


The Variations in Tracheid Length of Pseudotsuga menziesii (Mirb.) Franco Wood in Relation to Cambium Age, Site, and Growth



Abstract
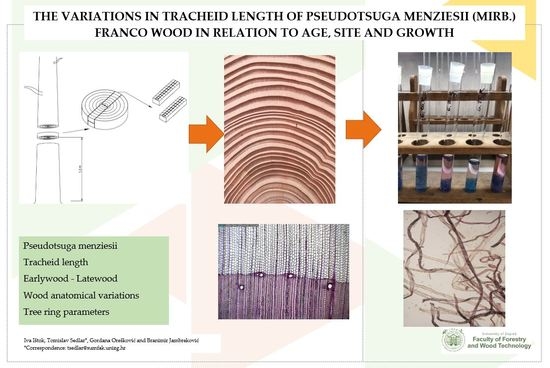
:
1. Introduction
2. Materials and Methods
2.1. Site and Study Materials
2.2. Sample Collection and Preparation
2.3. Tracheid Length Measurement
2.4. ARW Measurement
2.5. Statistical Analysis
3. Results
3.1. Effect of Annual Growth Rings, Tree, and Site on Tracheid Length and Changes in Earlywood and Latewood
3.2. Different Patterns of Correlations between Tree Ring Parameters and Tracheid Length
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECDiLibrary. Douglas-fir (Pseudotsuga menziesii). Available online: https://www.oecd-ilibrary.org/docserver/9789264095434-8en.pdf?expires=1682675767&id=id&accname=guest&checksum=76B694AF1FDBB1223FEE1FCF80150A95 (accessed on 23 February 2023).
- Da Ronch, F.; Caudullo, G.; de Rigo, D. Pseudotsuga menziesii in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Levanič, T.; Štraus, H. Effects of Climate on Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) Growth Southeast of the European Alps. Plants 2022, 11, 1571. [Google Scholar] [CrossRef] [PubMed]
- Iangum, C.E.; Yadama, V.; Iowell, E.C. Physical and Mechanical Properties of Young-Growth Douglas-Fir and Western Hemlock from Western Washington. For. Prod. J. 2009, 59, 37–47. [Google Scholar]
- Viguier, J.; Bourreau, D.; Bocquet, J.-F.; Pot, G.; Bléron, L.; Lanvin, J.D. Modelling mechanical properties of spruce and Douglas fir timber by means of X-ray and grain angle measurements for strength grading purpose. Eur. J. Wood Wood Prod. 2017, 75, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Henin, J.-M.; Pollet, C.; Schmitt, U.; Blohm, J.-H.; Koch, G.; Melcher, E.; Welling, J.; Brüchert, F.; Kohnle, U.; Sauter, U.H. Technical properties of Douglas-fir wood. In Douglas-fir—An Option for Europe; Spiecker, H., Lindner, M., Schuler, J., Eds.; The European Forest Institute: Joensuu, Finland, 2019. [Google Scholar]
- Wiedenhoeft, A.; Eberhardt, T.L. Structure and function of wood. In Wood Handbook Wood as an Engineering Material; General Technical Report FPL-GTR-282; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 2021; 20p. [Google Scholar]
- Panshin, A.J.; de Zeeuw, C. Textbook of Wood Technology: Structure, Identification, Properties, and Uses of the Commercial Woods of the United States and Canada; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Beaulieu, J. Genetic variation in tracheid length and relationships with growth and wood traits in eastern white spruce (Picea glauca). Wood Fiber Sci. 2003, 35, 609–616. [Google Scholar]
- Kibblewhite, R.P.; Evans, R.; Riddell, M.J.C. Kraft handsheet, and wood tracheid and chemical property interrelationships for 50 individual radiata pine trees. Appita J. 2003, 56, 229–233. [Google Scholar]
- Bendtsen, B.A.; Senft, J. Mechanical and anatomical properties in individual growth rings of plantation-grown eastern cottonwood and loblolly pine. Wood Fiber Sci. 1986, 18, 23–38. [Google Scholar]
- Gogoi, B.R.; Sharma, M.; Sharma, C.L. Intra-Ring Tracheid Length Variations in Khasi pine (Pinus kesiya Royle ex Gordon) at Breast Heigh. Int. J. Life Sci. Res. 2017, 5, 147–156. [Google Scholar]
- Tsoumis, G. Science and Technology of Wood: Structure, Properties, Utilization; Chapman & Hall: New York, NY, USA, 1991; pp. 66–83. [Google Scholar]
- Zobel, B.J.; Sprague, J.R. Juvenile Wood in Forest Trees; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Bao, F.C.; Jiang, Z.H.; Jiang, X.M.; Lu, X.X.; Luo, X.Q.; Zhang, S.Y. Differences in wood properties between juvenile wood and mature wood in 10 species grown in China. Wood Sci. Technol. 2001, 35, 363–375. [Google Scholar] [CrossRef]
- Acuna, M.A.; Murphy, G. Geospatial and within tree variation of wood density and spiral grain in Douglas-fir. For. Prod. J. 2006, 56, 81–85. [Google Scholar]
- Foti, D.; Passialis, C.; Voulgaridis, E.; Skaltsoyiannes, A.; Tsaktsira, M. Wood density and tracheid length of Aleppo pine (Pinus halepensis Mill.) grafts in relation to cambium age and growth rate. J. For. Sci. 2018, 64, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Dinwoodie, J.M. Tracheid and fibre length in timber—A review of literature. For. Int. J. For. Res. 1961, 34, 125–144. [Google Scholar] [CrossRef]
- Bouslimi, B.; Koubaa, A.; Bergeron, Y. Intra-ring Variations and Interrelationships for Selected Wood Anatomical and Physical Properties of Thuja Occidentalis L. Forests 2019, 10, 339. [Google Scholar] [CrossRef] [Green Version]
- Mvolo, C.S.; Goudiaby, V.; Koubaa, A.; Stewart, J.D. Influence of Four Spacings between Trees and Four Samplings Heights on Selected Wood Quality Attributes of White Spruce (Picea glauca (Moench) Voss). Forests 2022, 13, 1807. [Google Scholar] [CrossRef]
- Taylor, F.W.; Moore, J.S. A comparison of earlywood and latewood tracheid lengths of loblolly pine. Wood Fiber 1981, 13, 159–165. [Google Scholar]
- Zimmer, B.; Grosser, D.; Mehlen, S. Studies on the tracheid lengths of Douglas-fir wood. Holz als Roh-und Werkstoff 1998, 56, 252. [Google Scholar] [CrossRef]
- DeSoto, L.; De la Cruz, M.; Fonti, P. Intra-annual patterns of tracheid size in the Mediterranean tree Juniperus thurifera as an indicator of seasonal water stress. Can. J. For. Res. 2011, 41, 1280–1294. [Google Scholar] [CrossRef] [Green Version]
- Mvolo, C.S.; Koubaa, A.; Defo, M.; Beaulieu, J.; Yemele, M.-C.; Cloutier, A. Prediction of tracheid length and diameter in white spruce (Picea glauca). IAWA J. 2015, 36, 186–207. [Google Scholar] [CrossRef]
- Koubaa, A.; Zhang, S.Y.; Isabel, N.; Beaulieu, J.; Bousquet, J. Phenotypic correlations between juvenile-mature wood density and growth in black spruce. Wood Fiber Sci. 2000, 32, 61–71. [Google Scholar]
- Kiaei, M.; Sadegh, A.N.; Moya, R. Site variation of tracheid features and static bending properties in Pinus eldarica wood. Cellul. Chem. Technol. 2013, 47, 49–59. [Google Scholar]
- Muneri, A.; Balodis, V. Variation in Wood Density and Tracheid Length in Pinus patula grown in Zimbabwe. S. Afr. For. J. 2010, 182, 41–50. [Google Scholar]
- Klepac, D. Prilog poznavanju rasta i prirasta zelene duglazije i američkog borovca. Sumar List 1962, 86, 10–31. [Google Scholar]
- Orlić, S.; Ocvirek, M. Istraživanje provenijencije zelene duglazije (Pseudotsuga menziesii (Mirb.) Franco. u Hrvatskoj. Sumar List 1996, 11–12, 455–462. [Google Scholar]
- Orlić, S.; Perić, S. Proučavanje uspijevanja provenijencija duglazije (Pseudotsuga menziesii (Mirb.) Franco) na Krndiji. Sumar List 2005, 5–6, 243–250. [Google Scholar]
- Perić, S.; Tijardović, M.; Jazbec, A. Rezultati istraživanja provenijencija zelene duglazije u ekološki različitim pdoručjima kontinentalne Hrvatske. Sumar List Spec. Issue 2011, 135, 190–201. [Google Scholar]
- Đodan, M.; Dubravac, T.; Perić, S. Which Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances Provide the Best Productivity in the Hilly Area of Croatia? SEEFOR 2019, 10, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Wagenführ, R.; Scheiber, C. Holzatlas; VEB Fchbuchverlag: Leipzig, Germany, 2006; pp. 221–223. [Google Scholar]
- Rathgeber, C.B.K.; Decoux, V.; Leban, J.-M. Linking intra-tree-ring wood density variations and tracheid anatomical characteristics in Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco). Ann. For. Sci. 2006, 63, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Franklin, G.L. Preparation of thin-wood sections of synthetic resins and wood-resin composites, and a new macerating method for wood. Nature 1945, 155, 51. [Google Scholar] [CrossRef]
- IAWA Committee. IAWA list of microscopic features for softwood identification. IAWA J. 2004, 25, 1–70. [Google Scholar] [CrossRef]
- Wilkes, J. Interpreting patterns of variation in tracheid morphology in Pinus species. Commonw. For. Rev. 1987, 66, 177–190. [Google Scholar]
- Mvolo, C.S.; Koubaa, A.; Beaulieu, J.; Cloutier, A.; Defo, M.; Yemele, M.C. Phenotypic Correlations among Growth and Selected Wood Properties in White Spruce (Picea glauca (Moench) Voss). Forests 2019, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- Sanio, K. Über die Grösse Holzellen bei der gemeinen Kiefer (Pinus sylvestris). Jahrb. Wiss. Bot. 1872, 8, 401–420. [Google Scholar]
- Blohm, J.H.; Evans, R.; Koch, G.; Schnitt, U. Identification and characterization of Douglas-fir (Pseudotsuga menziesii (mirb.) Franco) juvenile and adult wood grown in southern Germany. Drewno 2016, 59, 41–47. [Google Scholar] [CrossRef]
- Harris, J.M. Physical properties, resin content, and tracheid length of lodgepole pine grown in New Zealand. N. Z. JI For. Sci. 1971, 3, 91–109. [Google Scholar]
- Adamopoulos, S.; Wimmer, R.; Milios, E. Tracheid length–growth relationships of young Pinus bruta grown on reforestation sites. IAWA J. 2012, 33, 39–49. [Google Scholar] [CrossRef]
- Zheng, J.; Li, Y.; Morris, H.; Vandelook, F.; Jansen, S. Variation in Tracheid Dimensions of Conifer Xylem Reveals Evidence of Adaptation to Environmental Conditions. Front. Plant Sci. 2022, 13, 774241. [Google Scholar] [CrossRef]
- Gartner, B.L.; Robbins, J.M.; Newton, M. Effects of pruning on wood density and tracheid in young Douglas-fir. Wood Fiber Sci. 2005, 37, 304–313. [Google Scholar]
- Fujivara, S.; Yang, K.C. The relationship between cell length and ring width and circumferential growth rate in five Canadian species. IAWA J. 2000, 21, 335–345. [Google Scholar] [CrossRef]
- Fabisiak, E.; Fabisiak, B.; Krauss, A. Radial Variation in Tracheid Lengths in Dominant Trees of Selected Coniferous Species. BioResources 2020, 15, 7330–7341. [Google Scholar] [CrossRef]
- Lenz, P.; Cloutier, A.; Mackay, J.; Beaulieu, J. Genetic control of wood properties in Picea glauca–An analysis of trends with cambial age. Can. J. For. Res. 2010, 40, 703–715. [Google Scholar] [CrossRef]
- Brändström, J. Micro and ultrastructural aspects of Norway spruce tracheids: A review. IAWA J. 2001, 22, 333–353. [Google Scholar] [CrossRef] [Green Version]
- McKimmy, M.D. Factors related to variation of specific gravity in young-growth Douglas-fir. Oregon For. Prod. Res. Cent. Bull. 1959, 8, 52. [Google Scholar]
- de Kort, I. Tracheid length in vital and non vital Douglas fir (Pseudotsuga menziesii) in the Netherlands. IAWA J 1990, 11, 203–209. [Google Scholar] [CrossRef]
- Zobel, B.J.; van Buijetenen, J.P. Wood Variation: Its Causes and Control; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Czajka, M.; Fabisiak, E. Variation in the tracheids length and macrostructural parameters of Douglas fir wood with developed reaction tissue. Ann. WULS-SGGW For. Wood Technol. 2015, 92, 74–79. [Google Scholar]
- Duchesne, I.; Zhang, S.Y. Variation in tree growth, wood density, and pulp fiber properties of 35 white spruce (Picea glauca (Moench) Voss) families grown in Quebec. Wood Fiber Sci. 2004, 36, 467–475. [Google Scholar]
- Larson, P.R.; Krestchmann, D.E.; Clark, A., III; Isebrands, J.G. Formation and Properties of Juvenile Wood in Southern Pines: A Synopsis; General Technical Report, FPL-GTR-129; United States, Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 2001; p. 42. [Google Scholar]
- Mergen, F.; Burley, J.; Yeatman, C.W. Variation in growth characteristics and wood properties of Norway spruce. Tappi 1964, 47, 499–504. [Google Scholar]
- Ahmad, S.S. Variation in tracheid dimensions within a single stem of fir. Pak. J. For. 1970, 20, 89–109. [Google Scholar]
- Sudo, S. Variation in tracheid length in Akamatsu (Pinus densiflora Sieb. et Zucc.). Mokuzai Gakkaishi 1973, 19, 569–575. [Google Scholar]
- Herman, M.; Dutilleul, P.; Avella-Shaw, T. Intra-ring and inter-ring variation of tracheid length in fast-grown versus slow-grown Norway spruces (Picea abies). IAWA J. 1998, 19, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.Y.; Zhang, S.Y.; Mansfield, S.D. The effects of initial spacing on wood density, fibre and pulp properties in jack pine (Pinus banksiana Lamb.). Holzforschung 2004, 58, 455–463. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Acronym | Description |
|---|---|
| RTL | Ring tracheid length |
| EWTL | Earlywood tracheid length |
| LWTL | Latewood tracheid length |
| ARW | Annual ring width |
| EWW | Earlywood width |
| LWW | Latewood width |
| Part 1—for All Annual Growth Rings per Site | |||||||
|---|---|---|---|---|---|---|---|
| Site | Annual Growth Ring | Property | |||||
| EWTL (mm) | LWTL (mm) | RTL (mm) | |||||
| Mean | SD | Mean | SD | Mean | SD | ||
| VG | 3 | 1.05 | 0.29 | 1.28 | 0.49 | 1.17 | 0.42 |
| 5 | 1.52 | 0.49 | 1.83 | 0.54 | 1.67 | 0.54 | |
| 10 | 2.07 | 0.61 | 2.96 | 0.63 | 2.51 | 0.76 | |
| 15 | 2.65 | 0.70 | 2.80 | 0.68 | 2.73 | 0.69 | |
| 20 | 3.55 | 0.65 | 3.26 | 1.00 | 3.41 | 0.85 | |
| 25 | 3.47 | 0.78 | 3.94 | 0.70 | 3.71 | 0.78 | |
| 30 | 3.78 | 0.93 | 3.34 | 0.79 | 3.56 | 0.89 | |
| 35 | 3.35 | 0.88 | 3.60 | 0.81 | 3.47 | 0.85 | |
| 40 | 3.04 | 0.55 | 3.27 | 0.80 | 3.15 | 0.69 | |
| 45 | 3.17 | 0.68 | 3.44 | 0.76 | 3.30 | 0.73 | |
| PO | 3 | 1.53 | 0.50 | 1.66 | 0.56 | 1.59 | 0.53 |
| 5 | 1.76 | 0.55 | 1.76 | 0.66 | 1.76 | 0.61 | |
| 10 | 2.40 | 0.53 | 2.51 | 0.76 | 2.45 | 0.66 | |
| 15 | 2.90 | 0.58 | 3.22 | 0.94 | 3.06 | 0.80 | |
| 20 | 2.85 | 0.71 | 3.34 | 0.77 | 3.09 | 0.78 | |
| 25 | 3.08 | 0.49 | 2.85 | 0.87 | 2.97 | 0.72 | |
| 30 | 2.88 | 0.72 | 3.28 | 0.85 | 3.08 | 0.81 | |
| 35 | 3.17 | 0.68 | 3.47 | 0.85 | 3.32 | 0.78 | |
| 40 | 2.62 | 0.51 | 2.66 | 0.73 | 2.64 | 0.63 | |
| VŽ | 3 | 1.5 | 0.36 | 1.68 | 0.54 | 1.59 | 0.47 |
| 5 | 1.74 | 0.42 | 1.93 | 0.5 | 1.83 | 0.47 | |
| 10 | 2.57 | 0.71 | 2.59 | 0.62 | 2.58 | 0.66 | |
| 15 | 2.66 | 0.64 | 3.13 | 0.61 | 2.9 | 0.67 | |
| 20 | 2.81 | 0.66 | 3.00 | 0.71 | 2.91 | 0.69 | |
| 25 | 3.05 | 0.66 | 2.61 | 0.64 | 2.83 | 0.69 | |
| 30 | 2.86 | 0.67 | 3.11 | 0.87 | 2.99 | 0.79 | |
| 35 | 2.84 | 0.65 | 3.15 | 0.94 | 2.99 | 0.82 | |
| 40 | 2.58 | 0.77 | 2.69 | 0.69 | 2.63 | 0.73 | |
| 45 | 2.86 | 0.66 | 3.00 | 0.72 | 2.93 | 0.69 | |
| 50 | 2.69 | 0.65 | 3.54 | 0.98 | 3.12 | 0.93 | |
| 60 | 2.83 | 0.54 | 2.83 | 0.75 | 2.83 | 0.65 | |
| 70 | 2.92 | 0.80 | 2.98 | 0.61 | 2.95 | 0.71 | |
| Part 2—All Data until the 40th Annual Growth Ring per Site | |||||||
| Site | Property | ||||||
| EWTL (mm) | LWTL (mm) | RTL (mm) | |||||
| Mean | SD | Mean | SD | Mean | SD | ||
| VG | 2.72 | 1.14 | 2.92 | 1.08 | 2.82 | 1.11 | |
| PO | 2.58 | 0.80 | 2.75 | 1.00 | 2.66 | 0.99 | |
| VŽ | 2.51 | 0.80 | 2.65 | 0.85 | 2.58 | 1.00 | |
| Site | Source | SS | d.f. | MS | F | p |
|---|---|---|---|---|---|---|
| VG | Annual growth rings × zone | 54.93 | 9 | 6.10 | 12.30 | 0.000000 |
| PO | Annual growth rings × zone | 18.58 | 8 | 2.32 | 4.95 | 0.000005 |
| VŽ | Annual growth rings × zone | 45.91 | 12 | 3.83 | 8.22 | 0.000000 |
| Site | Sources | SS | d.f. | MS | F | p |
|---|---|---|---|---|---|---|
| VG | Annual growth rings | 1179.66 | 9 | 131.07 | 249.15 | 0.000000 |
| Tree | 5.75 | 2 | 2.88 | 4.67 | 0.010522 | |
| PO | Annual growth rings | 548.69 | 8 | 68.59 | 139.79 | 0.000000 |
| Tree | 16.18 | 2 | 8.09 | 14.04 | 0.000002 | |
| VŽ | Annual growth rings | 463.95 | 12 | 38.66 | 92.43 | 0.000000 |
| Tree | 7.83 | 2 | 3.92 | 7.54 | 0.000719 |
| Source | SS | d.f. | MS | F | p | Tukey Post hoc | ||
|---|---|---|---|---|---|---|---|---|
| Site | p | |||||||
| Site | 47.29 | 2 | 23.65 | 42.29 | 0.000000 | VG | PO | 0.000022 |
| VŽ | 0.000022 | |||||||
| PO | VG | 0.000022 | ||||||
| VŽ | 0.006616 | |||||||
| VŽ | VG | 0.000022 | ||||||
| PO | 0.006616 | |||||||
| Site | Annual Growth Ring | Property | |||||
|---|---|---|---|---|---|---|---|
| EWW (mm) | LWW (mm) | ARW (mm) | |||||
| Mean | SD | Mean | SD | Mean | SD | ||
| VG | 3 | 2.91 | 3.34 | 1.84 | 0.44 | 6.13 | 2.09 |
| 5 | 4.76 | 1.26 | 2.92 | 1.79 | 7.47 | 0.43 | |
| 10 | 1.84 | 0.74 | 4.00 | 1.00 | 6.25 | 0.24 | |
| 15 | 2.70 | 0.20 | 2.73 | 0.76 | 5.52 | 0.20 | |
| 20 | 3.44 | 0.66 | 2.64 | 0.57 | 5.75 | 0.47 | |
| 25 | 1.81 | 0.17 | 2.01 | 0.81 | 3.87 | 0.15 | |
| 30 | 2.22 | 0.13 | 3.08 | 0.75 | 5.34 | 1.75 | |
| 35 | 1.52 | 0.31 | 1.39 | 0.48 | 2.73 | 0.29 | |
| 40 | 1.09 | 0.28 | 0.62 | 0.35 | 1.59 | 0.28 | |
| All | 2.60 | 1.57 | 2.36 | 1.21 | 4.96 | 2.24 | |
| PO | 3 | 2.61 | 0.39 | 1.77 | 1.12 | 4.38 | 1.16 |
| 5 | 2.91 | 1.17 | 3.99 | 3.19 | 6.90 | 4.22 | |
| 10 | 5.06 | 0.93 | 3.47 | 0.84 | 8.54 | 0.40 | |
| 15 | 2.16 | 0.87 | 1.69 | 0.19 | 3.86 | 1.01 | |
| 20 | 2.93 | 0.90 | 2.48 | 1.14 | 5.42 | 1.78 | |
| 25 | 3.62 | 0.09 | 3.28 | 1.12 | 6.90 | 1.16 | |
| 30 | 2.69 | 0.77 | 1.57 | 0.60 | 4.25 | 1.34 | |
| 35 | 2.76 | 0.22 | 1.62 | 0.48 | 4.38 | 0.69 | |
| 40 | 1.76 | 1.29 | 1.05 | 1.01 | 2.81 | 2.28 | |
| All | 2.94 | 1.14 | 2.33 | 1.49 | 5.27 | 2.36 | |
| VŽ | 3 | 3.38 | 1.19 | 1.62 | 1.66 | 5.00 | 1.47 |
| 5 | 1.23 | 0.69 | 1.39 | 1.30 | 2.63 | 1.99 | |
| 10 | 1.14 | 0.51 | 1.08 | 0.56 | 2.22 | 0.88 | |
| 15 | 1.33 | 0.49 | 1.65 | 0.95 | 2.98 | 1.43 | |
| 20 | 1.96 | 1.05 | 2.02 | 0.89 | 3.98 | 1.92 | |
| 25 | 2.06 | 0.56 | 1.65 | 0.57 | 3.71 | 1.06 | |
| 30 | 1.45 | 0.38 | 1.60 | 0.93 | 3.05 | 0.91 | |
| 35 | 1.77 | 0.62 | 1.71 | 0.17 | 3.48 | 0.73 | |
| 40 | 1.87 | 0.44 | 1.97 | 0.58 | 3.83 | 0.99 | |
| All | 1.80 | 0.88 | 1.63 | 0.83 | 3.43 | 1.37 | |
| Site | Correlation Coefficient | ||
|---|---|---|---|
| RTL–ARW | EWTL–EWW | LWTL–LWW | |
| r | r | r | |
| VG | −0.4495 | −0.4765 | −0.1798 |
| PO | −0.1492 | 0.0358 | −0.2328 |
| VŽ | −0.2163 | −0.3836 | 0.1277 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ištok, I.; Sedlar, T.; Orešković, G.; Jambreković, B. The Variations in Tracheid Length of Pseudotsuga menziesii (Mirb.) Franco Wood in Relation to Cambium Age, Site, and Growth. Forests 2023, 14, 1165. https://doi.org/10.3390/f14061165
Ištok I, Sedlar T, Orešković G, Jambreković B. The Variations in Tracheid Length of Pseudotsuga menziesii (Mirb.) Franco Wood in Relation to Cambium Age, Site, and Growth. Forests. 2023; 14(6):1165. https://doi.org/10.3390/f14061165
Chicago/Turabian StyleIštok, Iva, Tomislav Sedlar, Gordana Orešković, and Branimir Jambreković. 2023. "The Variations in Tracheid Length of Pseudotsuga menziesii (Mirb.) Franco Wood in Relation to Cambium Age, Site, and Growth" Forests 14, no. 6: 1165. https://doi.org/10.3390/f14061165
APA StyleIštok, I., Sedlar, T., Orešković, G., & Jambreković, B. (2023). The Variations in Tracheid Length of Pseudotsuga menziesii (Mirb.) Franco Wood in Relation to Cambium Age, Site, and Growth. Forests, 14(6), 1165. https://doi.org/10.3390/f14061165