Autographa Californica Multiple Nucleopolyhedrovirus Enters Host Cells via Clathrin-Mediated Endocytosis and Direct Fusion with the Plasma Membrane


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Construction of Clonal Cell Lines Expressing Endocytic Markers
2.3. One-Step Growth Curve Assay
2.4. Live-Cell Imaging
2.5. Transmission Electron Microscopy
3. Results
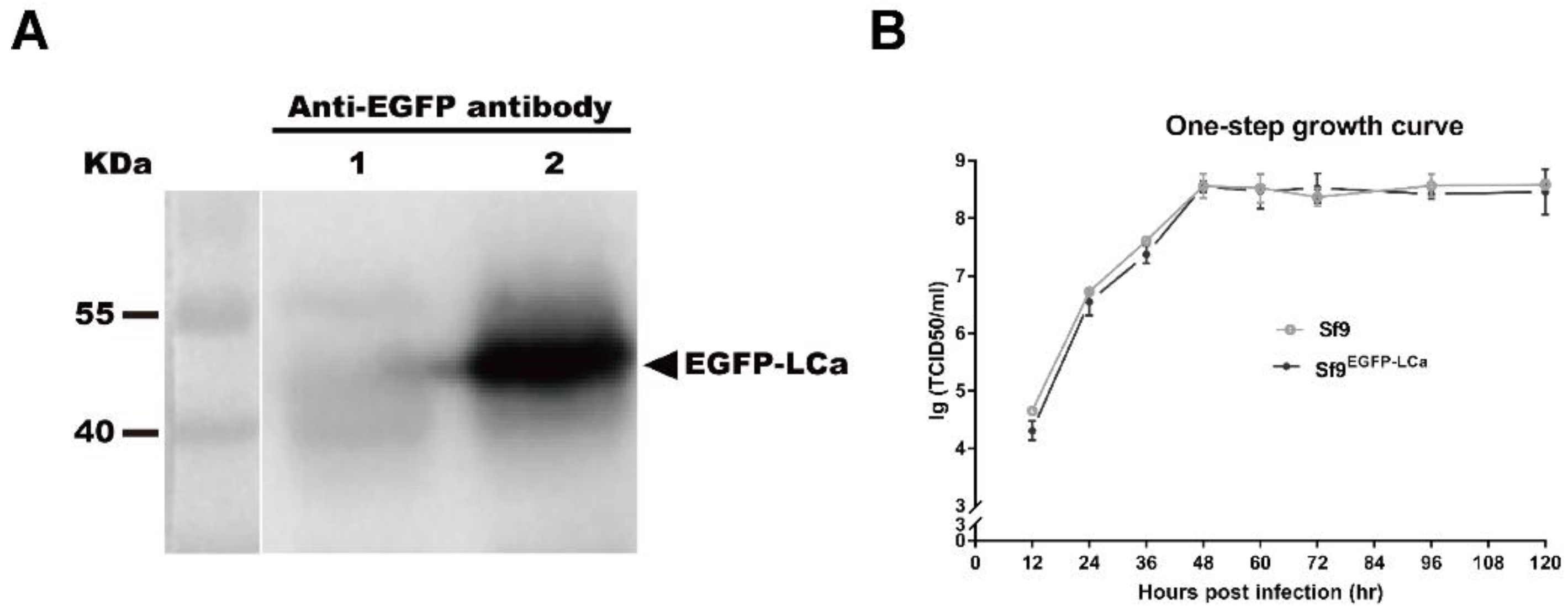
3.1. Construction and Characterization of Recombinant AcMNPV and Clonal Cell Lines
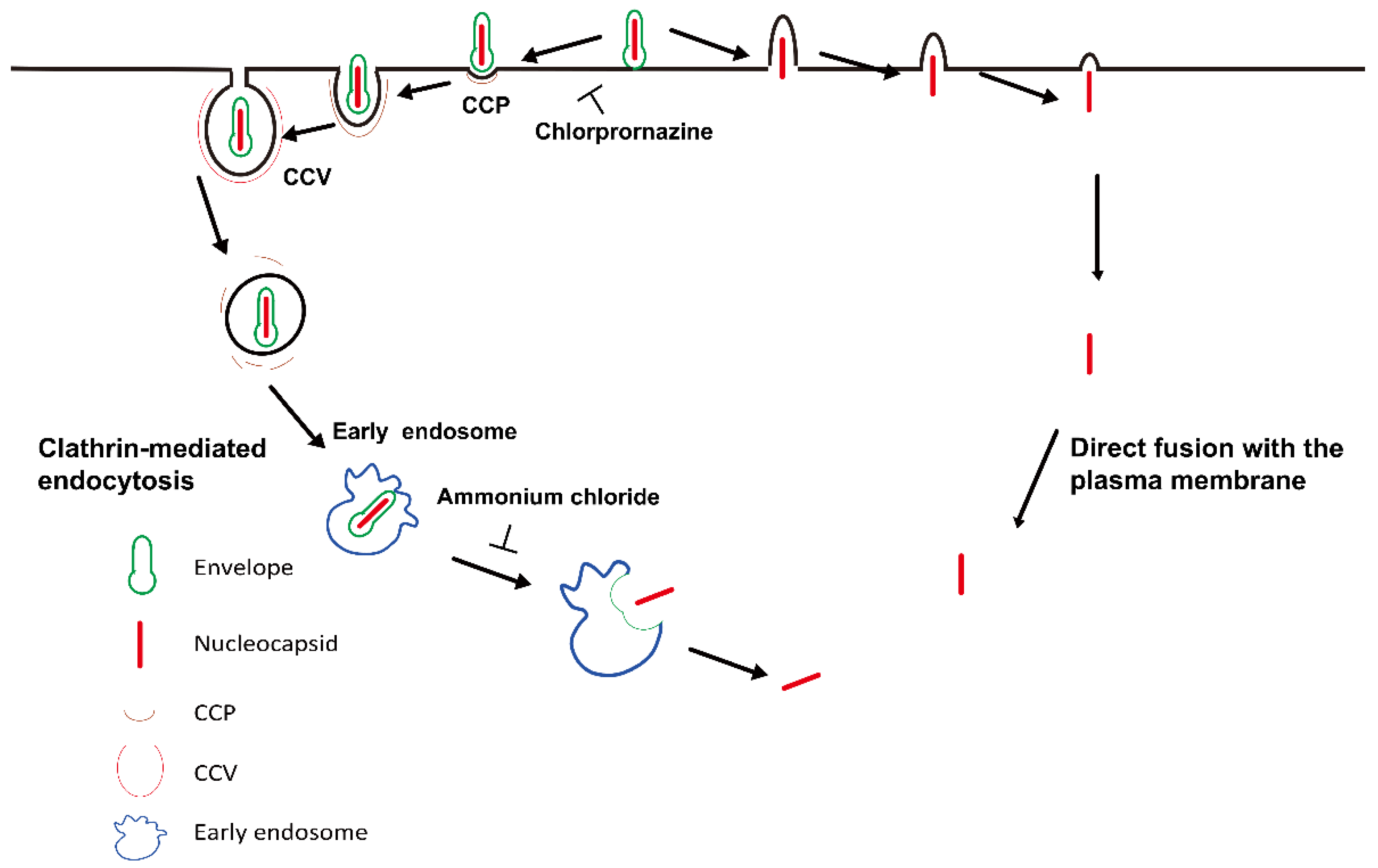
3.2. AcMNPV Enters Cells via Clathrin-Mediated Endocytosis and a Clathrin-Independent Pathway
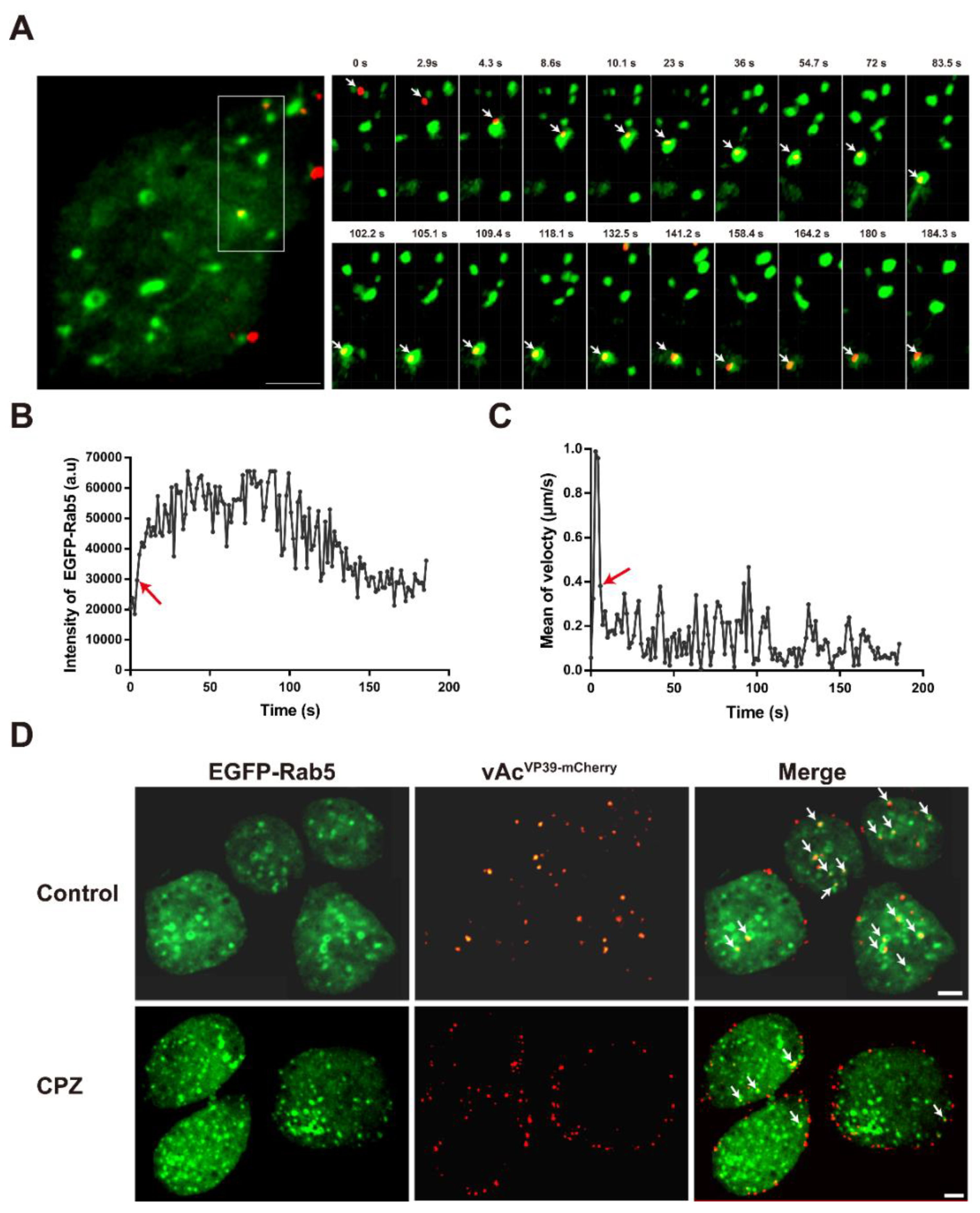
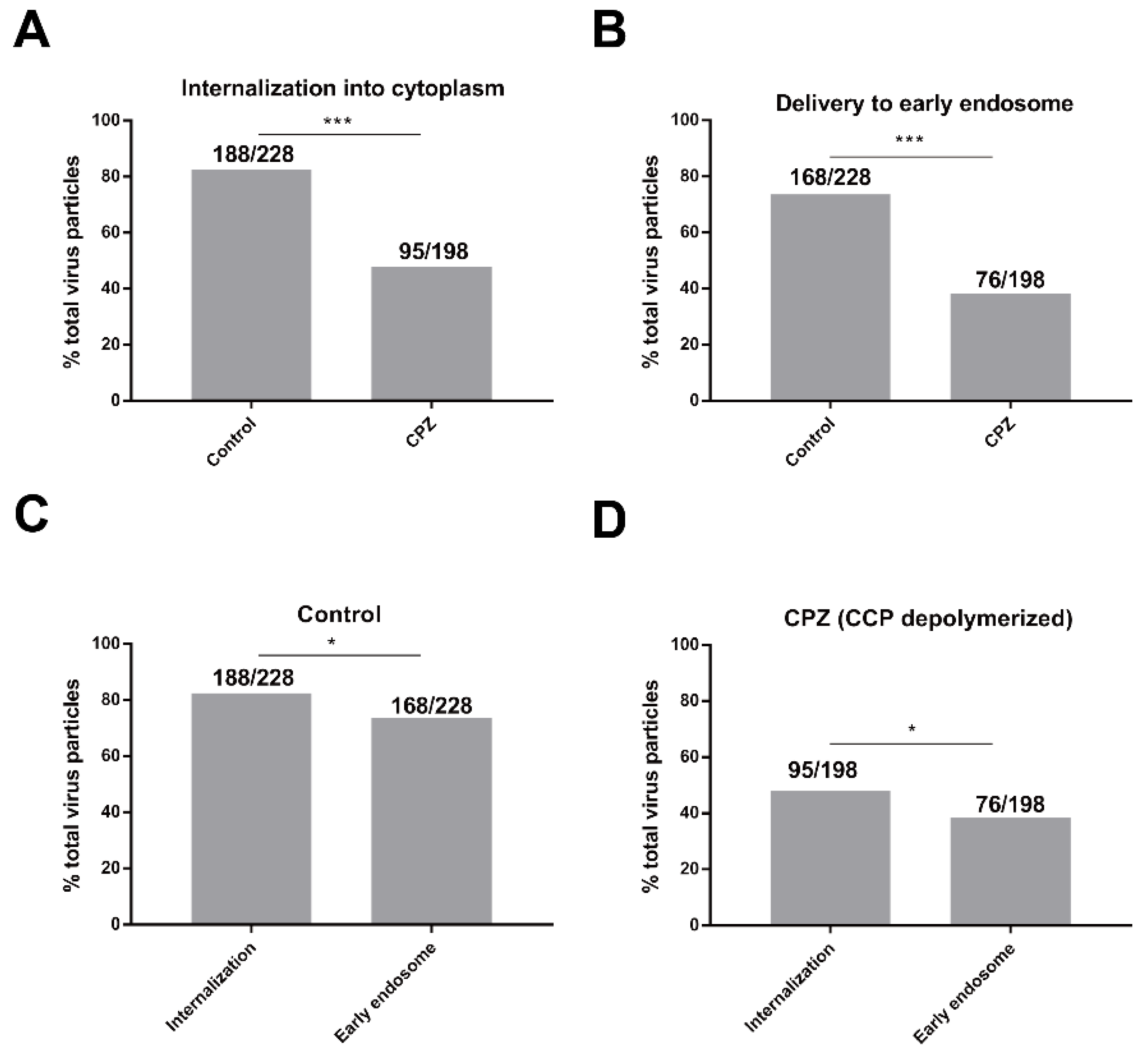
3.3. Depolymerization of CPPs Inhibits the Delivery of AcMNPV Particles into Early Endosomes
3.4. AcMNPV Enters Cells via an Early Endosome-Independent Pathway
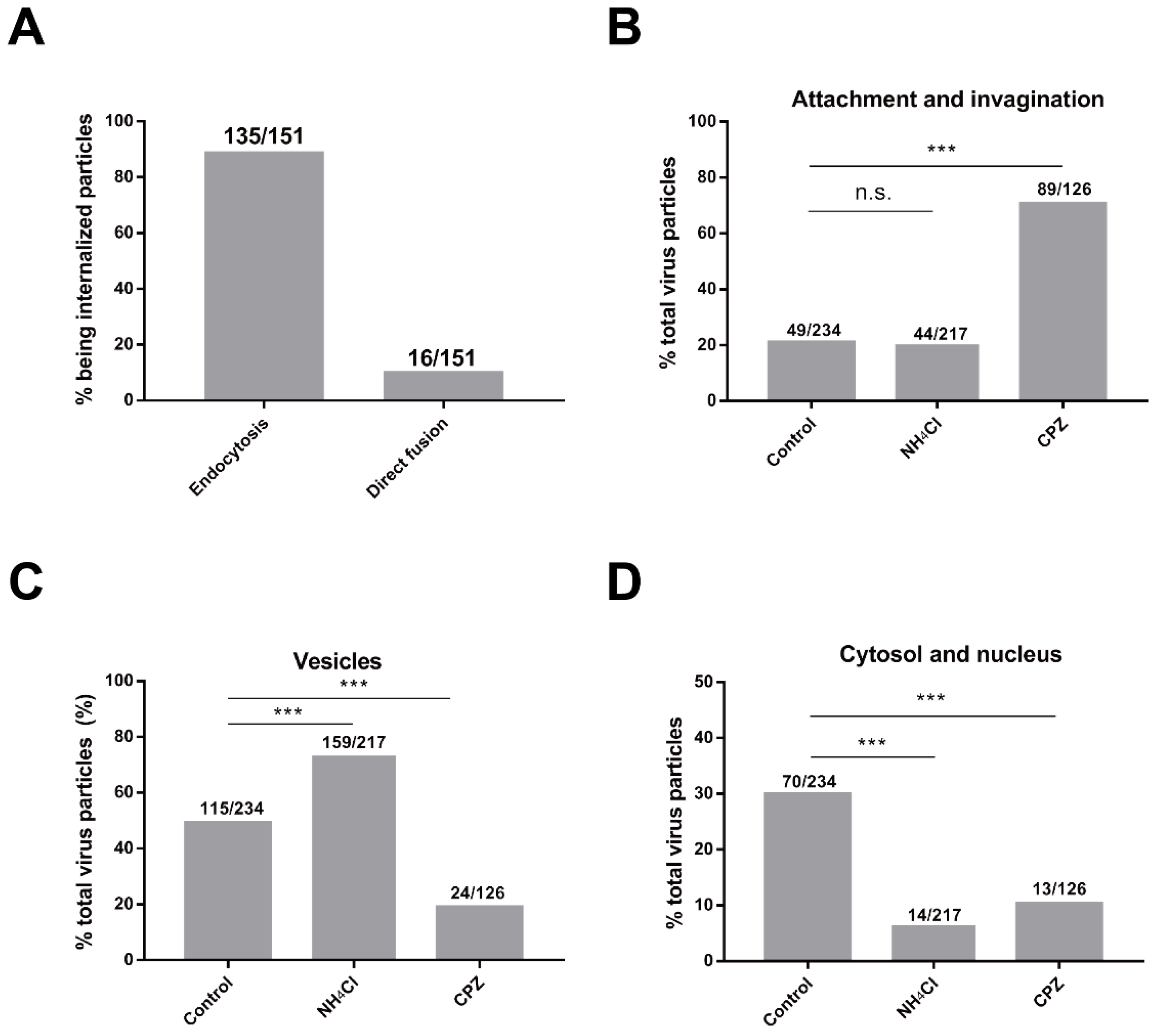
3.5. AcMNPV Enters Cells via Endocytosis and Direct Fusion with the Plasma Membrane
3.6. Depolymerization of CPPs Inhibits AcMNPV Internalization and Inhibition of Endosomal Acidification Blocks Particles out of Vesicles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.L. History and Current Status of Development and Use of Viral Insecticides in China. Viruses 2015, 7, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Xing, L.; Wang, M.; Wang, X.; Yin, M.; Wang, Q.; Hu, Z.; Zou, Z. Inhibition of melanization by serpin-5 and serpin-9 promotes baculovirus infection in cotton bollworm Helicoverpa armigera. PLoS Pathog. 2017, 13, e1006645. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.C.; Aksular, M.; Graves, L.P.; Irons, S.L.; Possee, R.D.; King, L.A. Overview of the Baculovirus Expression System. Curr. Protoc. Protein Sci. 2018, 91, 5.4.1–5.4.6. [Google Scholar] [PubMed]
- Van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus-insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Theze, J.; Lopez-Vaamonde, C.; Cory, J.S.; Herniou, E.A. Biodiversity, Evolution and Ecological Specialization of Baculoviruses: A Treasure Trove for Future Applied Research. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Volkman, L.E.; Summers, M.D. Autographa californica nuclear polyhedrosis virus: Comparative infectivity of the occluded, alkali-liberated, and nonoccluded forms. J. Invertebr. Pathol. 1977, 30, 102–103. [Google Scholar] [CrossRef]
- Goley, E.D.; Ohkawa, T.; Mancuso, J.; Woodruff, J.B.; D’Alessio, J.A.; Cande, W.Z.; Volkman, L.E.; Welch, M.D. Dynamic nuclear actin assembly by Arp2/3 complex and a baculovirus WASP-like protein. Science 2006, 314, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Zhang, Y.; Hu, Y.; Hu, X.; Zhou, Y.; Zhao, H.; Pei, R.; Wu, C.; Chen, J.; Zhao, H.; et al. Autographa californica Multiple Nucleopolyhedrovirus Ac34 Protein Retains Cellular Actin-Related Protein 2/3 Complex in the Nucleus by Subversion of CRM1-Dependent Nuclear Export. PLoS Pathog. 2016, 12, e1005994. [Google Scholar] [CrossRef] [PubMed]
- Ishimwe, E.; Hodgson, J.J.; Passarelli, A.L. Expression of the Cydia pomonella granulovirus matrix metalloprotease enhances Autographa californica multiple nucleopolyhedrovirus virulence and can partially substitute for viral cathepsin. Virology 2015, 481, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, L.; Zhao, H.; Liu, W. Disruption of Autographa Californica Multiple Nucleopolyhedrovirus ac111 Results in Reduced per os Infectivity in a Host-Dependent Manner. Viruses 2018, 10, 527. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Blissard, G.W.; Theilmann, D.A. Trichoplusia ni Kinesin-1 Associates with Autographa californica Multiple Nucleopolyhedrovirus Nucleocapsid Proteins and Is Required for Production of Budded Virus. J. Virol. 2016, 90, 3480–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boogaard, B.; van Oers, M.M.; van Lent, J.W.M. An Advanced View on Baculovirus per Os Infectivity Factors. Insects 2018, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Blissard, G.W.; Theilmann, D.A. Baculovirus Entry and Egress from Insect Cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Volkman, L.E.; Goldsmith, P.A. Mechanism of Neutralization of Budded Autographa-Californica Nuclear Polyhedrosis-Virus by a Monoclonal-Antibody—Inhibition of Entry by Adsorptive Endocytosis. Virology 1985, 143, 185–195. [Google Scholar] [CrossRef]
- Hefferon, K.L.; Oomens, A.G.P.; Monsma, S.A.; Finnerty, C.M.; Blissard, G.W. Host cell receptor binding by baculovirus GP64 and kinetics of virion entry. Virology 1999, 258, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Long, G.; Pan, X.Y.; Kormelink, R.; Vlak, J.M. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef] [PubMed]
- Volkman, L.E.; Goldsmith, P.A.; Hess, R.T. Alternate Pathway of Entry of Budded Autographa-Californica Nuclear Polyhedrosis-Virus—Fusion at the Plasma-Membrane. Virology 1986, 148, 288–297. [Google Scholar] [CrossRef]
- Dong, S.C.; Wang, M.L.; Qiu, Z.J.; Deng, F.; Vlak, J.M.; Hu, Z.H.; Wang, H.L. Autographa californica Multicapsid Nucleopolyhedrovirus Efficiently Infects Sf9 Cells and Transduces Mammalian Cells via Direct Fusion with the Plasma Membrane at Low pH. J. Virol. 2010, 84, 5351–5359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyauchi, K.; Kim, Y.; Latinovic, O.; Morozov, V.; Melikyan, G.B. HIV enters cells via endocytosis and dynamin-dependent fusion with endosomes. Cell 2009, 137, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.; Helenius, A. Virus entry: Open sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.J.; Lakadamyali, M.; Zhang, F.; Zhuang, X.W. Assembly of endocytic machinery around individual influenza viruses during viral entry. Nat. Struct. Mol. Biol. 2004, 11, 567–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X.W. Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes. Cell 2006, 124, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Rink, J.; Ghigo, E.; Kalaidzidis, Y.; Zerial, M. Rab conversion as a mechanism of progression from early to late endosomes. Cell 2005, 122, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, E.; Tscherne, D.M.; Wienholts, M.J.; Cobos-Jimenez, V.; Scholte, F.; Garcia-Sastre, A.; Rottier, P.J.M.; de Haan, C.A.M. Dissection of the Influenza A Virus Endocytic Routes Reveals Macropinocytosis as an Alternative Entry Pathway. PLoS Pathog. 2011, 7, e1001329. [Google Scholar] [CrossRef] [PubMed]
- Damm, E.M.; Pelkmans, L.; Kartenbeck, J.; Mezzacasa, A.; Kurzckalia, T.; Helenius, A. Clathrin- and caveolin-1-independent endocytosis: Entry of simian virus 40 into cells devoid of caveolae. J. Cell Biol. 2005, 168, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Pelkmans, L.; Puntener, D.; Helenius, A. Local actin polymerization and dynamin recruitment in SV40-induced internalization of caveolae. Science 2002, 296, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Cureton, D.K.; Massol, R.H.; Saffarian, S.; Kirchhausen, T.L.; Whelan, S.P.J. Vesicular Stomatitis Virus Enters Cells through Vesicles Incompletely Coated with Clathrin That Depend upon Actin for Internalization. PLoS Pathog. 2009, 5, e1000394. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, B.; Zhuang, X.W. Virus trafficking—Learning from single-virus tracking. Nat. Rev. Microbiol. 2007, 5, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.H.; Burckhardt, C.J.; Yakimovich, A.; Greber, U.F. Imaging, Tracking and Computational Analyses of Virus Entry and Egress with the Cytoskeleton. Viruses 2018, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Borrenberghs, D.; Rocha, S.; Hendrix, J. Single Viruses on the Fluorescence Microscope: Imaging Molecular Mobility, Interactions and Structure Sheds New Light on Viral Replication. Viruses 2018, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.Q.; Wang, H.J.; Li, X.Y.; Fang, N.; Yang, S.L.; Liu, H.Y.; Kang, X.N.; Sun, X.L.; Ji, S.P. Cloning and Characterization of Sf9 Cell Lamin and the Lamin Conformational Changes during Autographa californica multiple nucleopolyhedrovirus Infection. Viruses 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Xu, K.Y.; Wei, D.H.; Wu, W.B.; Yang, K.; Yuan, M.J. Baculovirus infection induces disruption of the nuclear lamina. Sci. Rep. 2017, 7, 7823. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Lin, Y.; Zheng, Z.H.; Zhang, Z.L.; Zhang, L.J.; Wang, L.Y.; Wang, H.Z.; Pang, D.W. Labeling the nucleocapsid of enveloped baculovirus with quantum dots for single-virus tracking. Biomaterials 2014, 35, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.X.; Zheng, Z.H.; Liu, S.L.; Lu, W.; Zhang, Z.F.; Zhang, C.L.; Zhou, P.; Zhang, Y.; Long, G.; He, Z.K.; et al. Self-biotinylation and site-specific double labeling of baculovirus using quantum dots for single-virus in-situ tracking. Biomaterials 2013, 34, 7506–7518. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, M.; Li, W.; Zhang, Z.; Zhang, X.; Tan, T.; Zhang, X.E.; Cui, Z. Live cell imaging of single genomic loci with quantum dot-labeled TALEs. Nat. Commun. 2017, 8, 15318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakadamyali, M.; Rust, M.J.; Babcock, H.P.; Zhuang, X.W. Visualizing infection of individual influenza viruses. Proc. Natl. Acad. Sci. USA 2003, 100, 9280–9285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Blanc, I.; Luyet, P.P.; Pons, V.; Ferguson, C.; Emans, N.; Petiot, A.; Mayran, N.; Demaurex, N.; Faure, J.; Sadoul, R.; et al. Endosome-to-cytosol transport of viral nucleocapsids. Nat. Cell Biol. 2005, 7, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loo, N.D.; Fortunati, E.; Ehlert, E.; Rabelink, M.; Grosveld, F.; Scholte, B.J. Baculovirus infection of nondividing mammalian cells: Mechanisms of entry and nuclear transport of capsids. J. Virol. 2001, 75, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Emery, V.C. Baculovirus expression vectors. Methods. Mol. Biol. 1992, 8, 319–326. [Google Scholar] [PubMed]
- Li, Y.M.; Shen, S.; Hu, L.B.; Deng, F.; Vlak, J.M.; Hu, Z.H.; Wang, H.L.; Wang, M.L. The Functional Oligomeric State of Tegument Protein GP41 Is Essential for Baculovirus Budded Virion and Occlusion-Derived Virion Assembly. J. Virol. 2018, 92, JVI-02083. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M.; Boll, W.; van Oijen, A.; Hariharan, R.; Chandran, K.; Nibert, M.L.; Kirchhausen, T. Endocytosis by random initiation and stabilization of clathrin-coated pits. Cell 2004, 118, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Xu, C.; Lei, C.; Hu, J.; Zheng, Z.; Peng, K.; Wang, H.; Sun, X. Actin and microtubules are essential for baculovirus nucleocapsid release from within early endosomes. mBio 2018. under review. [Google Scholar]
- Francis, A.C.; Melikyan, G.B. Live-Cell Imaging of Early Steps of Single HIV-1 Infection. Viruses 2018, 10, 275. [Google Scholar] [CrossRef] [PubMed]
- Van der Schaar, H.M.; Rust, M.J.; Chen, C.; van der Ende-Metselaar, H.; Wilschut, J.; Zhuang, X.W.; Smit, J.M. Dissecting the Cell Entry Pathway of Dengue Virus by Single-Particle Tracking in Living Cells. PLoS Pathog. 2008, 4, e1000244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.B.; Liu, Y.; Liu, S.L.; Pang, D.W.; Xiao, G.F. Clathrin-Mediated Endocytosis in Living Host Cells Visualized through Quantum Dot Labeling of Infectious Hematopoietic Necrosis Virus. J. Virol. 2011, 85, 6252–6262. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, F.; Xu, C.; Lei, C.; Hu, J.; Sun, X. Autographa Californica Multiple Nucleopolyhedrovirus Enters Host Cells via Clathrin-Mediated Endocytosis and Direct Fusion with the Plasma Membrane. Viruses 2018, 10, 632. https://doi.org/10.3390/v10110632
Qin F, Xu C, Lei C, Hu J, Sun X. Autographa Californica Multiple Nucleopolyhedrovirus Enters Host Cells via Clathrin-Mediated Endocytosis and Direct Fusion with the Plasma Membrane. Viruses. 2018; 10(11):632. https://doi.org/10.3390/v10110632
Chicago/Turabian StyleQin, Fujun, Congrui Xu, Chengfeng Lei, Jia Hu, and Xiulian Sun. 2018. "Autographa Californica Multiple Nucleopolyhedrovirus Enters Host Cells via Clathrin-Mediated Endocytosis and Direct Fusion with the Plasma Membrane" Viruses 10, no. 11: 632. https://doi.org/10.3390/v10110632
APA StyleQin, F., Xu, C., Lei, C., Hu, J., & Sun, X. (2018). Autographa Californica Multiple Nucleopolyhedrovirus Enters Host Cells via Clathrin-Mediated Endocytosis and Direct Fusion with the Plasma Membrane. Viruses, 10(11), 632. https://doi.org/10.3390/v10110632