An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2


 , ,
, ,  and
and 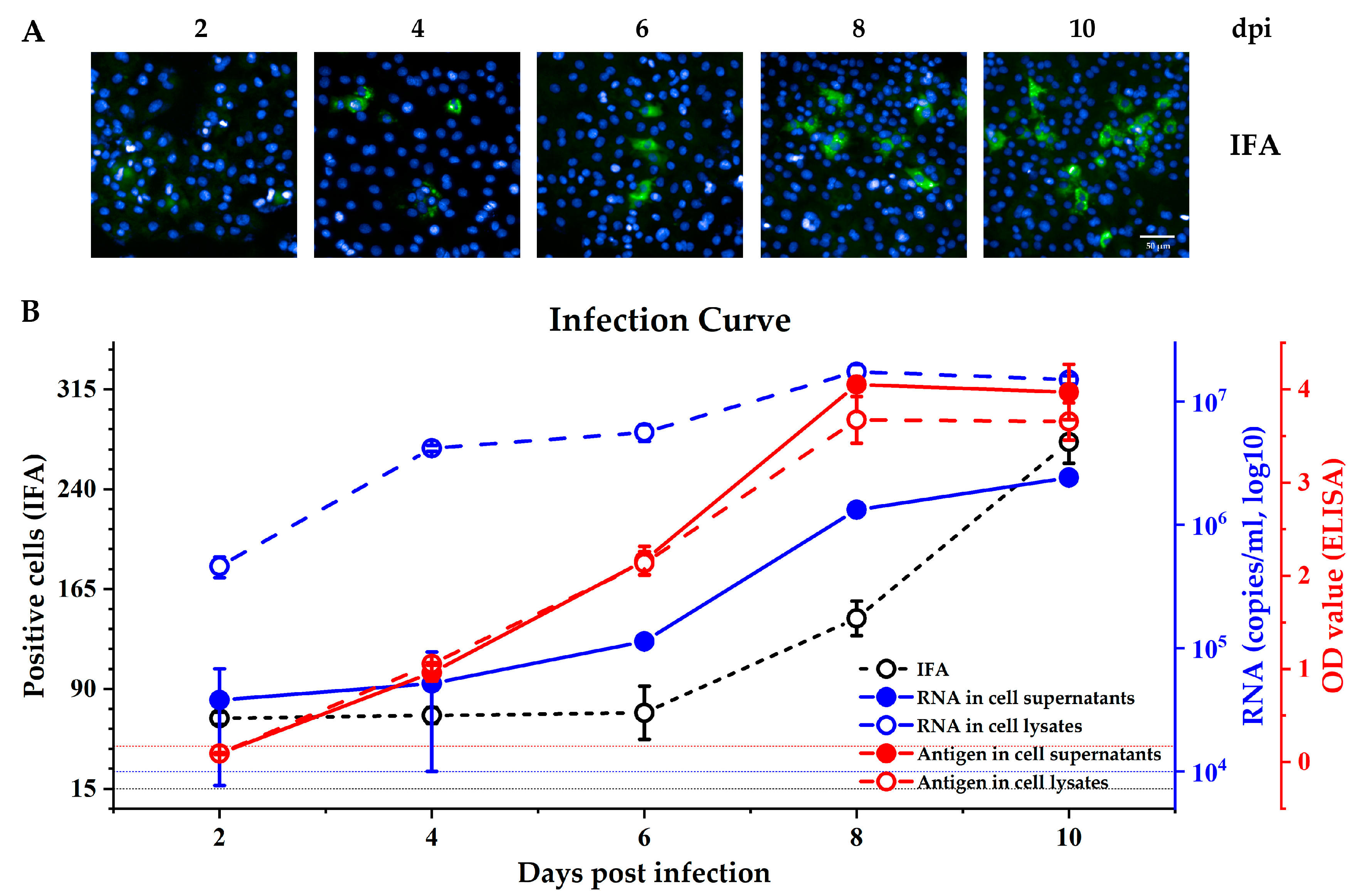{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell lines, Monoclonal Antibodies, and Viruses
2.2. Blood Serum Samples
2.3. Real-Time PCR
2.4. Virus Infection
2.5. Neutralization Assay
2.6. Immunofluorescence Assay
2.7. Detection of HEV Antigen
2.8. Statistical Analysis
3. Results
3.1. Comparing the Different Methods for Measuring Kernow C1-p6 Infection in HepG2/C3A Cells
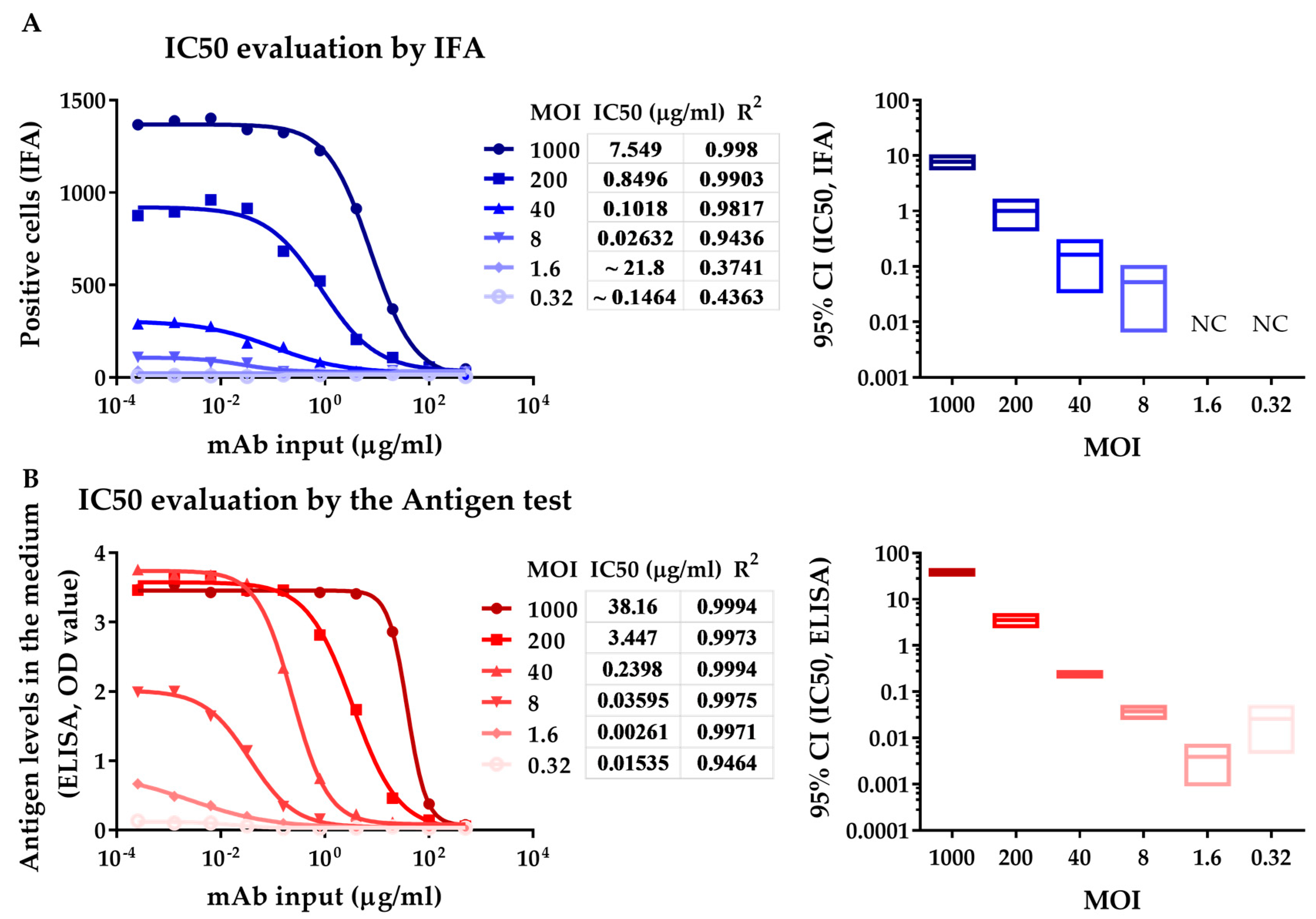
3.2. Neutralization Assay with a HEV-Specific mAb against Kernow C1 p6 Strain Analyzed by IFA and ELISA
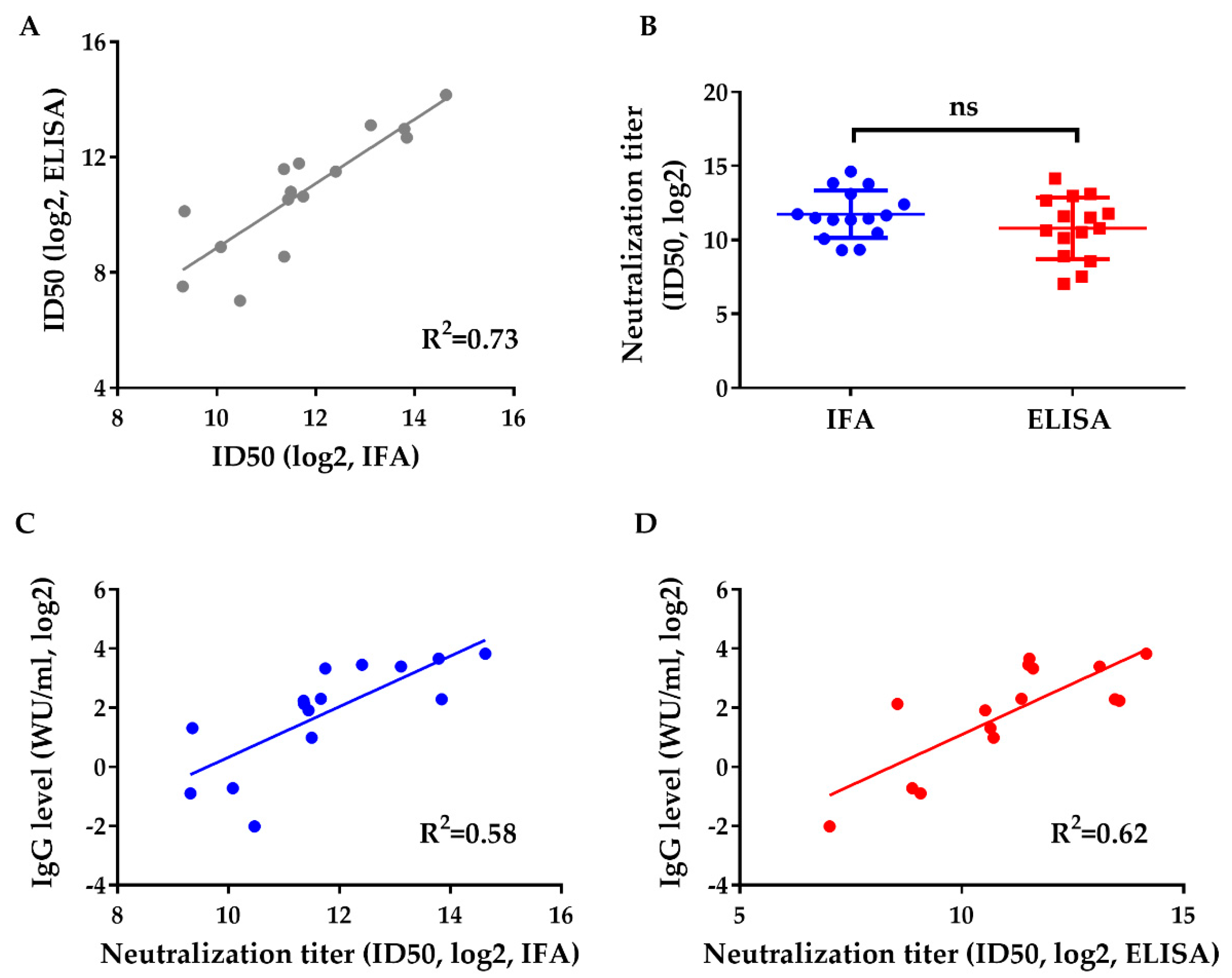
3.3. Neutralization of Human Sera after Gt 1 HEV Vaccine Administration against Gt 3 HEV (Kernow)
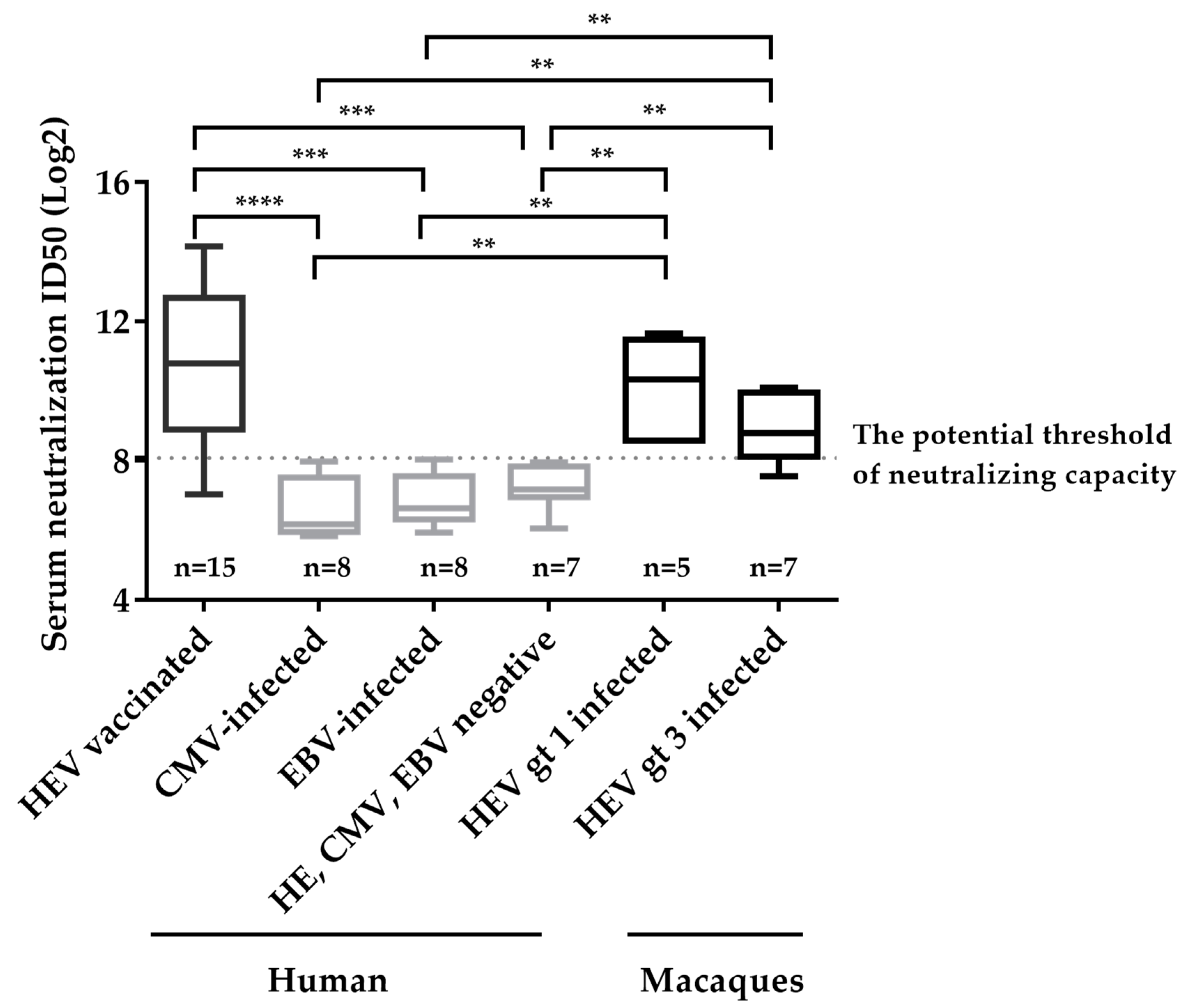
3.4. Determination of the Potential Threshold for Neutralizing Capacity and Specificity of Neutralizing Assay Involving Detection of Secreted pORF2
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis e virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on hepatitis e virology: Implications for clinical practice. J. Hepatol. 2016, 65, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Melgaco, J.G.; Gardinali, N.R.; de Mello, V.D.; Leal, M.; Lewis-Ximenez, L.L.; Pinto, M.A. Hepatitis e: Update on prevention and control. BioMed Res. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Tan, B.H.; Teo, E.C.Y.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic infection with camelid hepatitis e virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology 2016, 150, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.F.; Huang, S.J.; Wu, T.; Hu, Y.M.; Wang, Z.Z.; Wang, H.; Jiang, H.M.; Wang, Y.J.; Yan, Q.; et al. Long-term efficacy of a hepatitis e vaccine. N. Engl. J. Med. 2015, 372, 914–922. [Google Scholar] [CrossRef]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.M.; Walker, C.M.; Xia, N.S.; Zheng, Z.Z.; Feng, Z.D. Origin, antigenicity, and function of a secreted form of orf2 in hepatitis e virus infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent carriage of hepatitis e virus in patients with hiv infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef]
- Tanaka, T.; Takahashi, M.; Takahashi, H.; Ichiyama, K.; Hoshino, Y.; Nagashima, S.; Mizuo, H.; Okamoto, H. Development and characterization of a genotype 4 hepatitis e virus cell culture system using a he-jf5/15f strain recovered from a fulminant hepatitis patient. J. Clin. Microbiol. 2009, 47, 1906–1910. [Google Scholar] [CrossRef]
- Shukla, P.; Nguyen, H.T.; Faulk, K.; Mather, K.; Torian, U.; Engle, R.E.; Emerson, S.U. Adaptation of a genotype 3 hepatitis e virus to efficient growth in cell culture depends on an inserted human gene segment acquired by recombination. J. Virol. 2012, 86, 5697–5707. [Google Scholar] [CrossRef]
- Graff, J.; Nguyen, H.; Yu, C.; Elkins, W.R.; St Claire, M.S.; Purcell, R.H.; Emerson, S.U. The open reading frame 3 gene of hepatitis e virus contains a cis-reactive element and encodes a protein required for infection of macaques. J. Virol. 2005, 79, 6680–6689. [Google Scholar] [CrossRef]
- Yin, X.; Ambardekar, C.; Lu, Y.R.; Feng, Z.D. Distinct entry mechanisms for nonenveloped and quasi-enveloped hepatitis e viruses. J. Virol. 2016, 90, 4232–4242. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Tang, Z.; Wang, S.; Cai, W.; Wen, G.; Ji, W.; Yu, J.; Zhang, K.; Xia, N.; Zheng, Z. The establishment of high-throughput neutralization titer evaluation model for hepatitis e virus (hev). Chin. J. Virol. 2015, 31, 1. [Google Scholar]
- Gu, Y.; Tang, X.H.; Zhang, X.; Song, C.L.; Zheng, M.H.; Wang, K.H.; Zhang, J.; Ng, M.H.; Hew, C.L.; Li, S.W.; et al. Structural basis for the neutralization of hepatitis e virus by a cross-genotype antibody. Cell Res. 2015, 25, 604–620. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Jirintai; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; et al. Hepatitis e virus (hev) strains in serum samples can replicate efficiently in cultured cells despite the coexistence of hev antibodies: Characterization of hev virions in blood circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef]
- Behrendt, P.; Bremer, B.; Todt, D.; Brown, R.J.P.; Heim, A.; Manns, M.P.; Steinmann, E.; Wedemeyer, H. Hepatitis e virus (hev) orf2 antigen levels differentiate between acute and chronic hev infection. J. Infect Dis. 2016, 214, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the orf3 protein of hepatitis e virus (hev) can capture hev particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.C.; Saliou, J.M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F.; et al. Hepatitis e virus lifecycle and identification of 3 forms of the orf2 capsid protein. Gastroenterology 2018, 154, 211–223. [Google Scholar] [CrossRef]
- Zhang, F.; Li, X.H.; Li, Z.; Harrison, T.J.; Chong, H.H.; Qiao, S.; Huang, W.J.; Zhang, H.Y.; Zhuang, H.; Wang, Y.C. Detection of hev antigen as a novel marker for the diagnosis of hepatitis e. J. Med. Virol. 2006, 78, 1441–1448. [Google Scholar] [CrossRef]
- Zhang, J.; Gu, Y.; Ge, S.X.; Li, S.W.; He, Z.Q.; Huang, G.Y.; Zhuang, H.; Ng, M.H.; Xia, N.S. Analysis of hepatitis e virus neutralization sites using monoclonal antibodies directed against a virus capsid protein. Vaccine 2005, 23, 2881–2892. [Google Scholar] [CrossRef]
- Shukla, P.; Nguyen, H.T.; Torian, U.; Engle, R.E.; Faulk, K.; Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Purcell, R.H.; Emerson, S.U. Cross-species infections of cultured cells by hepatitis e virus and discovery of an infectious virus-host recombinant. Proc. Natl. Acad. Sci. USA 2011, 108, 2438–2443. [Google Scholar] [CrossRef]
- Feng, Z.D.; Hensley, L.; McKnight, K.L.; Hu, F.Y.; Madden, V.; Ping, L.F.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.C.; Zhang, J.; Zhang, X.F.; Zhou, C.; Wang, Z.Z.; Huang, S.J.; Wang, H.; Yang, C.L.; Jiang, H.M.; Cai, J.P.; et al. Efficacy and safety of a recombinant hepatitis e vaccine in healthy adults: A large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895–902. [Google Scholar] [CrossRef]
- Zhang, J.; Ge, S.X.; Huang, G.Y.; Li, S.W.; He, Z.Q.; Wang, Y.B.; Zheng, Y.J.; Gu, Y.; Ng, M.H.; Xia, N.S. Evaluation of antibody-based and nucleic acid-based assays for diagnosis of hepatitis e virus infection in a rhesus monkey model. J. Med. Virol. 2003, 71, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time rt-pcr assay for rapid and sensitive detection of hepatitis e virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.P.; Tang, Z.M.; Yang, F.; Zhang, K.; Ji, W.F.; Cai, W.; Huang, S.J.; Wu, T.; Zhang, J.; Zheng, Z.Z.; et al. A valuable antigen detection method for diagnosis of acute hepatitis e. J. Clin. Microbiol. 2015, 53, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis e virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef]
- Wang, C.H.; Flehmig, B.; Moeckli, R. Transmission of hepatitis-e virus by transfusion. Lancet 1993, 341, 825–826. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Cai, W.; Yin, X.; Tang, Z.; Wen, G.; Ambardekar, C.; Li, X.; Ying, D.; Feng, Z.; Zheng, Z.; et al. An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2. Viruses 2019, 11, 64. https://doi.org/10.3390/v11010064
Liu C, Cai W, Yin X, Tang Z, Wen G, Ambardekar C, Li X, Ying D, Feng Z, Zheng Z, et al. An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2. Viruses. 2019; 11(1):64. https://doi.org/10.3390/v11010064
Chicago/Turabian StyleLiu, Chang, Wei Cai, Xin Yin, Zimin Tang, Guiping Wen, Charuta Ambardekar, Xinlei Li, Dong Ying, Zongdi Feng, Zizheng Zheng, and et al. 2019. "An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2" Viruses 11, no. 1: 64. https://doi.org/10.3390/v11010064
APA StyleLiu, C., Cai, W., Yin, X., Tang, Z., Wen, G., Ambardekar, C., Li, X., Ying, D., Feng, Z., Zheng, Z., & Xia, N. (2019). An Optimized High-Throughput Neutralization Assay for Hepatitis E Virus (HEV) Involving Detection of Secreted Porf2. Viruses, 11(1), 64. https://doi.org/10.3390/v11010064