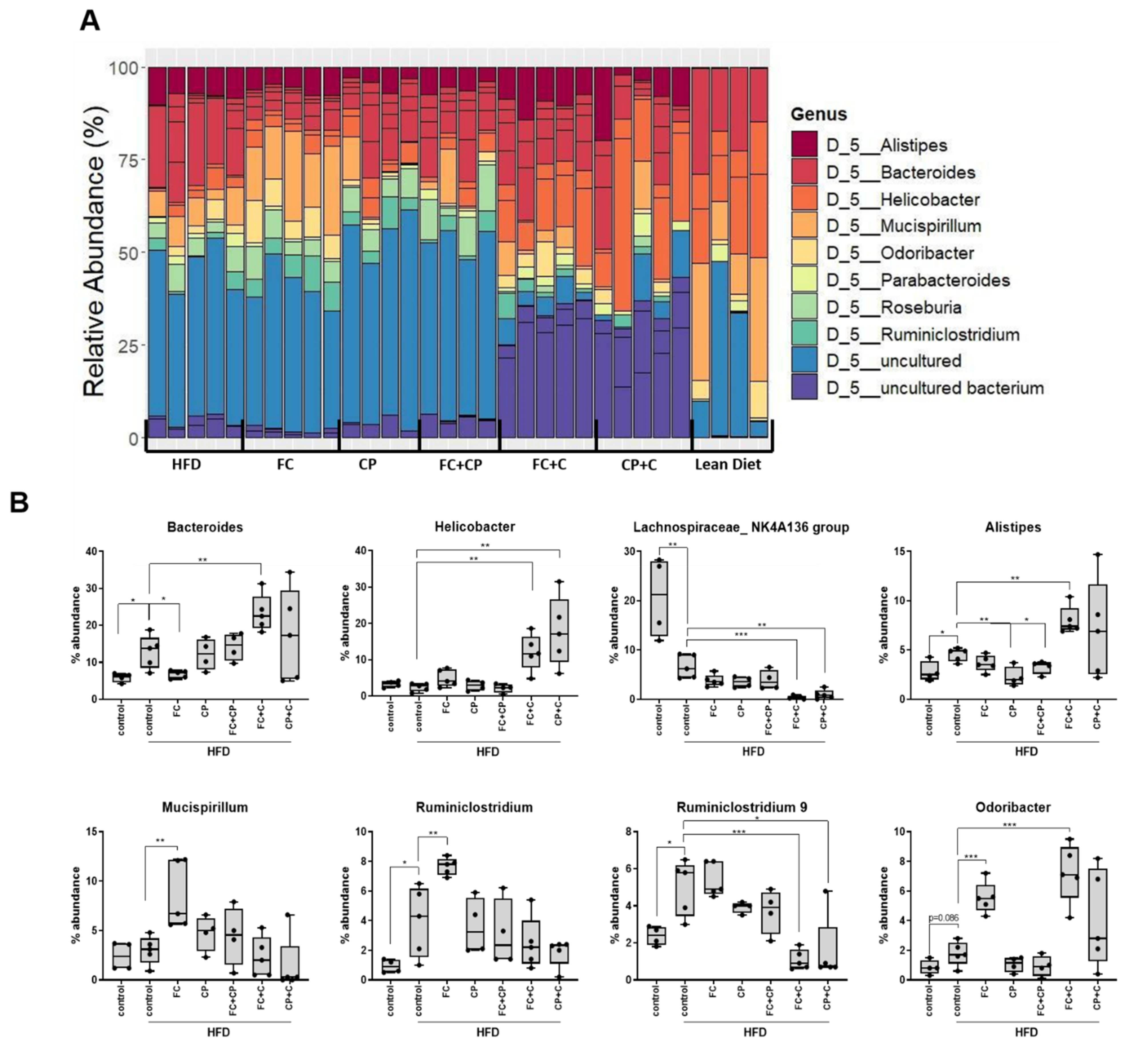
2.3. Fish Supplements Modulate the Gut Microbiome
3. Materials and Methods
3.1. Fish Extracts
3.2. Animal Maintenance-Protocols
3.3. RNA Extraction, cDNA Synthesis and Quantitative PCR
3.4. Microbiome Extraction and Sequencing
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- González-Muniesa, P.; Mártinez-González, M.A.; Hu, F.B.; Després, J.P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Prim. 2017, 3, 17034. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.M.; Olefsky, J.M. The origins and drivers of insulin resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosomworth, N.J. Normal-weight central obesity: Unique hazard of the toxic waist. Can. Fam. Phys. Med. Fam. Can. 2019, 65, 399–408. [Google Scholar]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Isganaitis, E.; Lustig, R.H. Fast food, central nervous system insulin resistance, and obesity. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2451–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivy, J.L. Role of exercise training in the prevention and treatment of insulin resistance and non-insulin-dependent diabetes mellitus. Sports Med. 1997, 24, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Stenvers, D.J.; Scheer, F.; Schrauwen, P.; la Fleur, S.E.; Kalsbeek, A. Circadian clocks and insulin resistance. Nat. Rev. Endocrinol. 2019, 15, 75–89. [Google Scholar] [CrossRef]
- Xu, H.; Li, X.; Adams, H.; Kubena, K.; Guo, S. Etiology of Metabolic Syndrome and Dietary Intervention. Int. J. Mol. Sci. 2018, 20, 128. [Google Scholar] [CrossRef] [Green Version]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Song, G.; Ren, X.; Zhang, L.; Gao, J.; Xia, X.; Zhu, B. Fish oil extracted from Coregonus peled improves obese phenotype and changes gut microbiota in a high-fat diet-induced mouse model of recurrent obesity. Food Funct. 2020, 11, 6158–6169. [Google Scholar] [CrossRef]
- Kim, Y.; Kwon, M.J.; Choi, J.W.; Lee, M.K.; Kim, C.; Jung, J.; Aprianita, H.; Nam, H.; Nam, T.J. Anti-obesity effects of boiled tuna extract in mice with obesity induced by a high-fat diet. Int. J. Mol. Med. 2016, 38, 1281–1288. [Google Scholar] [CrossRef]
- Pradhan, S.; Panchali, T.; Paul, B.; Khatun, A.; Rao Jarapala, S.; Mondal, K.C.; Ghosh, K.; Chakrabarti, S. Anti-obesity potentiality of Tapra fish (Opisthopterus tardoore) oil. J. Food Biochem. 2020, 44, e13448. [Google Scholar] [CrossRef]
- Oh, H.T.; Chung, M.J.; Kim, S.H.; Choi, H.J.; Ham, S.S. Masou salmon (Oncorhynchus masou) ethanol extract decreases 3-hydroxy-3-methylglutaryl coenzyme A reductase expression in diet-induced obese mice. Nutr. Res. 2009, 29, 123–129. [Google Scholar] [CrossRef]
- Astre, G.; Deleruyelle, S.; Dortignac, A.; Bonnet, C.; Valet, P.; Dray, C. Diet-induced obesity and associated disorders are prevented by natural bioactive type 1 fish collagen peptides (Naticol®) treatment. J. Physiol. Biochem. 2018, 74, 647–654. [Google Scholar] [CrossRef]
- Lee, E.J.; Hur, J.; Ham, S.A.; Jo, Y.; Lee, S.; Choi, M.J.; Seo, H.G. Fish collagen peptide inhibits the adipogenic differentiation of preadipocytes and ameliorates obesity in high fat diet-fed mice. Int. J. Biol. Macromol. 2017, 104, 281–286. [Google Scholar] [CrossRef]
- Woo, M.; Song, Y.O. Anti-Obesity Effects of Collagen Peptide Derived from Skate (Raja kenojei) Skin Through Regulation of Lipid Metabolism. Mar. Drugs 2018, 16, 306. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.H.; Chi, C.F.; Zhao, Y.Q.; Wang, B. Preparation, Physicochemical and Antioxidant Properties of Acid- and Pepsin-Soluble Collagens from the Swim Bladders of Miiuy Croaker (Miichthys miiuy). Mar. Drugs 2018, 16, 161. [Google Scholar] [CrossRef] [Green Version]
- Sousa, R.O.; Martins, E.; Carvalho, D.N.; Alves, A.L.; Oliveira, C.; Duarte, A.R.C.; Silva, T.H.; Reis, R.L. Collagen from Atlantic cod (Gadus morhua) skins extracted using CO2 acidified water with potential application in healthcare. J. Polym. Res. 2020, 27, 73. [Google Scholar] [CrossRef] [Green Version]
- Seixas, M.J.; Martins, E.; Reis, R.L.; Silva, T.H. Extraction and Characterization of Collagen from Elasmobranch Byproducts for Potential Biomaterial Use. Mar. Drugs 2020, 18, 617. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.F.; Chang, W.C.; Loh, C.H.; Hsieh, P.S. Leptin-mediated inflammatory signaling crucially links visceral fat inflammation to obesity-associated β-cell dysfunction. Life Sci. 2014, 116, 51–58. [Google Scholar] [CrossRef]
- Rabe, K.; Lehrke, M.; Parhofer, K.G.; Broedl, U.C. Adipokines and insulin resistance. Mol. Med. 2008, 14, 741–751. [Google Scholar] [CrossRef]
- Friedman, J. Fat in all the wrong places. Nature 2002, 415, 268–269. [Google Scholar] [CrossRef]
- Park, H.K.; Ahima, R.S. Physiology of leptin: Energy homeostasis, neuroendocrine function and metabolism. Metab. Clin. Exp. 2015, 64, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, E.; Bettencourt, R. Gene expression study in Bathymodiolus azoricus populations from three North Atlantic hydrothermal vent sites. Dev. Comp. Immunol. 2019, 99, 103390. [Google Scholar] [CrossRef] [PubMed]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome-A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krautkramer, K.A.; Kreznar, J.H.; Romano, K.A.; Vivas, E.I.; Barrett-Wilt, G.A.; Rabaglia, M.E.; Keller, M.P.; Attie, A.D.; Rey, F.E.; Denu, J.M. Diet-Microbiota Interactions Mediate Global Epigenetic Programming in Multiple Host Tissues. Mol. Cell 2016, 64, 982–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieder, R.; Wisniewski, P.J.; Alderman, B.L.; Campbell, S.C. Microbes and mental health: A review. Brain Behav. Immun. 2017, 66, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kong, Q.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. A High-Fat Diet Increases Gut Microbiota Biodiversity and Energy Expenditure Due to Nutrient Difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Link, A.; Selgrad, M. Helicobacter pylori: Perspectives and time trends. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 628–638. [Google Scholar] [CrossRef]
- Ménard, A.; Smet, A. Review: Other Helicobacter species. Helicobacter 2019, 24 (Suppl. 1), e12645. [Google Scholar] [CrossRef] [Green Version]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhi, F. Lower Level of Bacteroides in the Gut Microbiota Is Associated with Inflammatory Bowel Disease: A Meta-Analysis. BioMed. Res. Int. 2016, 2016, 5828959. [Google Scholar] [CrossRef] [Green Version]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Göker, M.; Gronow, S.; Zeytun, A.; Nolan, M.; Lucas, S.; Lapidus, A.; Hammon, N.; Deshpande, S.; Cheng, J.-F.; Pitluck, S.; et al. Complete genome sequence of Odoribacter splanchnicus type strain (1651/6). Stand. Genomic Sci. 2011, 4, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2014. Available online: http://www.r-project.org/ (accessed on 3 February 2021).
- McMurdie, P.J.; Holmes, S. Phyloseq: A bioconductor package for handling and analysis of high-throughput phylogenetic sequence data. Pac. Symp. Biocomput. Pac. Symp. Biocomput. 2012, 2012, 235–246. [Google Scholar]
- Albertsen, M.; Karst, S.M.; Ziegler, A.S.; Kirkegaard, R.H.; Nielsen, P.H. Back to Basics—The Influence of DNA Extraction and Primer Choice on Phylogenetic Analysis of Activated Sludge Communities. PLoS ONE 2015, 10, e0132783. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin, Germany, 2016; pp. 187–253. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Supplement | Composition |
|---|---|---|
| Standard | S | Soy protein |
| High-fat | S | Soy protein |
| FC | White fish powder | |
| CP | Cod powder | |
| FC + CP | 50%/50% White fish powder + cod powder | |
| FC + C | 50%/50% White fish powder + fish skin protein powder | |
| CP +C | 50%/50% Cod powder + fish skin protein powder |
| Chemical Composition | Unit | Collagen | Fish Complex | Cod Powder | ||
|---|---|---|---|---|---|---|
| Energy | Kcal/100 g extract | 389 | 304 | 385 | ||
| KJ | 1634 | 1274 | 1620 | |||
| Total solids | g/100 g extract | ≥92 | 94 | 94 | ||
| Protein | g/100 g extract | ≥95 | 66 | 87 | ||
| Ash | g/100 g extract | <2 | 22 | 7 | ||
| Salt | g/100 g extract | <1 | 1.2 | 2 | ||
| Fat | g/100 g extract | <0.5 | 4.4 | 4 | ||
| Saturated | 1.15 | 0.9 | ||||
| Monounsaturated | 2.08 | 0.9 | ||||
| Polyunsaturated | 1.74 | 1.8 | ||||
| Omega-6 | 0.2 | 0.2 | ||||
| Linoleic acid | 0.08 | 0.11 | ||||
| Omega-3 | 1.5 | 1.6 | ||||
| DHA | 0.87 | 0.97 | ||||
| EPA | 0.43 | 0.49 |
| Minerals | Unit | Collagen | Fish Complex | Cod Powder |
|---|---|---|---|---|
| Calcium | mg/100 g | not detected | 6810 | 934 |
| Phosphorus | mg/100 g | not detected | 4890 | 993 |
| Sodium | mg/100 g | not detected | 730 | 800 |
| Potassium | mg/100 g | not detected | 571 | 1360 |
| Magnesium | mg/100 g | not detected | 215 | 120 |
| Iron | mg/100 g | not detected | 20.8 | 1.1 |
| Zink | mg/100 g | not detected | 7.85 | 2.71 |
| Iodine | mg/100 g | not detected | 0.43 | 0.59 |
| Selenium | mg/100 g | not detected | 0.22 | 0.12 |
| Vitamin B12 | mg/100 g | not detected | 0.0067 | 0.0069 |
| Microorganism | Unit | Collagen | Fish Complex | Cod Powder |
|---|---|---|---|---|
| Total plate count | Max CFU/g | <1000 | 50,000 | 50,000 |
| Enterobacteriaceae | Max CFU/g | <100 | 100 | 100 |
| Escherichia coli | Max CFU/g | Negative in 1 g | Negative in 1 g | <10 |
| Salmonella | Max CFU/g | Negative in 25 g | Negative in 25 g | Negative in 25 g |
| Mold and yeast | Max CFU/g | <200 | 500 | 1000 |
| Sulphite-Reducing Clostridia | Max CFU/g | <100 | <100 | - |
| Listeria monocytogenes | Max CFU/g | Negative in 25 g | Negative 25 g | Negative in 25 g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Axarlis, K.; Daskalaki, M.G.; Michailidou, S.; Androulaki, N.; Tsoureki, A.; Mouchtaropoulou, E.; Kolliniati, O.; Lapi, I.; Dermitzaki, E.; Venihaki, M.; et al. Diet Supplementation with Fish-Derived Extracts Suppresses Diabetes and Modulates Intestinal Microbiome in a Murine Model of Diet-Induced Obesity. Mar. Drugs 2021, 19, 268. https://doi.org/10.3390/md19050268
Axarlis K, Daskalaki MG, Michailidou S, Androulaki N, Tsoureki A, Mouchtaropoulou E, Kolliniati O, Lapi I, Dermitzaki E, Venihaki M, et al. Diet Supplementation with Fish-Derived Extracts Suppresses Diabetes and Modulates Intestinal Microbiome in a Murine Model of Diet-Induced Obesity. Marine Drugs. 2021; 19(5):268. https://doi.org/10.3390/md19050268
Chicago/Turabian StyleAxarlis, Konstantinos, Maria G. Daskalaki, Sofia Michailidou, Nikolais Androulaki, Antiopi Tsoureki, Evangelia Mouchtaropoulou, Ourania Kolliniati, Ioanna Lapi, Eirini Dermitzaki, Maria Venihaki, and et al. 2021. "Diet Supplementation with Fish-Derived Extracts Suppresses Diabetes and Modulates Intestinal Microbiome in a Murine Model of Diet-Induced Obesity" Marine Drugs 19, no. 5: 268. https://doi.org/10.3390/md19050268
APA StyleAxarlis, K., Daskalaki, M. G., Michailidou, S., Androulaki, N., Tsoureki, A., Mouchtaropoulou, E., Kolliniati, O., Lapi, I., Dermitzaki, E., Venihaki, M., Kousoulaki, K., Argiriou, A., Marsni, Z. E., & Tsatsanis, C. (2021). Diet Supplementation with Fish-Derived Extracts Suppresses Diabetes and Modulates Intestinal Microbiome in a Murine Model of Diet-Induced Obesity. Marine Drugs, 19(5), 268. https://doi.org/10.3390/md19050268