


High-Cell-Density Yeast Oil Production with Diluted Substrates Imitating Microalgae Hydrolysate Using a Membrane Bioreactor

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain and Cultivation Medium
2.2. Membrane Bioreactor with Total Cell Retention
2.3. Semi-Continuous Operation of the MBR
2.4. Optical Density and Cell Dry Weight
2.5. Total Lipid Analysis
2.6. Fatty Acid Composition
2.7. Substrate Anaylsis
2.8. Elemental Analysis
2.9. Carbon Balance and Conversion Efficiency
3. Results and Discussion
3.1. High-Cell-Density Yeast Oil Production in the MBR
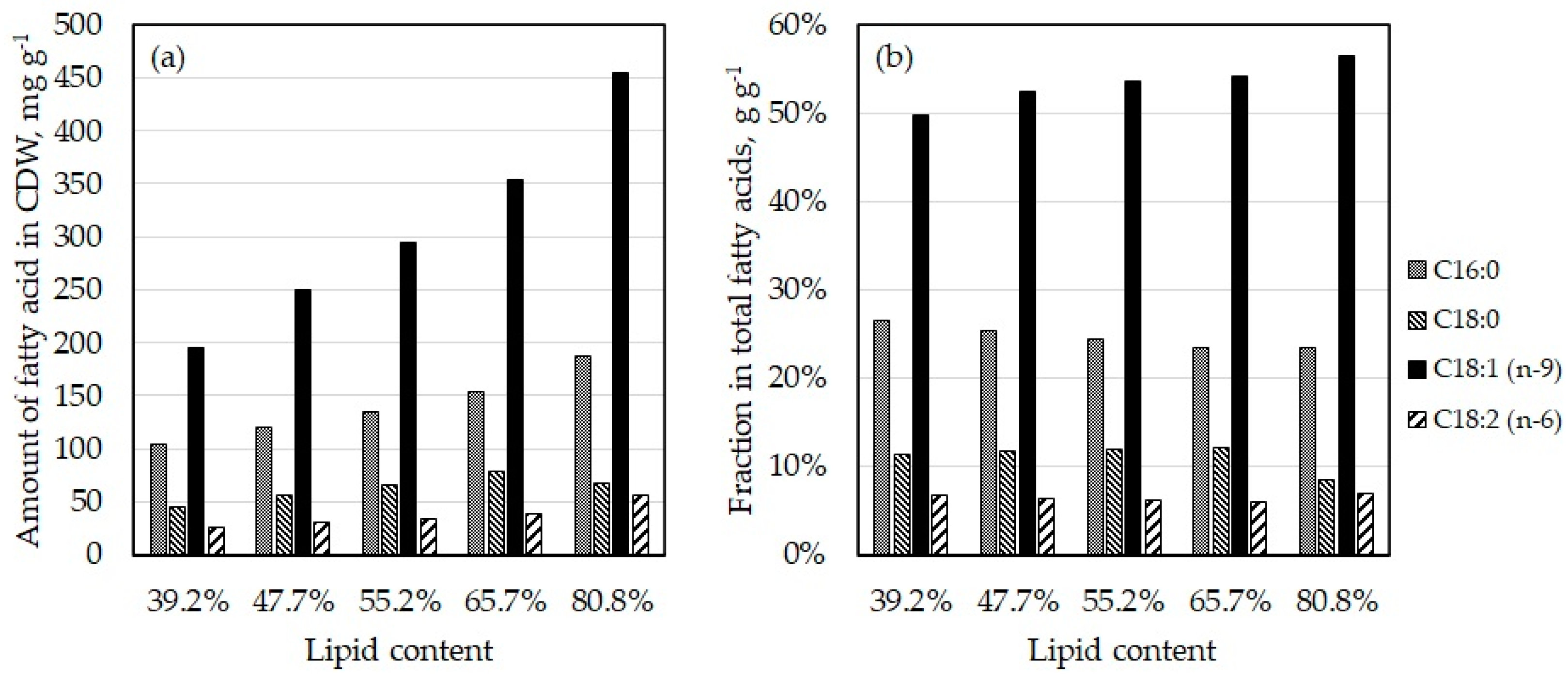
3.2. Fatty Acid Composition
3.3. Elemental Composition
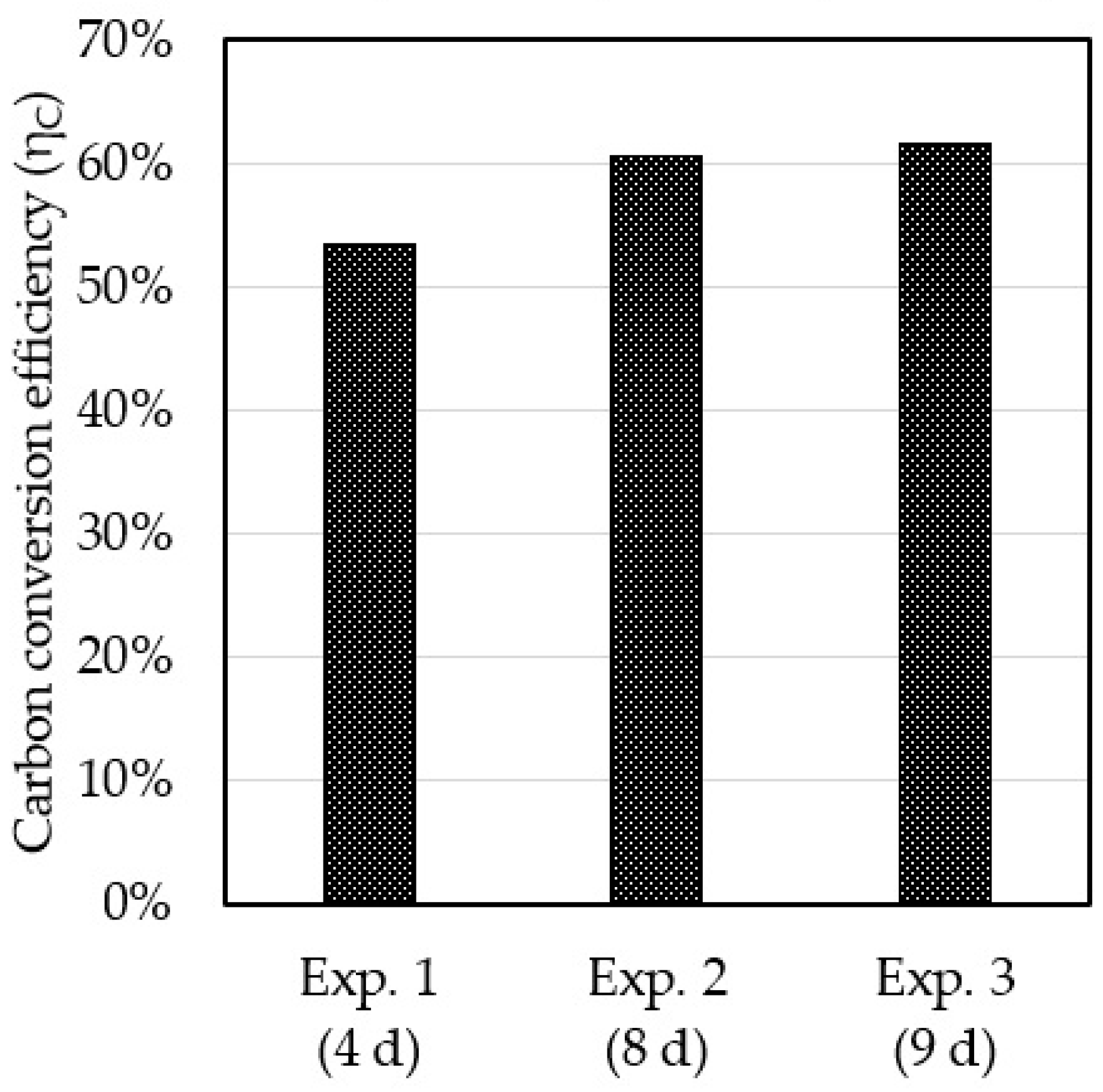
3.4. Carbon Balance and Conversion Efficiency
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green Deal: EU Agrees Law to Fight Global Deforestation and Forest Degradation Driven by EU Production and Consumption. Available online: https://ec.europa.eu/commission/presscorner/detail/en/ip_22_7444 (accessed on 30 January 2023).
- The Paris Agreement. Available online: https://unfccc.int/process-and-meetings/the-paris-agreement/the-paris-agreement (accessed on 30 January 2023).
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Ratledge, C. Single cell oils—Have they a biotechnological future? Trends Biotechnol. 1993, 11, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Masri, M.A.; Garbe, D.; Mehlmer, N.; Brück, T. A sustainable, high-performance process for the economic production of waste-free microbial oils that can replace plant-based equivalents. Energy Environ. Sci. 2019, 12, 2717–2732. [Google Scholar] [CrossRef]
- Thorpe, R.F.; Ratledge, C. Fatty Acid Distribution in Triglycerides of Yeasts Grown on Glucose or n-Alkanes. J. Gen. Microbiol. 1972, 72, 151–163. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The history, state of the art and future prospects for oleaginous yeast research. Microb. Cell Fact. 2021, 20, 221. [Google Scholar] [CrossRef] [PubMed]
- Angerbauer, C.; Siebenhofer, M.; Mittelbach, M.; Guebitz, G.M. Conversion of sewage sludge into lipids by Lipomyces starkeyi for biodiesel production. Bioresour. Technol. 2008, 99, 3051–3056. [Google Scholar] [CrossRef]
- Xue, F.; Miao, J.; Zhang, X.; Luo, H.; Tan, T. Studies on lipid production by Rhodotorula glutinis fermentation using monosodium glutamate wastewater as culture medium. Bioresour. Technol. 2008, 99, 5923–5927. [Google Scholar] [CrossRef]
- Yousuf, A.; Sannino, F.; Addorisio, V.; Pirozzi, D. Microbial Conversion of Olive Oil Mill Wastewaters into Lipids Suitable for Biodiesel Production. J. Agric. Food Chem. 2010, 58, 8630–8635. [Google Scholar] [CrossRef]
- Shaigani, P.; Awad, D.; Redai, V.; Fuchs, M.; Haack, M.; Mehlmer, N.; Brueck, T. Oleaginous yeasts-substrate preference and lipid productivity: A view on the performance of microbial lipid producers. Microb. Cell Fact. 2021, 20, 220. [Google Scholar] [CrossRef]
- Grubišić, M.; Perečinec, M.G.; Peremin, I.; Mihajlovski, K.; Beluhan, S.; Šantek, B.; Šantek, M.I. Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus. Energies 2022, 15, 3208. [Google Scholar] [CrossRef]
- Grubišić, M.; Mihajlovski, K.; Gruičić, A.M.; Beluhan, S.; Šantek, B.; Šantek, M.I. Strategies for Improvement of Lipid Production by Yeast Trichosporon oleaginosus from Lignocellulosic Biomass. J. Fungi 2021, 7, 934. [Google Scholar] [CrossRef] [PubMed]
- Moon, N.J.; Hammond, E.G.; Glatz, B.A. Conversion of Cheese Whey and Whey Permeate to Oil and Single-Cell Protein. J. Dairy Sci. 1978, 61, 1537–1547. [Google Scholar] [CrossRef]
- Masri, M.A.; Younes, S.; Haack, M.; Qoura, F.; Mehlmer, N.; Brück, T. A Seagrass-Based Biorefinery for Generation of Single-Cell Oils for Biofuel and Oleochemical Production. Energy Technol. 2018, 6, 1026–1038. [Google Scholar] [CrossRef]
- Younes, S.; Bracharz, F.; Awad, D.; Qoura, F.; Mehlmer, N.; Brueck, T. Microbial lipid production by oleaginous yeasts grown on Scenedesmus obtusiusculus microalgae biomass hydrolysate. Bioprocess Biosyst. Eng. 2020, 43, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Di Fidio, N.; Minonne, F.; Antonetti, C.; Galletti, A.M.R. Cutaneotrichosporon oleaginosus: A Versatile Whole-Cell Biocatalyst for the Production of Single-Cell Oil from Agro-Industrial Wastes. Catalysts 2021, 11, 1291. [Google Scholar] [CrossRef]
- Schädler, T.; Cerbon, D.C.; de Oliveira, L.; Garbe, D.; Brück, T.; Weuster-Botz, D. Production of lipids with Microchloropsis salina in open thin-layer cascade photobioreactors. Bioresour. Technol. 2019, 289, 121682. [Google Scholar] [CrossRef]
- Brenner, A.; Abeliovich, A. Water Purification: Algae in Wastewater Oxidation Ponds. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 595–601. [Google Scholar]
- Kaplan, D. Absorption and Adsorption of Heavy Metals by Microalgae. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 602–611. [Google Scholar]
- Becker, E.W. Microalgae for Aquaculture: Nutritional Aspects. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 671–691. [Google Scholar]
- Muller-Feuga, A. Microalgae for Aquaculture: The Current Global Situation and Future Trends. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 613–627. [Google Scholar]
- Ratledge, C. Single Cell Oils for the 21st Century. In Single Cell Oils: Microbial and Algal Oils; AOCS Publishing: New York, NY, USA, 2005; pp. 1–20. [Google Scholar]
- Meo, A.; Priebe, X.L.; Weuster-Botz, D. Lipid production with Trichosporon oleaginosus in a membrane bioreactor using microalgae hydrolysate. J. Biotechnol. 2017, 241, 1–10. [Google Scholar] [CrossRef]
- Kourist, R.; Bracharz, F.; Lorenzen, J.; Kracht, O.N.; Chovatia, M.; Daum, C.; Deshpande, S.; Lipzen, A.; Nolan, M.; Ohm, R.A.; et al. Genomics and Transcriptomics Analyses of the Oil-Accumulating Basidiomycete Yeast Trichosporon oleaginosus: Insights into Substrate Utilization and Alternative Evolutionary Trajectories of Fungal Mating Systems. mBio 2015, 6, e00918-15. [Google Scholar] [CrossRef]
- Yaguchi, A.; Rives, D.; Blenner, M. New kids on the block: Emerging oleaginous yeast of biotechnological importance. AIMS Microbiol. 2017, 3, 227–247. [Google Scholar] [CrossRef]
- Karayannis, D.; Papanikolaou, S.; Vatistas, C.; Paris, C.; Chevalot, I. Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants. Int. J. Mol. Sci. 2023, 24, 714. [Google Scholar] [CrossRef]
- Ykema, A.; Verbree, E.C.; Kater, M.M.; Smit, H. Optimization of lipid production in the oleaginous yeast Apiotrichum curvatum in wheypermeate. Appl. Microbiol. Biotechnol. 1988, 29, 211–218. [Google Scholar] [CrossRef]
- Yaguchi, A.; Robinson, A.; Mihealsick, E.; Blenner, M. Metabolism of aromatics by Trichosporon oleaginosus while remaining oleaginous. Microb. Cell Fact. 2017, 16, 206. [Google Scholar] [CrossRef] [PubMed]
- Bracharz, F.; Beukhout, T.; Mehlmer, N.; Brück, T. Opportunities and challenges in the development of Cutaneotrichosporon oleaginosus ATCC 20509 as a new cell factory for custom tailored microbial oils. Microb. Cell Fact. 2017, 16, 178. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zeng, J.; Zheng, Y.; Chen, S. Effect of lignocellulose degradation products on microbial biomass and lipid production by the oleaginous yeast Cryptococcus curvatus. Process Biochem. 2014, 49, 457–465. [Google Scholar] [CrossRef]
- Yu, X.; Zheng, Y.; Dorgan, K.M.; Chen, S. Oil production by oleaginous yeasts using the hydrolysate from pretreatment of wheat straw with dilute sulfuric acid. Bioresour. Technol. 2011, 102, 6134–6140. [Google Scholar] [CrossRef]
- Sabelfeld, M.; Geißen, S.-U. Processes for the Elimination and Recovery of Phosphorus from Municipal Wastewater. Chem. Ing. Tech. 2011, 83, 782–795. [Google Scholar] [CrossRef]
- Hassan, M.; Blanc, P.J.; Granger, L.-M.; Pareilleux, A.; Goma, G. Influence of nitrogen and iron limitations on lipid production by Cryptococcus curvatus grown in batch and fed-batch culture. Process Biochem. 1996, 31, 355–361. [Google Scholar] [CrossRef]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.-W. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Griffiths, M.J.; van Hille, R.P.; Harrison, S.T.L. Selection of Direct Transesterification as the Preferred Method for Assay of Fatty Acid Content of Microalgae. Lipids 2010, 45, 1053–1060. [Google Scholar] [CrossRef]
- Thurn, A.L.; Stock, A.; Gerwald, S.; Weuster-Botz, D. Simultaneous photoautotrophic production of DHA and EPA by Tisochrysis lutea and Microchloropsis salina in co-culture. Bioresour. Bioprocess. 2022, 9, 130. [Google Scholar] [CrossRef]
- Zhang, G.; French, W.T.; Hernandez, R.; Hall, J.; Sparks, D.; Holmes, W. Microbial lipid production as biodiesel feedstock from N-acetylglucosamine by oleaginous microorganisms. J. Chem. Technol. Biotechnol. 2011, 86, 642–650. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijberts, G.N.M.; Eggink, G. High-cell-density cultivation of the lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Zhou, W.; Tang, M.; Zou, T.; Peng, N.; Zhao, M.; Gong, Z. Phosphate removal combined with acetate supplementation enhances lipid production from water hyacinth by Cutaneotrichosporon oleaginosum. Biotechnol. Biofuels 2019, 12, 148. [Google Scholar] [CrossRef] [PubMed]
- Awad, D.; Bohnen, F.; Mehlmer, N.; Brück, T. Multi-Factorial-Guided Media Optimization for Enhanced Biomass and Lipid Formation by the Oleaginous Yeast Cutaneotrichosporon oleaginosus. Front. Bioeng. Biotechnol. 2019, 7, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Lehrbuch der Lebensmittelchemie, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 659–690. [Google Scholar]
- Minkevich, I.G.; Dedyukhina, E.G.; Chistyakova, T.I. The effect of lipid content on the elemental composition and energy capacity of yeast biomass. Appl. Microbiol. Biotechnol. 2010, 88, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, Y.; Zhang, Y.; Zhao, Z.K. Energy efficiency evaluation of lipid production by oleaginous yeast Rhodosporidium toruloides. J. Therm. Anal. Calorim. 2012, 108, 119–126. [Google Scholar] [CrossRef]
- von Stockar, U.; Liu, J.S. Does microbial life always feed on negative entropy? Thermodynamic analysis of microbial growth. Biochim. Biophys. Acta Bioenerg. 1999, 1412, 191–211. [Google Scholar] [CrossRef]
- Kruger, J.S.; Cleveland, N.S.; Yeap, R.Y.; Dong, T.; Ramirez, K.J.; Nagle, N.J.; Lowell, A.C.; Beckham, G.T.; McMillan, J.D.; Biddy, M.J. Recovery of Fuel-Precursor Lipids from Oleaginous Yeast. ACS Sustain. Chem. Eng. 2018, 6, 2921–2931. [Google Scholar] [CrossRef]
- Khot, M.; Raut, G.; Ghosh, D.; Alarcón-Vivero, M.; Contreras, D.; Ravikumar, A. Lipid recovery from oleaginous yeasts: Perspectives and challenges for industrial applications. Fuel 2020, 259, 116292. [Google Scholar] [CrossRef]
- Scholz, M.J.; Weiss, T.L.; Jinkerson, R.E.; Jing, J.; Roth, R.; Goodenough, U.; Posewitz, M.C.; Gerken, H.G. Ultrastructure and Composition of the Nannochloropsis gaditana Cell Wall. Eukaryot. Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef] [Green Version]
- Demuez, M.; Mahdy, A.; Tomás-Pejó, E.; González-Fernández, C.; Ballesteros, M. Enzymatic cell disruption of microalgae biomass in biorefinery processes. Biotechnol. Bioeng. 2015, 112, 1955–1966. [Google Scholar] [CrossRef] [PubMed]

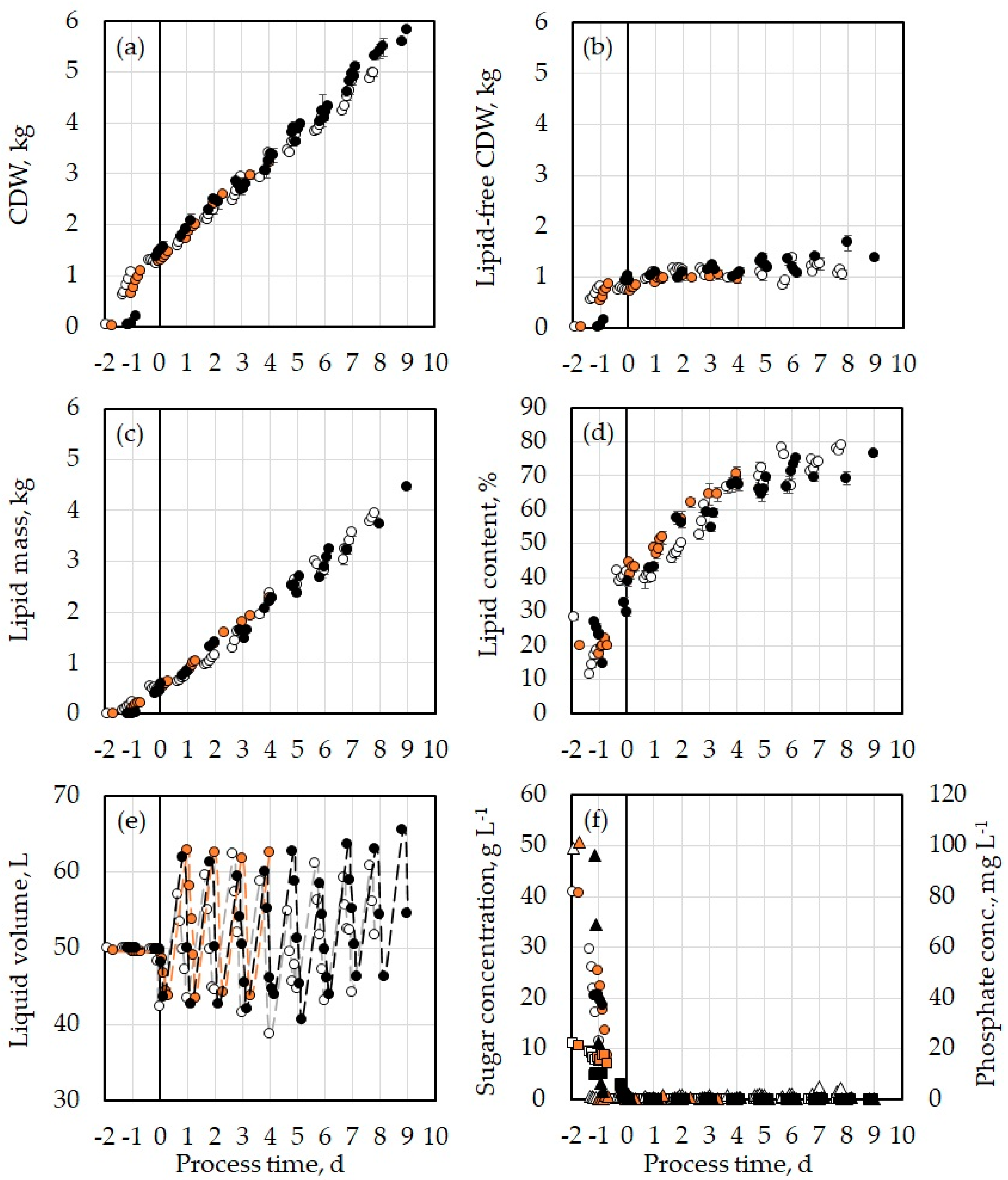
 ), those of Experiment 2 in white (
), those of Experiment 2 in white ( ), and those of Experiment 3 in black (
), and those of Experiment 3 in black ( ). The error bars show the standard deviation of the triplicate measurements. Presented are: (a) cell dry weight; (b) lipid-free cell dry weight; (c) lipid mass; (d) lipid content of the yeast cells; (e) liquid volume inside the MBR; (f) concentrations of glucose (●) and mannose (■) on the primary vertical axis and phosphate (▲) concentration on the secondary vertical axis with the corresponding marker color of each experiment.
), those of Experiment 2 in white (), and those of Experiment 3 in black (). The error bars show the standard deviation of the triplicate measurements. Presented are: (a) cell dry weight; (b) lipid-free cell dry weight; (c) lipid mass; (d) lipid content of the yeast cells; (e) liquid volume inside the MBR; (f) concentrations of glucose (●) and mannose (■) on the primary vertical axis and phosphate (▲) concentration on the secondary vertical axis with the corresponding marker color of each experiment.
). The error bars show the standard deviation of the triplicate measurements. Presented are: (a) cell dry weight; (b) lipid-free cell dry weight; (c) lipid mass; (d) lipid content of the yeast cells; (e) liquid volume inside the MBR; (f) concentrations of glucose (●) and mannose (■) on the primary vertical axis and phosphate (▲) concentration on the secondary vertical axis with the corresponding marker color of each experiment.
), those of Experiment 2 in white (), and those of Experiment 3 in black (). The error bars show the standard deviation of the triplicate measurements. Presented are: (a) cell dry weight; (b) lipid-free cell dry weight; (c) lipid mass; (d) lipid content of the yeast cells; (e) liquid volume inside the MBR; (f) concentrations of glucose (●) and mannose (■) on the primary vertical axis and phosphate (▲) concentration on the secondary vertical axis with the corresponding marker color of each experiment.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process Parameter | Value |
|---|---|
| Reactor type | Stirred-tank bioreactor |
| Agitator type | 2 × Rushton turbines |
| Working volume including bypass | 40–67 L |
| Bypass volume | 7.25 L |
| Temperature | 30 °C |
| pH | 6.5 |
| Aeration rate | 50–100 NL min−1 |
| Agitation | 350–800 rpm |
| Cross-flow filter type | Microfilter with capillaries |
| Cross-flow filter material | Ceramic (>99% SIC) |
| Cross-flow filter pore size | 0.25 μm |
| Volumetric flow rate in bypass | 150 L h−1 |
| Mean residence time in bypass | 2.9 min |
| Fraction in Total Fatty Acids, % (w/w) | |||||||
|---|---|---|---|---|---|---|---|
| Fatty Acid | Trivial Name | Exp. 1 | Exp. 2 | Meo et al. [24] | Zhang et al. [38] | Olive oil [42] | Palm oil [42] |
| C14:0 | Myristic acid | 0.6 | 0.6 | 0.7 | 1.9 | 0 | 1 |
| C16:0 | Palmitic acid | 23.1 | 23.5 | 25.9 | 32.0 | 11.5 | 43.8 |
| C16:1 | Palmitoleic acid | 0.5 | 0.7 | - | 1.3 | 1.5 | 0.5 |
| C18:0 | Stearic acid | 11.8 | 8.4 | 15.4 | 10.2 | 2.5 | 5 |
| C18:1 (n-9) | Oleic acid | 52.8 | 56.4 | 53.6 | 44.9 | 75.5 | 39 |
| C18:2 (n-6) | Linoleic acid | 5.8 | 7.1 | 3.6 | 5.0 | 7.5 | 10 |
| C18:3 (n-3) | α-Linolenic acid | 0.5 | 0.4 | 0.9 | 0.9 | 1.0 | 0.2 |
| C20:0 | Arachidic acid | 0.4 | 0.3 | - | - | 0.5 | 0.5 |
| C22:0 | Behenic acid | 0.2 | 0.2 | - | - | - | |
| C24:0 | Lignoceric acid | 3.1 | 1.4 | - | - | - | |
| C22:6 (n-3) | DHA | 0.6 | 0.6 | - | - | - | |
| C24:1 (n-9) | Nervonic acid | 0.3 | 0.0 | - | - | - | |
| Others | 0.2 | 0.3 | - | - | - | ||
| Mass Fraction of the Element, % | ||||||
|---|---|---|---|---|---|---|
| C | H | N | S | O | P | |
| Dry yeast cells (14.8% (w/w) lipids) | 50.42 | 7.44 | 9.39 | 0.23 | 31.62 | 0.91 |
| Yeast oil | 76.21 | 12.63 | 0.03 | 0.00 | 11.05 | 0.06 |
| Lipid-free yeast dry mass (calculated) | 45.94 | 6.54 | 11.01 | 0.26 | 35.19 | 1.06 |
| Fraction of Total Carbon Consumed Converted into, % | |||
|---|---|---|---|
| Lipid-Free CDW | Lipids | CO2 | |
| Exp. 1 (4 d) | 13.4 | 53.3 | 33.3 |
| Exp. 2 (8 d) | 9.6 | 60.3 | 30.1 |
| Exp. 3 (9 d) | 11.4 | 61.3 | 27.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koruyucu, A.; Blums, K.; Peest, T.; Schmack-Rauscher, L.; Brück, T.; Weuster-Botz, D. High-Cell-Density Yeast Oil Production with Diluted Substrates Imitating Microalgae Hydrolysate Using a Membrane Bioreactor. Energies 2023, 16, 1757. https://doi.org/10.3390/en16041757
Koruyucu A, Blums K, Peest T, Schmack-Rauscher L, Brück T, Weuster-Botz D. High-Cell-Density Yeast Oil Production with Diluted Substrates Imitating Microalgae Hydrolysate Using a Membrane Bioreactor. Energies. 2023; 16(4):1757. https://doi.org/10.3390/en16041757
Chicago/Turabian StyleKoruyucu, Ayşe, Karlis Blums, Tillmann Peest, Laura Schmack-Rauscher, Thomas Brück, and Dirk Weuster-Botz. 2023. "High-Cell-Density Yeast Oil Production with Diluted Substrates Imitating Microalgae Hydrolysate Using a Membrane Bioreactor" Energies 16, no. 4: 1757. https://doi.org/10.3390/en16041757
APA StyleKoruyucu, A., Blums, K., Peest, T., Schmack-Rauscher, L., Brück, T., & Weuster-Botz, D. (2023). High-Cell-Density Yeast Oil Production with Diluted Substrates Imitating Microalgae Hydrolysate Using a Membrane Bioreactor. Energies, 16(4), 1757. https://doi.org/10.3390/en16041757