Cytokine Signature Associated with Disease Severity in Dengue

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Detection of Dengue Virus
2.3. Analyses of 41plex Cytokines Using Multiplex Luminex System
2.4. Principal Component Analysis (PCA), Partial Least Squares Discriminant Analysis (PLS-DA)
2.5. Statistical Analysis
3. Results
3.1. Population Demographics
3.2. Dengue Testing
3.3. Sequences and NCBI Accession Number
3.4. Multiplex Cytokine Bead Array (Inflammatory Markers as Predictors of The Dengue Severity) and Principal Component Assay
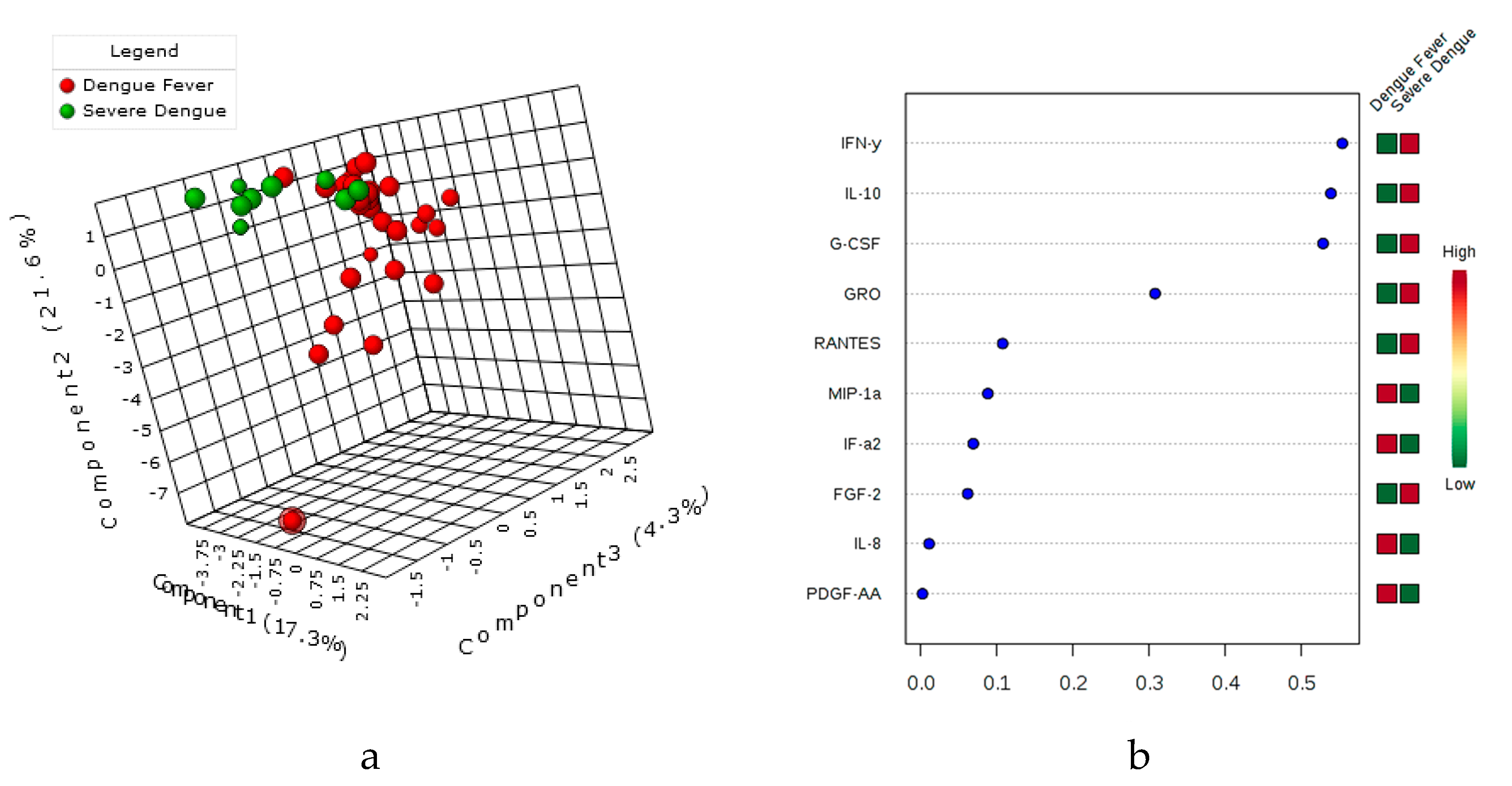
3.5. Principal Component Assay (PCA), sparsePLS-DA & the variable’s importance VIP Score
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
Presented in Part
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenbach, B.D.; Murray, C.L.; Thiel, H.J.; Rice, C.M. Flaviviridiae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer, Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 712–746. [Google Scholar]
- Halstead, S.B. Pathogenesis of dengue: Challenges to molecular biology. Science 1988, 239, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A.L. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Screaton, G.; Mongkolsapaya, J.; Yacoub, S.; Roberts, C. New insights into the immunopathology and control of dengue virus infection. Nat. Rev. Immunol. 2015, 15, 745–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waggoner, J.J.; Balmaseda, A.; Gresh, L.; Sahoo, M.K.; Montoya, M.; Wang, C.; Abeynayake, J.; Kuan, G.; Pinsky, B.A.; Harris, E. Homotypic Dengue Virus Reinfections in Nicaraguan Children. J. Infect. Dis. 2016, 214, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [Green Version]
- Bozza, F.A.; Cruz, O.G.; Zagne, S.M.; Azeredo, E.L.; Nogueira, R.M.; Assis, E.F.; Bozza, P.T.; Kubelka, C.F. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 2008, 8, 86. [Google Scholar] [CrossRef]
- Soo, K.M.; Khalid, B.; Ching, S.M.; Tham, C.L.; Basir, R.; Chee, H.Y. Meta-analysis of biomarkers for severe dengue infections. PeerJ 2017, 5, e3589. [Google Scholar] [CrossRef]
- Costa, V.V.; Fagundes, C.T.; Souza, D.G.; Teixeira, M.M. Inflammatory and innate immune responses in dengue infection: Protection versus disease induction. Am. J. Pathol. 2013, 182, 1950–1961. [Google Scholar] [CrossRef] [PubMed]
- Rathakrishnan, A.; Wang, S.M.; Hu, Y.; Khan, A.M.; Ponnampalavanar, S.; Lum, L.C.; Manikam, R.; Sekaran, S.D. Cytokine expression profile of dengue patients at different phases of illness. PLoS ONE 2012, 7, e52215. [Google Scholar] [CrossRef] [PubMed]
- Won, J.H.; Goldberger, O.; Shen-Orr, S.S.; Davis, M.M.; Olshen, R.A. Significance analysis of xMap cytokine bead arrays. Proc. Natl. Acad. Sci. USA 2012, 109, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Recommendations of the Strategic Advisory Group of Experts on Immunization (SAGE) on the Use of Dengvaxia. Available online: http://www.who.int/immunization/diseases/dengue/revised_SAGE_recommendations_dengue_vaccines_apr2018/en/ (accessed on 13 September 2018).
- WHO/TDR. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; WHO/TDR: Geneva, Switzerland, 2009; Available online: http://www.who.int/tdr/publications/documents/dengue-diagnosis.pdf (accessed on 27 June 2018).
- Alexander, N.; Balmaseda, A.; Coelho, I.C.; Dimaano, E.; Hien, T.T.; Hung, N.T.; Janisch, T.; Kroeger, A.; Lum, L.C.; Martinez, E.; et al. Multicentre prospective study on dengue classification in four South-east Asian and three Latin American countries. Trop. Med. Int. Health 2011, 16, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [PubMed]
- Saswat, T.; Kumar, A.; Kumar, S.; Mamidi, P.; Muduli, S.; Debata, N.K.; Pal, N.S.; Pratheek, B.M.; Chattopadhyay, S. High rates of co-infection of Dengue and Chikungunya virus in Odisha and Maharashtra, India during 2013. Infect. Genet. Evol. 2015, 35, 134–141. [Google Scholar] [CrossRef]
- Patro, A.R.K.; Prusty, B.K.; Gaikwad, S.; Singh, D.K.; Mohanty, S.; Das, B.K.; Ravindran, B. Identification of inflammatory biomarkers in dengue disease severity in eastern India. In Proceedings of the Joint American Society for Cell Biology (ASCB) and European Molecular Biology Organization (EMBO) 2017 Annual Meeting, Pennsylvania Convention Center, Philadelphia, PA, USA, 2–6 December 2017; Abstract B837. p. 3542. [Google Scholar]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A.; Nisalak, A. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef]
- OhAinle, M.; Balmaseda, A.; Macalalad, A.R.; Tellez, Y.; Zody, M.C.; Saborío, S.; Nuñez, A.; Lennon, N.J.; Birren, B.W.; Gordon, A.; et al. Dynamics of Dengue Disease Severity Determined by the Interplay Between Viral Genetics and Serotype-Specific Immunity. Sci. Transl. Med. 2011, 3, 114ra128. [Google Scholar] [CrossRef]
- Rico-Hesse, R.; Harrison, L.M.; Salas, R.A.; Tovar, D.; Nisalak, A.; Ramos, C.; Boshell, J.; de Mesa, M.T.; Nogueira, R.M.; da Rosa, A.T. Origins of dengue type 2 viruses associated with increased pathogenicity in the Americas. Virology 1997, 230, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Srikiatkhachorn, A.; Mathew, A.; Rothman, A.L. Immune-mediated cytokine storm and its role in severe dengue. Semin. Immunopathol. 2017, 39, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Green, S.; Vaughn, D.W.; Kalayanarooj, S.; Nimmannitya, S.; Suntayakorn, S.; Nisalak, A.; Rothman, A.L.; Ennis, F.A. Elevated plasma interleukin-10 levels in acute dengue correlate with disease severity. J. Med. Virol. 1999, 59, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Butthep, P.; Chunhakan, S.; Yoksan, S.; Tangnararatchakit, K.; Chuansumrit, A. Alteration of cytokines and chemokines during febrile episodes associated with endothelial cell damage and plasma leakage in dengue hemorrhagic fever. Pediatr. Infect. Dis. J. 2012, 31, e232–e238. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Lei, H.Y.; Nguyen, T.L.; Lin, Y.S.; Huang, K.J.; Le, B.L.; Lin, C.F.; Yeh, T.M.; Do, Q.H.; Vu, T.Q.; et al. Dengue hemorrhagic fever in infants: A study of clinical and cytokine profiles. J. Infect. Dis. 2004, 189, 221–232. [Google Scholar] [PubMed]
- Priyadarshini, D.; Gadia, R.R.; Tripathy, A.; Gurukumar, K.R.; Bhagat, A.; Patwardhan, S.; Mokashi, N.; Vaidya, D.; Shah, P.S.; Cecilia, D. Clinical findings and pro-inflammatory cytokines in dengue patients in Western India: A facility-based study. PLoS ONE 2010, 5, e8709. [Google Scholar] [CrossRef] [PubMed]
- Malavige, G.N.; Gomes, L.; Alles, L.; Chang, T.; Salimi, M.; Fernando, S.; Nanayakkara, K.D.; Jayaratne, S.; Ogg, G.S. Serum IL-10 as a marker of severe dengue infection. BMC Infect. Dis. 2013, 13, 341. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Leong, W.Y.; Wilder-Smith, A. Markers of dengue severity: A systematic review of cytokines and chemokines. J. Gen. Virol. 2016, 97, 3103–3119. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Duong, V.; Tawfik, A.; Ung, S.; Ly, S.; Casademont, I.; Prot, M.; Courtejoie, N.; Bleakley, K.; Buchy, P.; et al. Increased adaptive immune responses and proper feedback regulation protect against clinical dengue. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Kurane, I.; Innis, B.L.; Nisalak, A.; Hoke, C.; Nimmannitya, S.; Meager, A.; Ennis, F.A. Human T cell responses to dengue virus antigens. Proliferative responses and interferon gamma production. J. Clin. Investig. 1989, 83, 506–513. [Google Scholar] [CrossRef]
- Spain-Santana, T.A.; Marglin, S.; Ennis, F.A.; Rothman, A.L. MIP-1 alpha and MIP-1 beta induction by dengue virus. J. Med. Virol. 2001, 65, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.D.; Proost, P.; Murphy, W.J.; Anver, M.; Longo, D.L.; van Damme, J.; Oppenheim, J.J. Monocyte chemotactic protein-1 (MCP-1), -2, and -3 are chemotactic for human T lymphocytes. J. Clin. Investig. 1995, 95, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Rutledge, B.; Fiorillo, J.; Ernst, C.; Grewal, I.; Flavell, R.; Gladue, R.; Rollins, B. In vivo properties of monocyte chemoattractant protein-1. J. Leukoc. Biol. 1997, 62, 577–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.R.; Liu, M.T.; Lei, H.Y.; Liu, C.C.; Wu, J.M.; Tung, Y.C.; Lin, Y.S.; Yeh, T.M.; Chen, S.H.; Liu, H.S. MCP-1, a highly expressed chemokine in dengue haemorrhagic fever/dengue shock syndrome patients, may cause permeability change, possibly through reduced tight junctions of vascular endothelium cells. J. Gen. Virol. 2006, 87, 3623–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lê Cao, K.A.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dengue Serotype N = 47 | Frequency (%) |
|---|---|
| Single Infection (N = 29) | |
| DENV-1 | 1 (3.44) |
| DENV-2 | 20 (68.86) |
| DENV-3 | 7 (24.13) |
| DENV-4 | 1 (3.44) |
| Mixed infections (N = 18) | |
| DENV-1 & -2 | 9 (50) |
| DENV-1 & -4 | 3 (16.66) |
| DENV-2 & -3 | 5 (27.77) |
| DENV-2 & -4 | 1 (5.55) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patro, A.R.K.; Mohanty, S.; Prusty, B.K.; Singh, D.K.; Gaikwad, S.; Saswat, T.; Chattopadhyay, S.; Das, B.K.; Tripathy, R.; Ravindran, B. Cytokine Signature Associated with Disease Severity in Dengue. Viruses 2019, 11, 34. https://doi.org/10.3390/v11010034
Patro ARK, Mohanty S, Prusty BK, Singh DK, Gaikwad S, Saswat T, Chattopadhyay S, Das BK, Tripathy R, Ravindran B. Cytokine Signature Associated with Disease Severity in Dengue. Viruses. 2019; 11(1):34. https://doi.org/10.3390/v11010034
Chicago/Turabian StylePatro, A. Raj Kumar, Sriprasad Mohanty, Birendra K. Prusty, Diwakar K. Singh, Sagar Gaikwad, Tanuja Saswat, Soma Chattopadhyay, Bidyut K. Das, Rina Tripathy, and Balachandran Ravindran. 2019. "Cytokine Signature Associated with Disease Severity in Dengue" Viruses 11, no. 1: 34. https://doi.org/10.3390/v11010034
APA StylePatro, A. R. K., Mohanty, S., Prusty, B. K., Singh, D. K., Gaikwad, S., Saswat, T., Chattopadhyay, S., Das, B. K., Tripathy, R., & Ravindran, B. (2019). Cytokine Signature Associated with Disease Severity in Dengue. Viruses, 11(1), 34. https://doi.org/10.3390/v11010034