Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism

Abstract
:1. Introduction
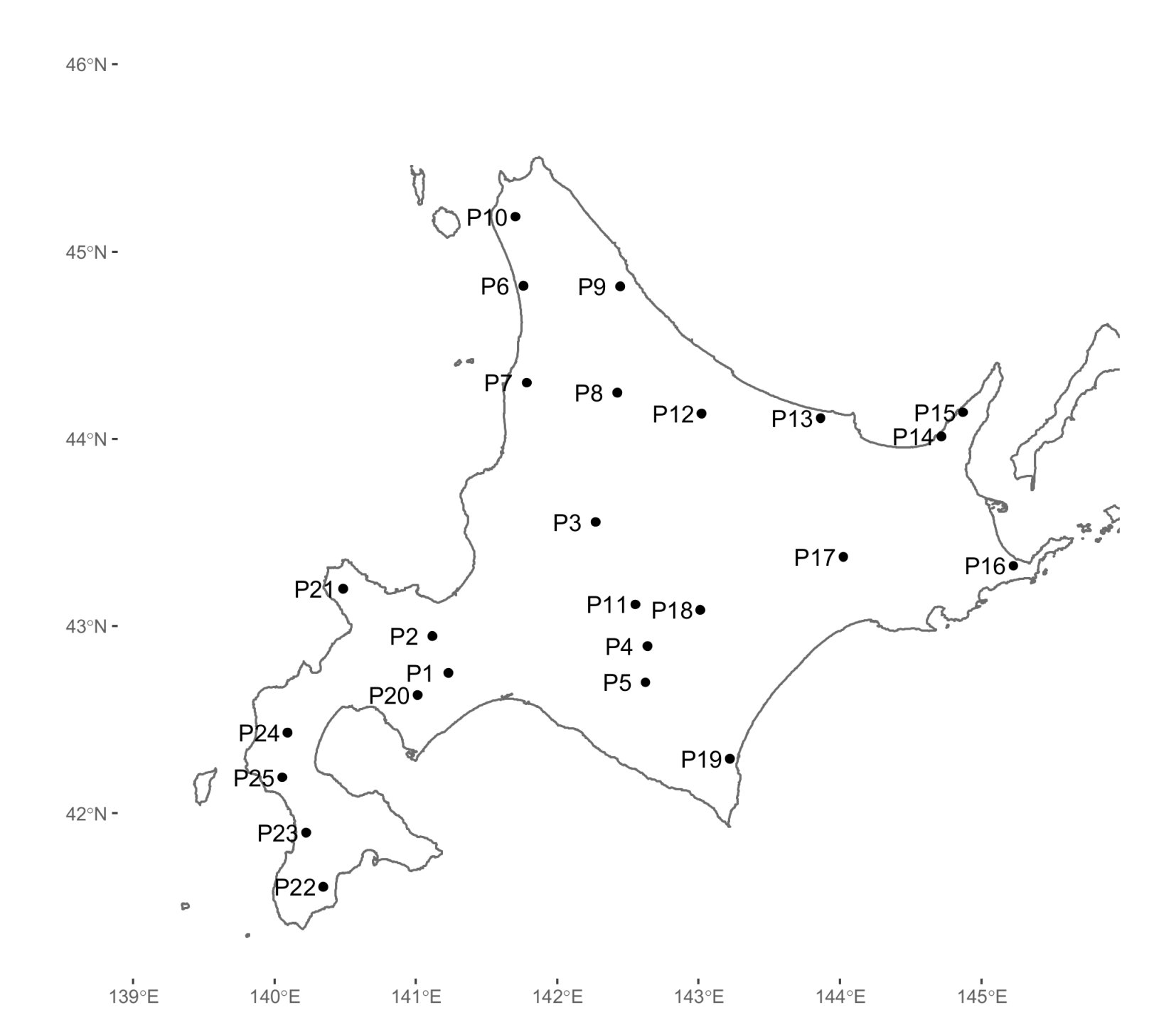
2. Materials and Methods
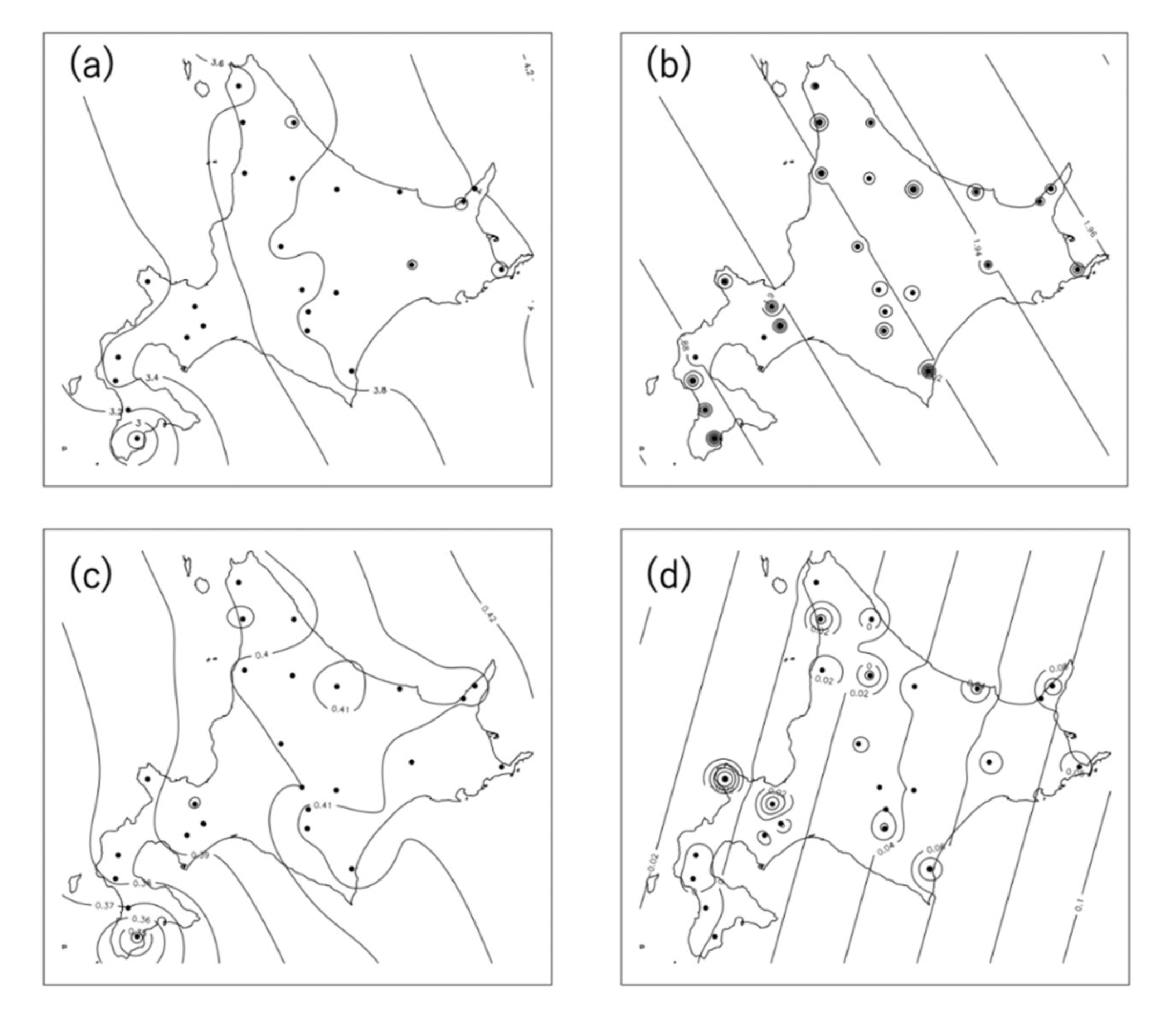
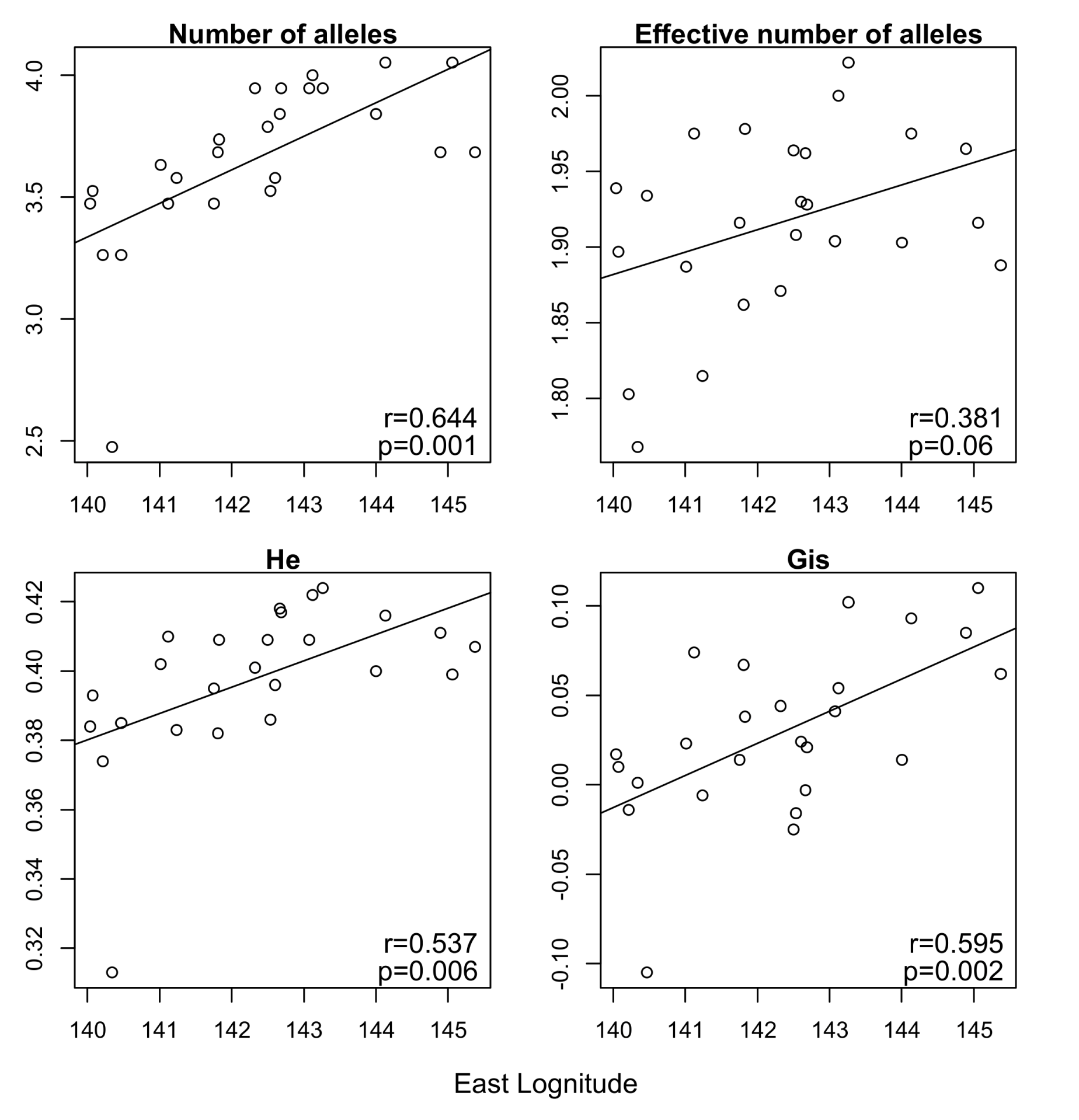
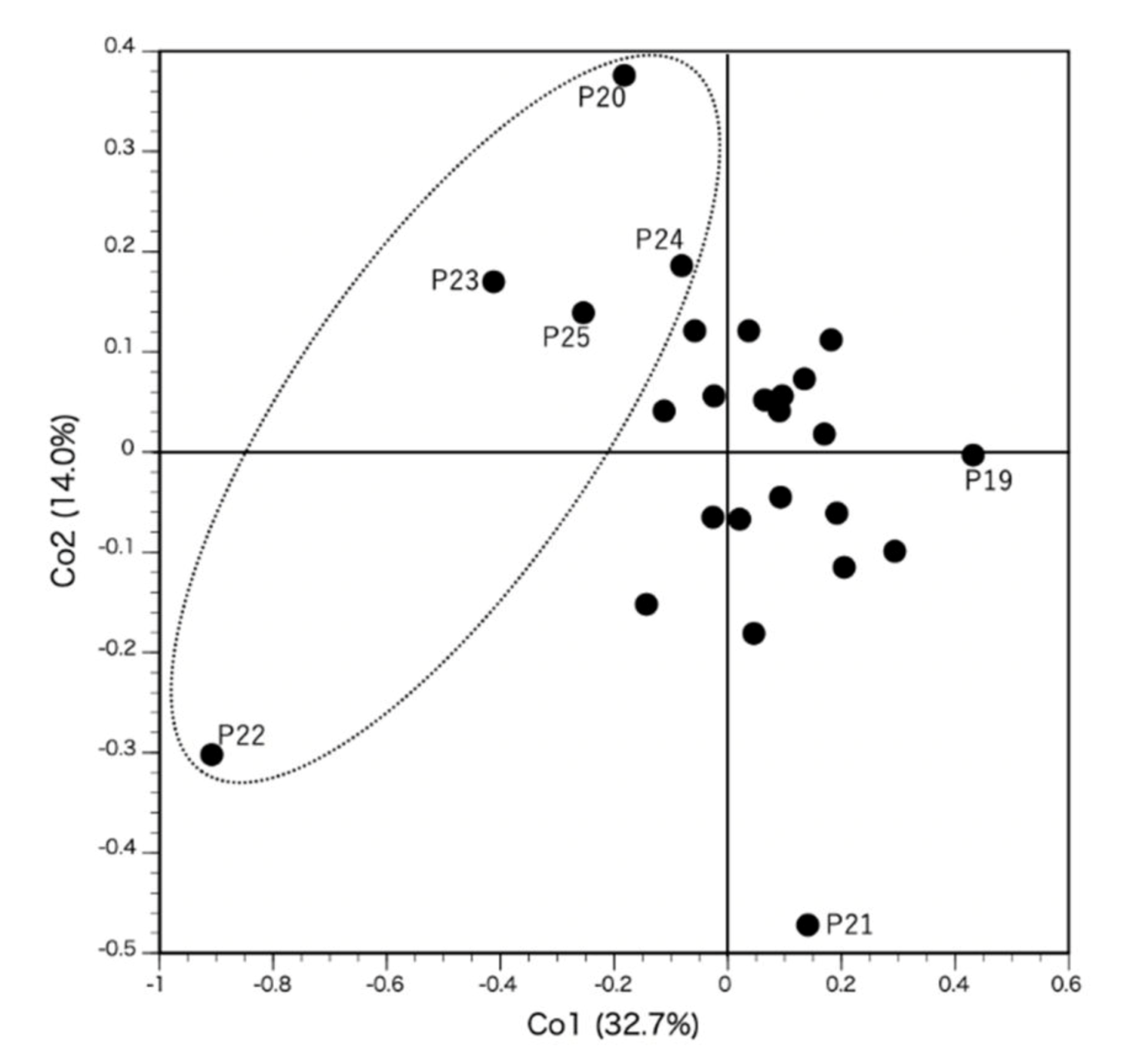
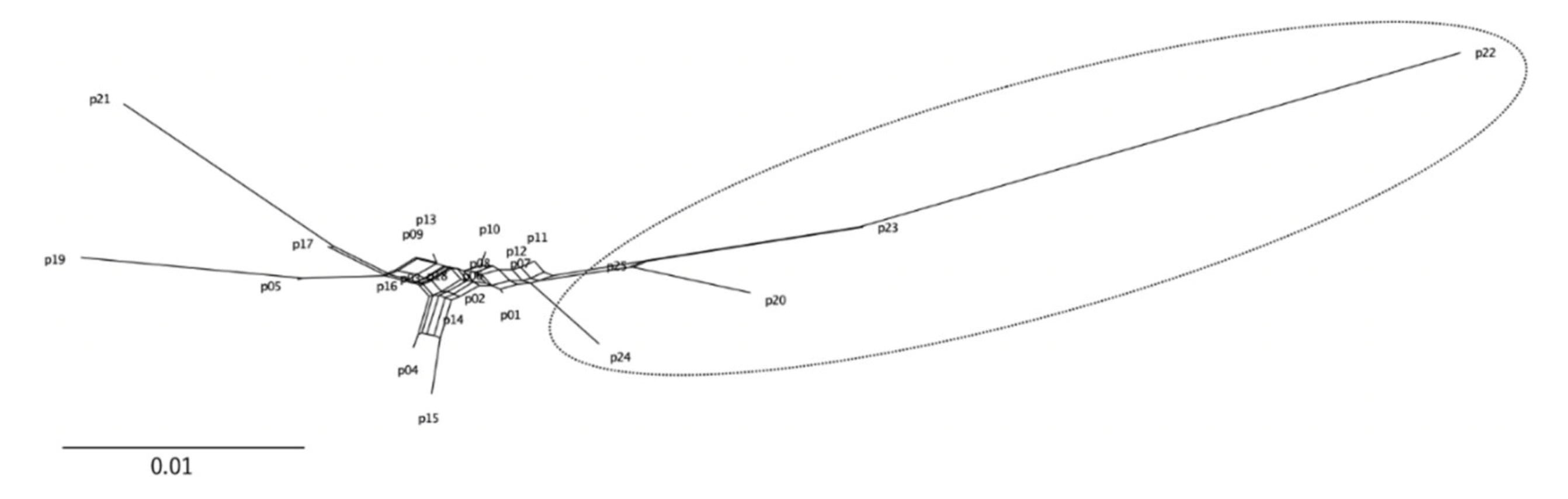
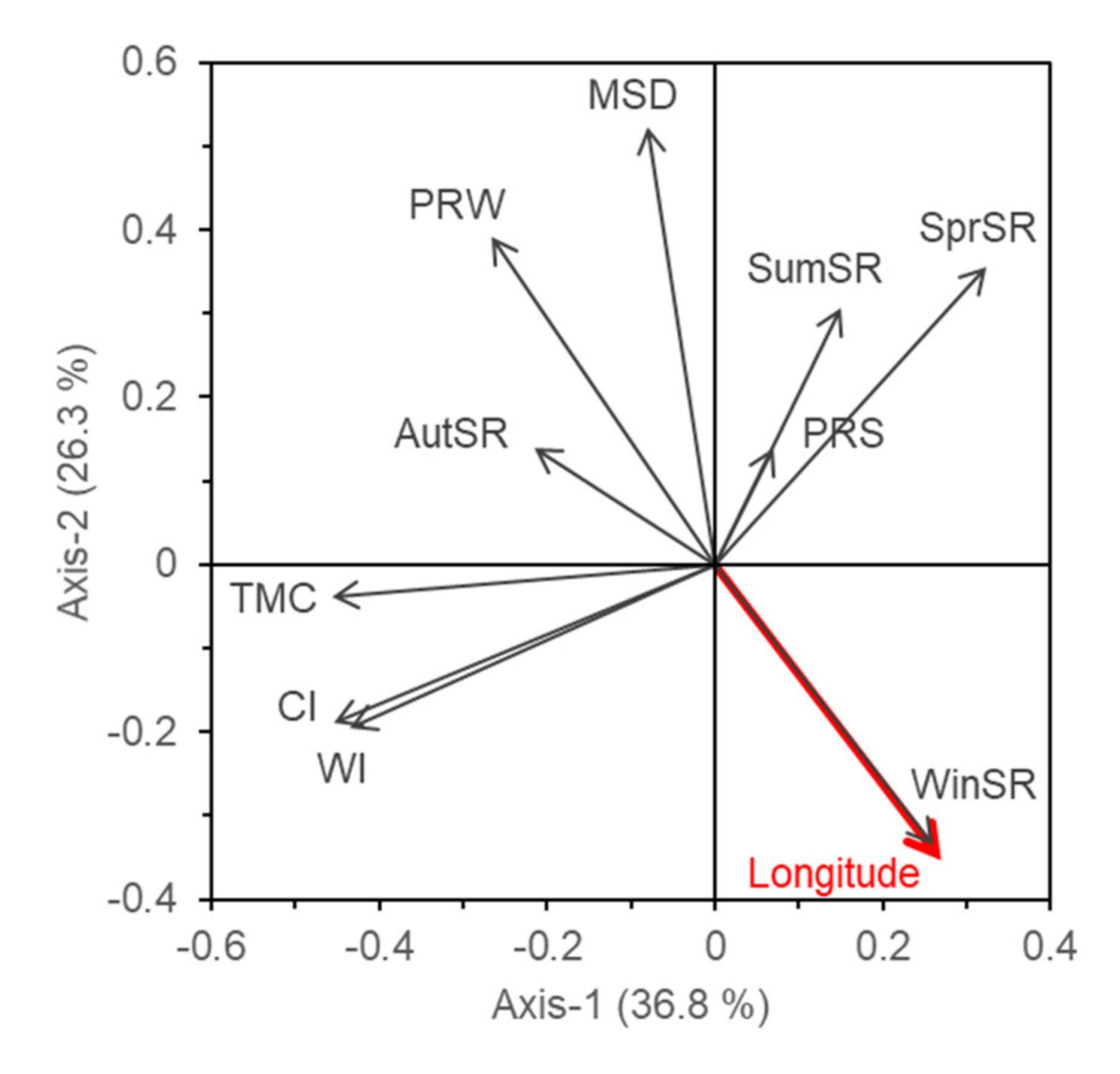
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petit, R.J.; El-Mousadik, A.; Pons, O. Identifying populations for conservation on the basis of genetic markers. Conserv. Biol. 1998, 12, 844–855. [Google Scholar] [CrossRef]
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Sagarin, R.D.; Gaines, S.D. The ‘abundant centre’ distribution: To what extent is it a biogeographical rule? Ecol. Lett. 2002, 5, 137–147. [Google Scholar] [CrossRef]
- Kitamura, K.; Matsui, T.; Kobayashi, M.; Saitou, H.; Namikawa, K.; Tsuda, Y. Decline in gene diversity and strong genetic drift in the northward-expanding marginal populations of Fagus crenata. Tree Genet. Genomes. 2015, 11, 36. [Google Scholar] [CrossRef]
- Eckert, C.G.; Samis, K.E.; Lougheed, S.C. Genetic variation across species’ geographical ranges: The central-marginal hypothesis and beyond. Mol. Ecol. 2008, 17, 1170–1188. [Google Scholar] [CrossRef]
- Arnaud-Haond, S.; Teixeira, S.; Massa, S.I.; Billot, C.; Saenger, P.; Coupland, G.; Duarte, C.M.; Serrao, E.A. Genetic structure at range edge: Low diversity and high inbreeding in Southeast Asian mangrove (Avicennia marina) populations. Mol. Ecol. 2006, 15, 3515–3525. [Google Scholar] [CrossRef]
- Tóth, E.G.; Vendramin, G.G.; Bagnoli, F.; Cseke, K.; Höhn, M. High genetic diversity and distinct origin of recently fragmented Scots pine (Pinus sylvestris L.) populations along the Carpathians and the Pannonian Basin. Tree Genet. Genomes 2017, 13, 47. [Google Scholar]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Petit, R.J.; Bialozyt, R.; Garnier-Géré, P.; Hampe, A. Ecology and genetics of tree invasions: From recent introductions to Quaternary migrations. For. Ecol. Manag. 2004, 197, 117–137. [Google Scholar] [CrossRef]
- Tatewaki, M. Forest ecology of the islands of the north Pacific ocean. J. Fac. Agric. Hokkaido Univ. 1958, 50, 341–486. [Google Scholar]
- Kitamura, S.; Murata, G. Colored Illustrations of Woody Plants of Japan Volume II; Hoikusya Co., Ltd.: Osaka, Japan, 1979; p. 545. [Google Scholar]
- Matsuda, S. Distribution status of Abies sachalinensis and Picea jezoensis in the Hokkaido national forest. Hokkaido Ringyo Kaiho 1936, 34, 351–361. [Google Scholar]
- Matsui, T.; Kitamura, K.; Saito, H.; Namikawa, K.; Terazawa, K.; Haruki, M.; Itaya, A.; Honma, Y.; Miyoshi, Y.; Uchida, K.; et al. Habitat and vegetation of an outlying Fagus crenata population in its northern range limit at Rebunge Pass, Hokkaido. J. Phyto. Taxon. 2012, 59, 113–123. [Google Scholar]
- Kurahashi, A.; Hamaya, T. Variation of morphological characters and growth response of Saghalin fir (Abies sachalinensis) in different altitude. Bull. Tokyo Univ. For. 1981, 71, 101–151. [Google Scholar]
- Hatakeyama, S. Genetical and breeding studies on the regional differences of interprovenance variation in Abies sachalinensis Mast. Bull. Hokkaido For. Exp. Stn. 1981, 19, 1–91. [Google Scholar]
- Eiga, S. Ecological study on the freezing resisntace of Saghalin fir (Abies sachalinensis Mast.) in Hokkaido. Bull. For. Tree Breed. Inst. 1984, 2, 61–107. [Google Scholar]
- Ishizuka, W.; Goto, S. Modeling intraspecific adaptation of Abies sachalinensis to local altitude and responses to global warming, based on a 36-year reciprocal transplant experiment. Evol. Appl. 2012, 5, 229–244. [Google Scholar] [CrossRef]
- Ishizuka, W.; Ono, K.; Hara, T.; Goto, S. Use of intraspecific variation in thermal responses for estimating an elevational cline in the timing of cold hardening in a sub-boreal conifer. Plant Biol. 2015, 17, 177–185. [Google Scholar] [CrossRef]
- Goto, S.; Kajiya-Kanegae, H.; Ishizuka, W.; Kitamura, K.; Ueno, S.; Hisamoto, Y.; Kudoh, H.; Yasugi, M.; Nagano, A.J.; Iwata, H. Genetic mapping of local adaptation along the altitudinal gradient in Abies sachalinensis. Tree Genet. Genomes 2017, 13, 104. [Google Scholar] [CrossRef]
- Nagasaka, K.; Wang, Z.M.; Tanaka, K. Genetic variation among natural Abies sachalinensis population in relation to environmental gradients in Hokkaido, Japan. For. Genet. 1997, 4, 43–50. [Google Scholar]
- Suyama, Y.; Yoshimaru, H.; Tsumura, Y. Molecular phylogenetic position of Japanese Abies (Pinaceae) based on chloroplast DNA sequences. Mol. Phylogenetics Evol. 2000, 16, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Postolache, D.; Leonarduzzi, C.; Piotti, A.; Spanu, I.; Roig, A.; Fady, B.; Roschanski, A.; Liepelt, S.; Vendramin, G.G. Transcriptome versus genomic microsatellite markers: Highly informative multiplexes for genotyping Abies alba Mill. and congeneric species. Plant Mol. Biol. Rep. 2014, 32, 750–760. [Google Scholar] [CrossRef]
- Ueno, S.; Nakamura, Y.; Kobayashi, M.; Terashima, S.; Ishizuka, W.; Uchiyama, K.; Tsumura, Y.; Yano, K.; Goto, S. TodoFirGene: Developing transcriptome resources for genetic analysis of Abies sachalinensis. Plant Cell Physiol. 2018, 59, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Ueno, S.; Moriguchi, Y.; Uchiyama, K.; Ujino-Ihara, T.; Futamura, N.; Sakurai, T.; Shinohara, K.; Tsumura, Y. A second generation framework for the analysis of microsatellites in expressed sequence tags and the development of EST-SSR markers for a conifer, Cryptomeria japonica. BMC Genom. 2012, 13, 136. [Google Scholar] [CrossRef] [Green Version]
- Meirmans, P.G.; Van Tienderen, P.H. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes. 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Mase, S. Geostatistics and Kriging Method; Ohmsha: Tokyo, Japan, 2010. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 18 July 2018).
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- The Japan Meterological Agency. Mesh Climate Data 2000, CD-ROM; The Japan Meterological Agency: Tokyo, Japan, 2002. [Google Scholar]
- Tanaka, N.; Nakao, K.; Tsuyama, I.; Matsui, T. Assessing impact of climate warming on potential habitats of ten conifer species in Japan. AIRIES 2009, 14, 153–164. [Google Scholar]
- Conord, C.; Gurevitch, J.; Fady, B. Large-scale longitudinal gradients of genetic diversity: A meta-analysis across six phyla in the Mediterranean basin. Ecol. Evol. 2012, 2, 2600–2614. [Google Scholar] [CrossRef]
- Yokota, S. Scleroderris canker of Todo-fir in Hokkaido, Northern Japan. Eur. J. For. Path. 1975, 5, 13–21. [Google Scholar] [CrossRef]
- Okada, S.; Mukaide, H.; Sakai, A. Genetic variation in Saghalien fir from different areas of Hokkaido. Silvae Genet. 1973, 22, 1–2. [Google Scholar]
- Jump, A.S.; Hunt, J.M.; Martínez-Izquierdo, J.A.; Peñuelas, J. Natural selection and climate change: Temperature-linked spatial and temporal trends in gene frequency in Fagus sylvatica. Mol. Ecol. 2006, 15, 3469–3480. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.K.; Chase, M.W.; De Bruijn, A.; Fay, M.F.; Woodward, F.I. Temperature-based population segregation in birch. Ecol. Lett. 2003, 6, 87–89. [Google Scholar] [CrossRef]
- Grivet, D.; Sebastiani, F.; Alía, R.; Bataillon, T.; Torre, S.; Zabal-Aguirre, M.; Vendramin, G.G.; González-Martínez, S.C. Molecular footprints of local adaptation in two Mediterranean conifers. Mol. Biol. Evol. 2010, 28, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, D.; Smithson, A.; Krauss, S.L. Signatures of diversifying selection at EST-SSR loci and association with climate in natural Eucalyptus populations. Mol. Ecol. 2013, 22, 5112–5129. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Peñuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Mitton, J.B.; Grant, M.C. Observations on the ecology and evolution of quaking aspen, Populus tremuloides, in the Colorado front range. Am. J. Bot. 1980, 67, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Kitamura, K.; Lian, C.; Goto, S. The effects of multilocus heterozygosity on the longevity of seedlings established on fallen logs in Picea jezoensis and Abies sachalinensis. Open J. For. 2015, 5, 422–430. [Google Scholar]
- Leimu, R.; Mutikainen, P.I.A.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Mitton, J.B.; Grant, M.C. Associations among protein heterozygosity, growth rate, and developmental homeostasis. Annu. Rev. Ecol. Syst. 1984, 15, 479–499. [Google Scholar] [CrossRef]
- Smith, J.M.; Haigh, J. The hitch-hiking effect of a favourable gene. Genet. Res. 1974, 23, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kado, T.; Yoshimaru, H.; Tsumura, Y.; Tachida, H. DNA variation in a conifer, Cryptomeria japonica (Cupressaceae sensu lato). Genetics 2003, 164, 1547–1559. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945. [Google Scholar] [PubMed]
- Leonarduzzi, C.; Piotti, A.; Spanu, I.; Vendramin, G.G. Effective gene flow in a historically fragmented area at the southern edge of silver fir (Abies alba Mill.) distribution. Tree Genet. Genomes 2016, 12, 95. [Google Scholar] [CrossRef]
- Iwaizumi, M.G.; Aizawa, M.; Watanabe, A.; Goto, S. Highly polymorphic nuclear microsatellite markers reveal detailed patterns of genetic variation in natural populations of Yezo spruce in Hokkaido. J. For. Res. 2015, 20, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, M.; Yoshimaru, H.; Takahashi, M.; Kawahara, T.; Sugita, H.; Saito, H.; Sabirov, R.N. Genetic structure of Sakhalin spruce (Picea glehnii) in northern Japan and adjacent regions revealed by nuclear microsatellites and mitochondrial gene sequences. J. Plant Res. 2015, 128, 91–102. [Google Scholar] [CrossRef]
- Hayashi, E.; Ubukata, M.; Iizuka, K.; Itahana, N. Genetic differentiation of organelle DNA polymorphisms in Sakhalin fir from Hokkaido. For. Genet. 2000, 7, 31–38. [Google Scholar]
- Takiya, M.; Hagiwara, N. Vegetational history of Mt. Yokotsudake, southwestern Hokkaido, since the Last Glacial. Quat. Res. 1997, 36, 217–234. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population ID | Longitude East (°) | Latitude North (°) | Alt 1 (m) | N 2 | Nttl_a 3 | Na 4 | Nef 5 | HO | HE | GIS |
|---|---|---|---|---|---|---|---|---|---|---|
| P1 | 141.2365 | 42.7691 | 800 | 44 | 68 | 3.579 | 1.815 | 0.385 | 0.383 | −0.006 |
| P2 | 141.1201 | 42.9663 | 580 | 48 | 66 | 3.474 | 1.975 | 0.379 | 0.410 | 0.074 |
| P3 | 142.3221 | 43.5685 | 300 | 50 | 75 | 3.947 | 1.871 | 0.383 | 0.401 | 0.044 |
| P4 | 142.6851 | 42.9003 | 930 | 47 | 75 | 3.947 | 1.928 | 0.408 | 0.417 | 0.021 |
| P5 | 142.6656 | 42.7072 | 700 | 48 | 73 | 3.842 | 1.962 | 0.419 | 0.418 | −0.003 |
| P6 | 141.8080 | 44.8361 | 5 | 46 | 70 | 3.684 | 1.862 | 0.356 | 0.382 | 0.067 |
| P7 | 141.8255 | 44.3187 | 90 | 48 | 71 | 3.737 | 1.978 | 0.394 | 0.409 | 0.038 |
| P8 | 142.4986 | 44.2581 | 285 | 48 | 72 | 3.789 | 1.964 | 0.419 | 0.409 | −0.025 |
| P9 | 142.5356 | 44.8255 | 260 | 48 | 67 | 3.526 | 1.908 | 0.392 | 0.386 | −0.016 |
| P10 | 141.7521 | 45.2064 | 10 | 48 | 66 | 3.474 | 1.916 | 0.389 | 0.395 | 0.014 |
| P11 | 142.6033 | 43.1250 | 360 | 43 | 68 | 3.579 | 1.930 | 0.387 | 0.396 | 0.024 |
| P12 | 143.1218 | 44.1365 | 250 | 48 | 76 | 4.000 | 2.000 | 0.399 | 0.422 | 0.054 |
| P13 | 144.0025 | 44.0930 | 200 | 48 | 73 | 3.842 | 1.903 | 0.395 | 0.400 | 0.014 |
| P14 | 144.8912 | 43.9676 | 73 | 48 | 70 | 3.684 | 1.965 | 0.376 | 0.411 | 0.085 |
| P15 | 145.0595 | 44.0907 | 100 | 47 | 77 | 4.053 | 1.916 | 0.355 | 0.399 | 0.110 |
| P16 | 145.3731 | 43.2588 | 20 | 48 | 70 | 3.684 | 1.888 | 0.382 | 0.407 | 0.062 |
| P17 | 144.1321 | 43.3482 | 950 | 48 | 77 | 4.053 | 1.975 | 0.377 | 0.416 | 0.093 |
| P18 | 143.0752 | 43.0874 | 750 | 48 | 75 | 3.947 | 1.904 | 0.392 | 0.409 | 0.041 |
| P19 | 143.2620 | 42.2881 | 73 | 48 | 75 | 3.947 | 2.022 | 0.380 | 0.424 | 0.102 |
| P20 | 141.0124 | 42.6518 | 450 | 48 | 69 | 3.632 | 1.887 | 0.393 | 0.402 | 0.023 |
| P21 | 140.4678 | 43.2183 | 200 | 48 | 62 | 3.263 | 1.934 | 0.425 | 0.385 | −0.105 |
| P22 | 140.3397 | 41.6239 | 90 | 48 | 47 | 2.474 | 1.768 | 0.312 | 0.313 | 0.001 |
| P23 | 140.2135 | 41.9132 | 470 | 48 | 62 | 3.263 | 1.803 | 0.379 | 0.374 | −0.014 |
| P24 | 140.0727 | 42.4468 | 620 | 48 | 67 | 3.526 | 1.897 | 0.389 | 0.393 | 0.010 |
| P25 | 140.0379 | 42.2073 | 100 | 48 | 66 | 3.474 | 1.939 | 0.378 | 0.384 | 0.017 |
| Overall | 1189 | 0.386 | 0.398 | 0.030 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitamura, K.; Uchiyama, K.; Ueno, S.; Ishizuka, W.; Tsuyama, I.; Goto, S. Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism. Forests 2020, 11, 233. https://doi.org/10.3390/f11020233
Kitamura K, Uchiyama K, Ueno S, Ishizuka W, Tsuyama I, Goto S. Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism. Forests. 2020; 11(2):233. https://doi.org/10.3390/f11020233
Chicago/Turabian StyleKitamura, Keiko, Kentaro Uchiyama, Saneyoshi Ueno, Wataru Ishizuka, Ikutaro Tsuyama, and Susumu Goto. 2020. "Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism" Forests 11, no. 2: 233. https://doi.org/10.3390/f11020233
APA StyleKitamura, K., Uchiyama, K., Ueno, S., Ishizuka, W., Tsuyama, I., & Goto, S. (2020). Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism. Forests, 11(2), 233. https://doi.org/10.3390/f11020233