
Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids


Abstract
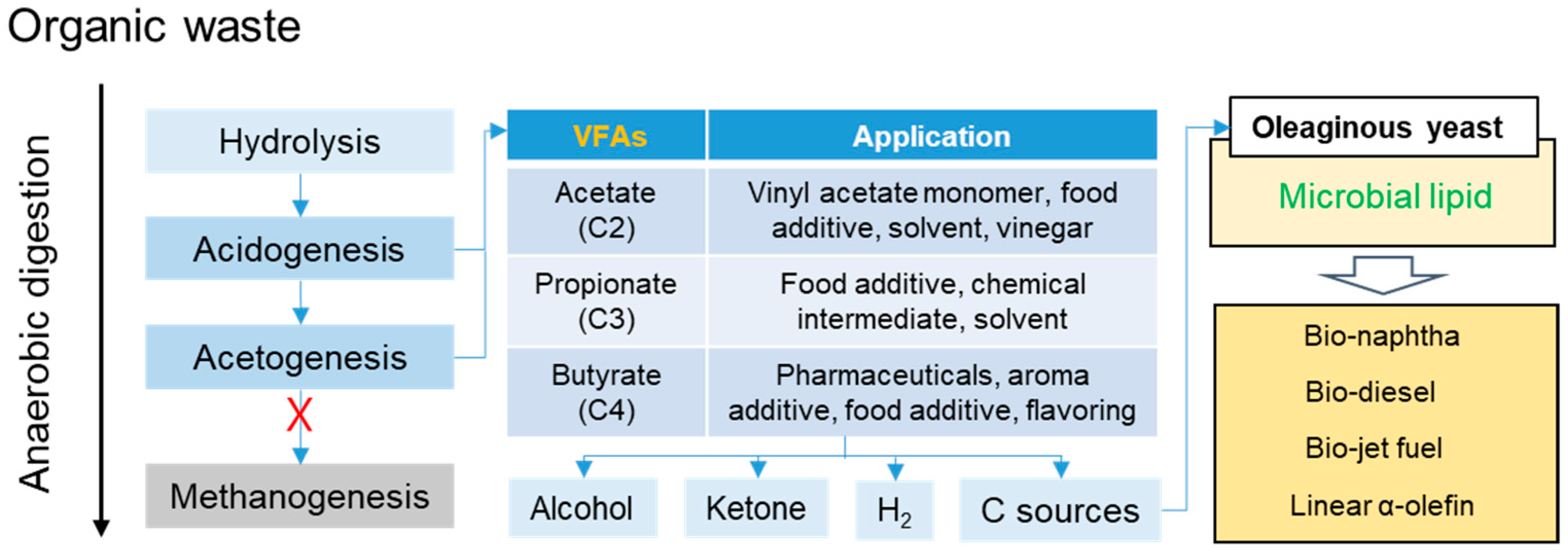
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Flask Culture Condition
2.3. Fed-Batch Cultivation in 5 L Fermenter
2.4. Analysis
3. Results and Discussion
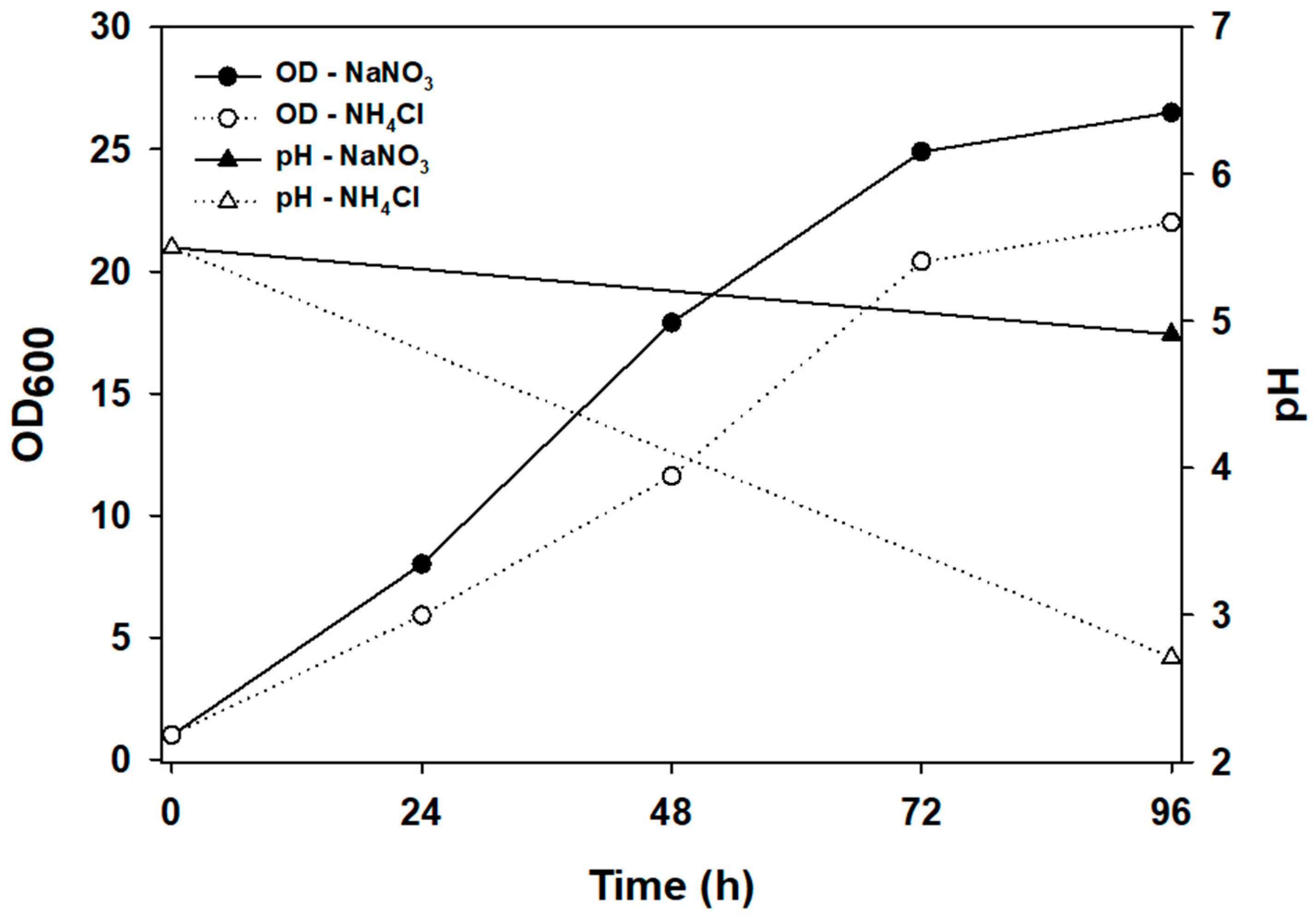
3.1. Effect of Nitrogen Source on pH of the Medium in Flask Culture
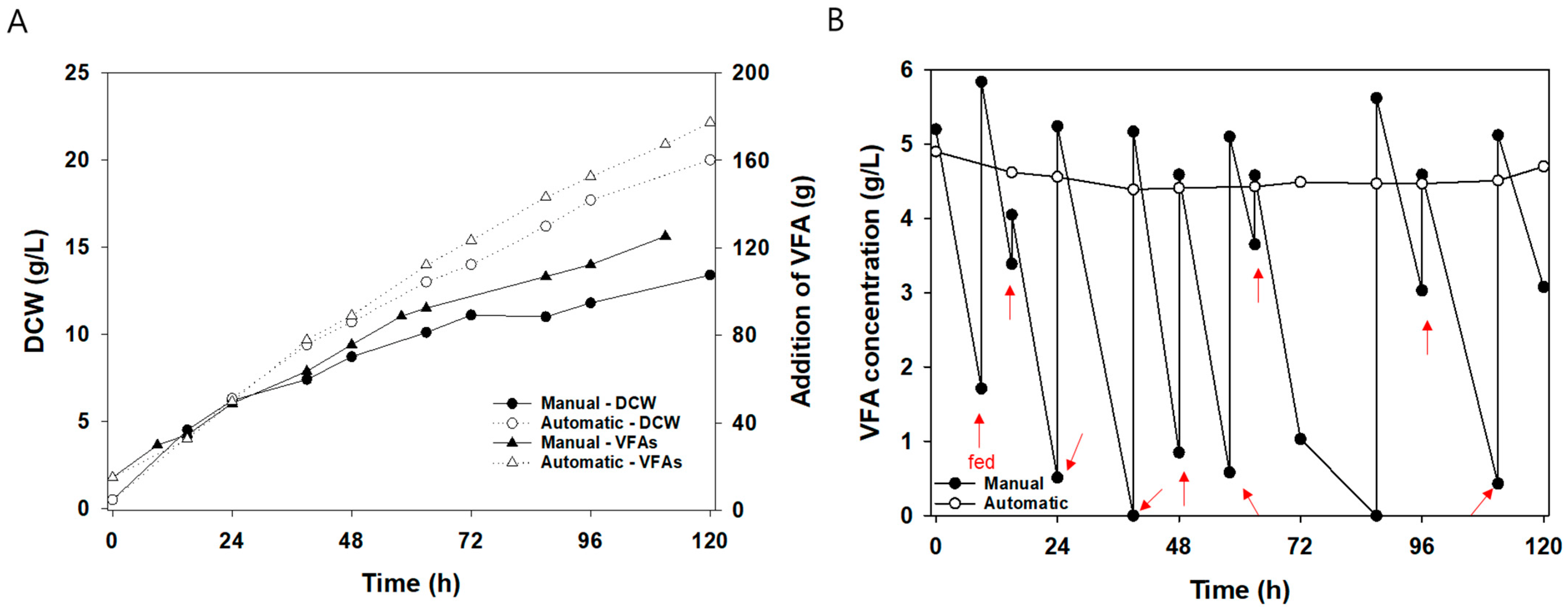
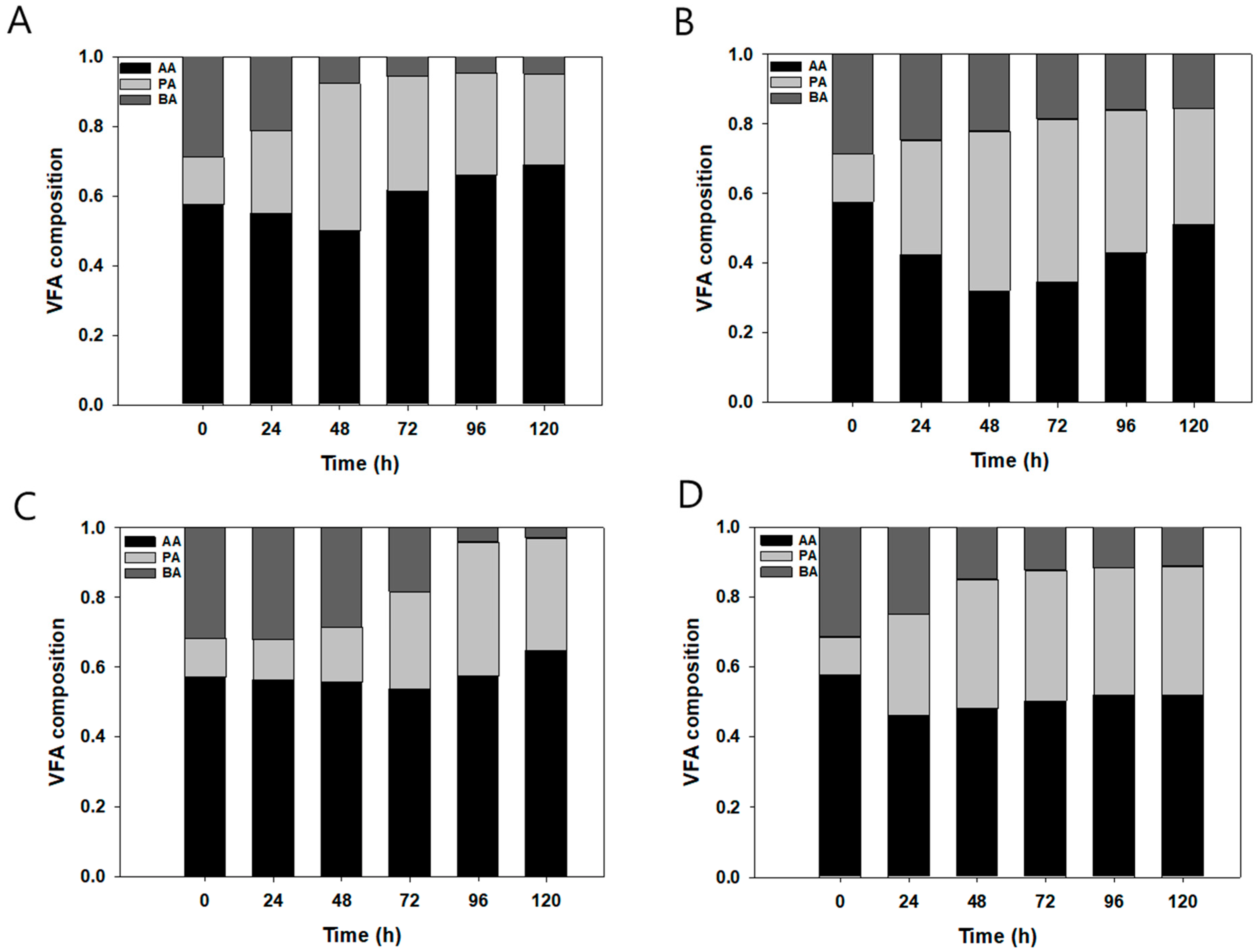
3.2. Yeast Cultivation with Manual and Automatic Fed-Batch in 5 L Fermenter
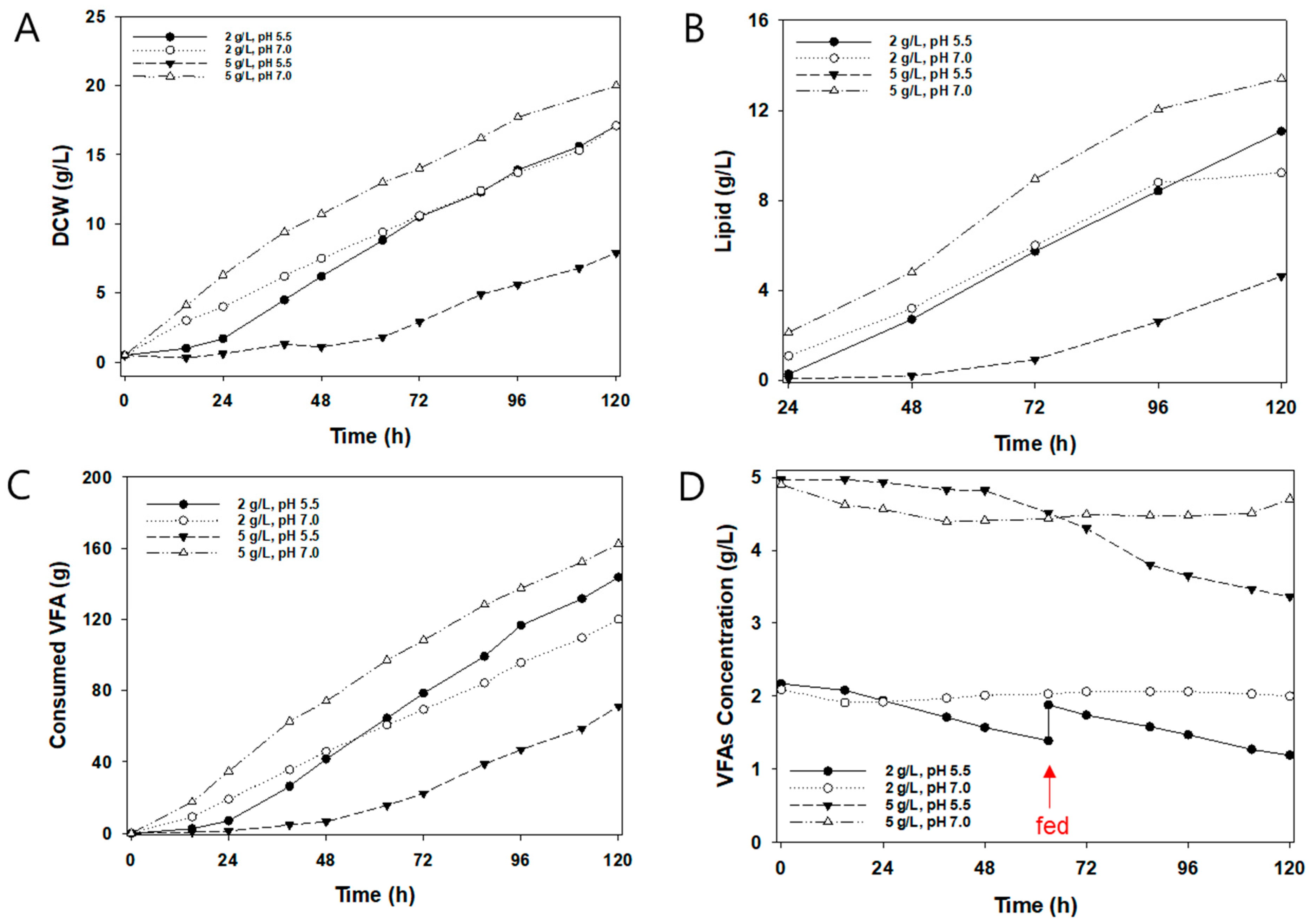
3.3. Effects of VFA Concentration and Initial pH on 5 L Automatic Fed-Batch
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Withanage, S.V.; Dias, G.M.; Habib, K. Review of household food waste quantification methods: Focus on composition analysis. J. Clean. Prod. 2021, 279, 123722. [Google Scholar] [CrossRef]
- Kiran, E.U.; Trzcinski, A.P.; Ng, W.J.; Liu, Y. Bioconversion of food waste to energy: A review. Fuel 2014, 134, 389–399. [Google Scholar] [CrossRef]
- Gao, A.; Tian, Z.; Wang, Z.; Wennersten, R.; Sun, Q. Comparison between the technologies for food waste treatment. Energy Procedia 2017, 105, 3915–3921. [Google Scholar] [CrossRef]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous yeasts as cell factories for the sustainable production of microbial lipids by the valorization of agri-food wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Chang, H.N.; Kim, N.-J.; Kang, J.; Jeong, C.M. Biomass-derived volatile fatty acid platform for fuels and chemicals. Biotechnol. Bioprocess Eng. 2010, 15, 1–10. [Google Scholar] [CrossRef]
- Park, G.W.; Seo, C.; Jung, K.; Chang, H.N.; Kim, W.; Kim, Y.-C. A comprehensive study on volatile fatty acids production from rice straw coupled with microbial community analysis. Bioprocess Biosyst. Eng. 2015, 38, 1157–1166. [Google Scholar] [CrossRef]
- Grubišić, M.; Mihajlovski, K.; Gruičić, A.M.; Beluhan, S.; Šantek, B.; Ivančić Šantek, M. Strategies for Improvement of Lipid Production by Yeast Trichosporon oleaginosus from Lignocellulosic Biomass. J. Fungi 2021, 7, 934. [Google Scholar] [CrossRef]
- Patel, A.; Sarkar, O.; Rova, U.; Christakopoulos, P.; Matsakas, L. Valorization of volatile fatty acids derived from low-cost organic waste for lipogenesis in oleaginous microorganisms-A review. Bioresour. Technol. 2021, 321, 124457. [Google Scholar] [CrossRef]
- Park, G.W.; Kim, N.J.; Chang, H.N. Microbial Lipid Production from Volatile Fatty Acids by Oleaginous Yeast. Emerg. Areas Bioeng. 2018, 1, 203–213. [Google Scholar]
- Park, G.W.; Son, S.; Moon, M.; Sin, S.; Min, K.; Lee, J.-S.; Chang, H.N. Volatile Fatty Acids from Lipid-Extracted Yeast Provide Additional Feedstock for Microbial Lipid Production. Catalysts 2021, 11, 1009. [Google Scholar] [CrossRef]
- Robles-Iglesias, R.; Naveira-Pazos, C.; Fernández-Blanco, C.; Veiga, M.C.; Kennes, C. Factors affecting the optimisation and scale-up of lipid accumulation in oleaginous yeasts for sustainable biofuels production. Renew. Sustain. Energy Rev. 2023, 171, 113043. [Google Scholar] [CrossRef]
- Scaldaferri, C.A.; Pasa, V.M.D. Hydrogen-free process to convert lipids into bio-jet fuel and green diesel over niobium phosphate catalyst in one-step. Chem. Eng. J. 2019, 370, 98–109. [Google Scholar] [CrossRef]
- Fei, Q.; Chang, H.N.; Shang, L.; Choi, J.-D.-R. Exploring low-cost carbon sources for microbial lipids production by fed-batch cultivation of Cryptococcus albidus. Biotechnol. Bioprocess Eng. 2011, 16, 482–487. [Google Scholar] [CrossRef]
- Llamas, M.; Magdalena, J.A.; González-Fernández, C.; Tomás-Pejó, E. Volatile fatty acids as novel building blocks for oil-based chemistry via oleaginous yeast fermentation. Biotechnol. Bioeng. 2020, 117, 238–250. [Google Scholar] [CrossRef]
- van der Klis, F.; Le Nôtre, J.; Blaauw, R.; van Haveren, J.; van Es, D.S. Renewable linear alpha olefins by selective ethenolysis of decarboxylated unsaturated fatty acids. Eur. J. Lipid Sci. Technol. 2012, 114, 911–918. [Google Scholar] [CrossRef]
- Beitel, S.M.; Coelho, L.F.; Contiero, J. Efficient conversion of agroindustrial waste into D(−) lactic acid by Lactobacillus delbrueckii using fed-batch fermentation. BioMed Res. Int. 2020, 2020, 4194052. [Google Scholar] [CrossRef] [Green Version]
- Falk, H.M.; Reichling, P.; Andersen, C.; Benz, R. Online monitoring of concentration and dynamics of volatile fatty acids in anaerobic digestion processes with mid-infrared spectroscopy. Bioprocess Biosyst. Eng. 2015, 38, 237–249. [Google Scholar] [CrossRef]
- Li, X.-Y.; Feng, Y.; Duan, J.-L.; Feng, L.-J.; Wang, Q.; Ma, J.-Y.; Liu, W.-Z.; Yuan, X.-Z. Model-based mid-infrared spectroscopy for on-line monitoring of volatile fatty acids in the anaerobic digester. Environ. Res. 2022, 206, 112607. [Google Scholar] [CrossRef]
- Kim, B.S.; Lee, S.C.; Lee, S.Y.; Chang, Y.K.; Chang, H.N. High cell density fed-batch cultivation of Escherichia coli using exponential feeding combined with pH-stat. Bioprocess Biosyst. Eng. 2004, 26, 147–150. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Drogui, P.; Tyagi, R.D. The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour. Technol. 2018, 259, 237–243. [Google Scholar] [CrossRef]
- Fei, Q.; O’Brien, M.; Nelson, R.; Chen, X.; Lowell, A.; Dowe, N. Enhanced lipid production by Rhodosporidium toruloides using different fed-batch feeding strategies with lignocellulosic hydrolysate as the sole carbon source. Biotechnol. Biofuels 2016, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anschau, A.; Xavier, M.C.; Hernalsteens, S.; Franco, T.T. Effect of feeding strategies on lipid production by Lipomyces starkeyi. Bioresour. Technol. 2014, 157, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Gorte, O.; Kugel, M.; Ochsenreither, K. Optimization of carbon source efficiency for lipid production with the oleaginous yeast Saitozyma podzolica DSM 27192 applying automated continuous feeding. Biotechnol. Biofuels 2020, 13, 181. [Google Scholar] [CrossRef] [PubMed]
- Park, G.W.; Shin, S.; Kim, S.J.; Lee, J.-S.; Moon, M.; Min, K. Rice straw-derived lipid production by HMF/furfural-tolerant oleaginous yeast generated by adaptive laboratory evolution. Bioresour. Technol. 2023, 367, 128220. [Google Scholar] [CrossRef]
- Ryu, B.-G.; Kim, W.; Heo, S.-W.; Kim, D.; Choi, G.-G.; Yang, J.-W. Advanced treatment of residual nitrogen from biologically treated coke effluent by a microalga-mediated process using volatile fatty acids (VFAs) under stepwise mixotrophic conditions. Bioresour. Technol. 2015, 191, 488–495. [Google Scholar] [CrossRef]
- Park, G.W.; Chang, H.N.; Jung, K.; Seo, C.; Kim, Y.-C.; Choi, J.H.; Woo, H.C.; Hwang, I.-J. Production of microbial lipid by Cryptococcus curvatus on rice straw hydrolysates. Process Biochem. 2017, 56, 147–153. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.-K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Sci. Rep. 2019, 9, 19383. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Yoon, J.H.; Lee, S.H.; Park, T.H. Hydrogen production from formic acid in pH-stat fed-batch operation for direct supply to fuel cell. Bioresour. Technol. 2010, 101, S53–S58. [Google Scholar] [CrossRef]
- Chi, Z.; Zheng, Y.; Ma, J.; Chen, S. Oleaginous yeast Cryptococcus curvatus culture with dark fermentation hydrogen production effluent as feedstock for microbial lipid production. Int. J. Hydrogen Energy 2011, 36, 9542–9550. [Google Scholar] [CrossRef]
- Gong, Z.; Shen, H.; Zhou, W.; Wang, Y.; Yang, X.; Zhao, Z.K. Efficient conversion of acetate into lipids by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2015, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhou, W.; He, Q.; Zhao, M.; Gong, Z. Microbial Lipid Production from High Concentration of Volatile Fatty Acids via Trichosporon cutaneum for Biodiesel Preparation. Appl. Biochem. Biotechnol. 2022, 194, 2968–2979. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Hakkaart, X.; Liu, Y.; Hulst, M.; El Masoudi, A.; Peuscher, E.; Pronk, J.; van Gulik, W.; Daran-Lapujade, P. Physiological responses of Saccharomyces cerevisiae to industrially relevant conditions: Slow growth, low pH, and high CO2 levels. Biotechnol. Bioeng. 2020, 117, 721–735. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yuan, M.; Liu, J.-N.; Huang, X.-F. Bioconversion of mixed volatile fatty acids into microbial lipids by Cryptococcus curvatus ATCC 20509. Bioresour. Technol. 2017, 241, 645–651. [Google Scholar] [CrossRef]
- Xu, X.; Kim, J.Y.; Cho, H.U.; Park, H.R.; Park, J.M. Bioconversion of volatile fatty acids from macroalgae fermentation into microbial lipids by oleaginous yeast. Chem. Eng. J. 2015, 264, 735–743. [Google Scholar] [CrossRef]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef]
- Annamalai, N.; Sivakumar, N.; Fernandez-Castane, A.; Oleskowicz-Popiel, P. Production of microbial lipids utilizing volatile fatty acids derived from wastepaper: A biorefinery approach for biodiesel production. Fuel 2020, 276, 118087. [Google Scholar] [CrossRef]
- Zheng, Y.; Chi, Z.; Ahring, B.K.; Chen, S. Oleaginous yeast Cryptococcus curvatus for biofuel production: Ammonia’s effect. Biomass Bioenergy 2012, 37, 114–121. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Bao, W.; Cheng, S.; Zheng, L. Enhanced lipid production by Yarrowia lipolytica cultured with synthetic and waste-derived high-content volatile fatty acids under alkaline conditions. Biotechnol. Biofuels 2020, 13, 3. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, Z. Biodiesel production by direct methanolysis of oleaginous microbial biomass. J. Chem. Technol. Biotechnol. 2007, 82, 775–780. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Manual Fed-Batch | Automatic Fed-Batch |
|---|---|---|
| DCW (g/L) | 13.40 | 19.95 |
| Biomass yield (g/g) | 0.35 | 0.37 |
| Lipid content (%) | 59 | 67 |
| Lipid (g/L) * | 7.91 | 13.37 |
| VFA consumption rate (g/L/h) | 0.32 | 0.45 |
| Category | Cultivation Condition (VFA Concentration, pH) | |||
|---|---|---|---|---|
| 2 g/L, pH 5.5 | 2 g/L, pH 7.0 | 5 g/L, pH 5.5 | 5 g/L, pH 7.0 | |
| Biomass yield (g/g) | 0.35 | 0.43 | 0.31 | 0.37 |
| Lipid yield (g/g) | 0.23 | 0.23 | 0.18 | 0.25 |
| VFA consumption rate (g/L/h) | 0.41 | 0.33 | 0.21 | 0.45 |
| Biomass productivity (g/L/h) | 0.14 | 0.14 | 0.04 | 0.17 |
| Lipid productivity (g/L/h) | 0.09 | 0.08 | 0.07 | 0.11 |
| Category | Cultivation Condition (VFA Concentration, pH) | ||||
|---|---|---|---|---|---|
| 2 g/L, pH 5.5 | 2 g/L, pH 7.0 | 5 g/L, pH 5.5 | 5 g/L, pH 7.0 | ||
| Fatty acid composition a (%) | C16:0 | 11.08 | 11.56 | 15.34 | 14.96 |
| C17:0 | 6.32 | 8.68 | 6.65 | 7.09 | |
| C17:1 | 1.23 | 2.19 | 1.62 | 1.50 | |
| C18:0 | 14.18 | 11.71 | 11.21 | 14.80 | |
| C18:1 | 53.64 | 49.88 | 50.17 | 47.67 | |
| C18:2 | 7.81 | 9.72 | 9.39 | 8.40 | |
| C18:3 | 2.62 | 3.29 | 2.36 | 2.84 | |
| Consumed VFA (g) | AA | 88.37 | 72.58 | 44.69 | 98.63 |
| PA | 14.47 | 10.87 | 5.50 | 12.63 | |
| BA | 45.24 | 36.88 | 25.79 | 51.77 | |
| Consumed VFA ratio | 59.7:9.8:30.6 | 60.3:9.0:30.6 | 58.8:7.2:34 | 60.5:7.7:31.8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.; Go, J.H.; Moon, M.; Park, G.W. Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids. Energies 2023, 16, 1996. https://doi.org/10.3390/en16041996
Shin S, Go JH, Moon M, Park GW. Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids. Energies. 2023; 16(4):1996. https://doi.org/10.3390/en16041996
Chicago/Turabian StyleShin, Subin, Jae Hee Go, Myounghoon Moon, and Gwon Woo Park. 2023. "Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids" Energies 16, no. 4: 1996. https://doi.org/10.3390/en16041996
APA StyleShin, S., Go, J. H., Moon, M., & Park, G. W. (2023). Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids. Energies, 16(4), 1996. https://doi.org/10.3390/en16041996