Molecular Surveillance of EHV-1 Strains Circulating in France during and after the Major 2009 Outbreak in Normandy Involving Respiratory Infection, Neurological Disorder, and Abortion


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
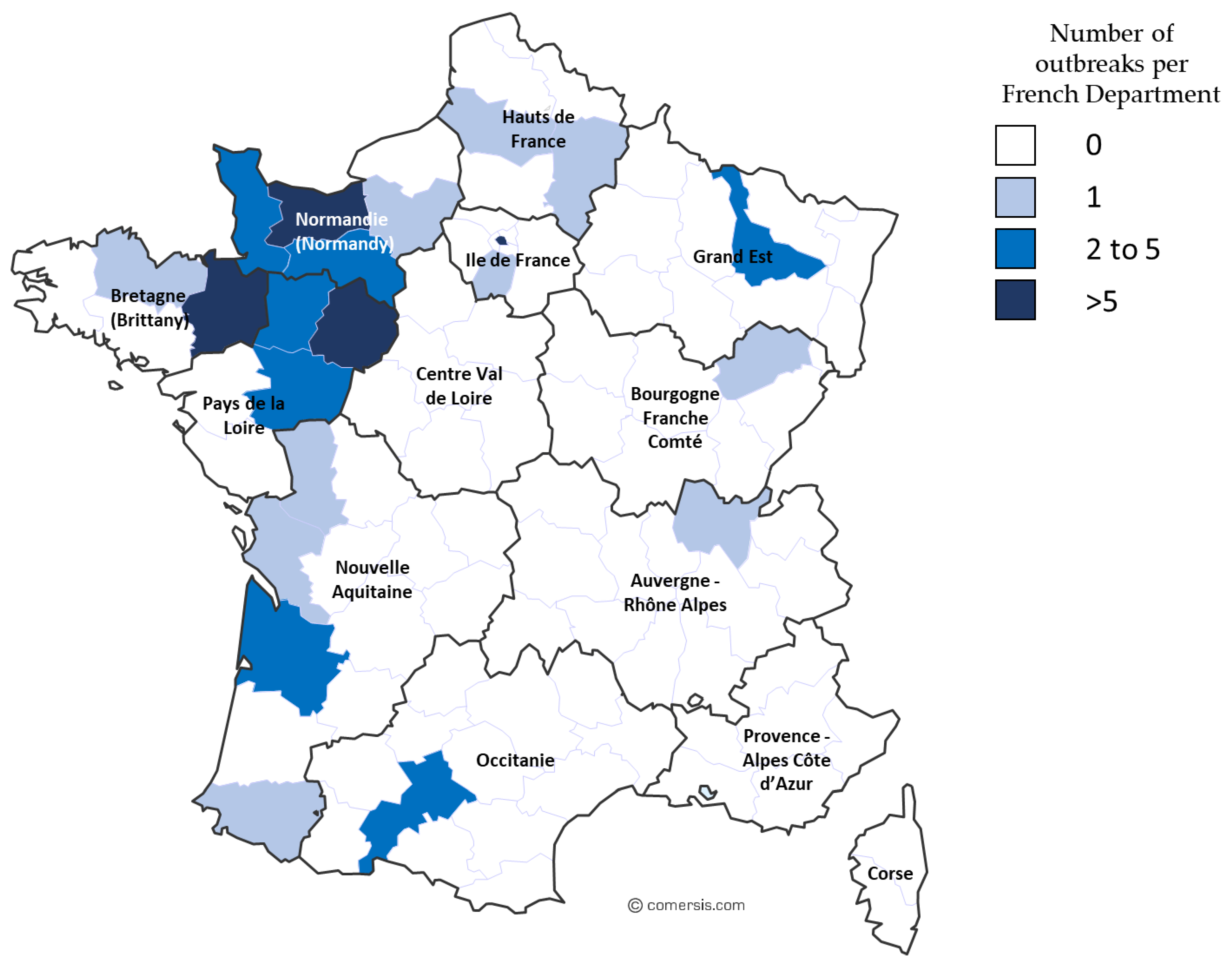
2.1. EHV-1 Outbreak Data Collection
2.2. Biological Samples And Nucleic acids
2.3. EHV-1 Identification And Quantification
2.4. EHV-1 Strain Typing (ORF30 DNA Polymerase, Position 2254)
2.5. ORF30 Sequencing And Analysis
2.6. Multi-locus Sequence Typing (MLST) Analysis
2.7. Statistical Analysis
3. Results
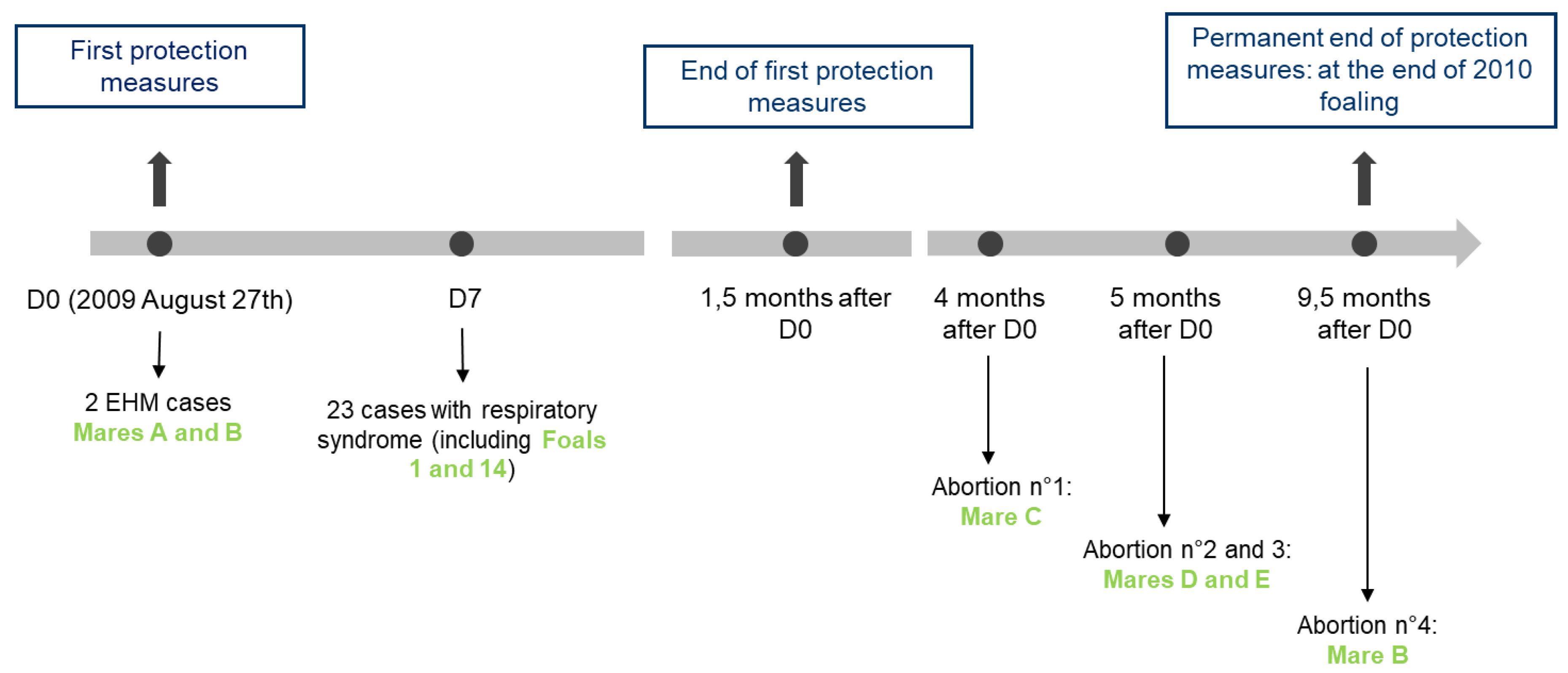
3.1. The 2009 Multi-Syndromic EHV-1 Outbreak
3.1.1. Neurological Cases
3.1.2. Respiratory Infections
3.1.3. Abortion Cases
3.1.4. Biosafety Measures
3.2. Surveillance and Phylogeny from 2009 to 2018
3.2.1. Outbreaks, Forms of Disease, and ORF30 A2254G Typing
3.2.2. ORF30 Sequence Analysis
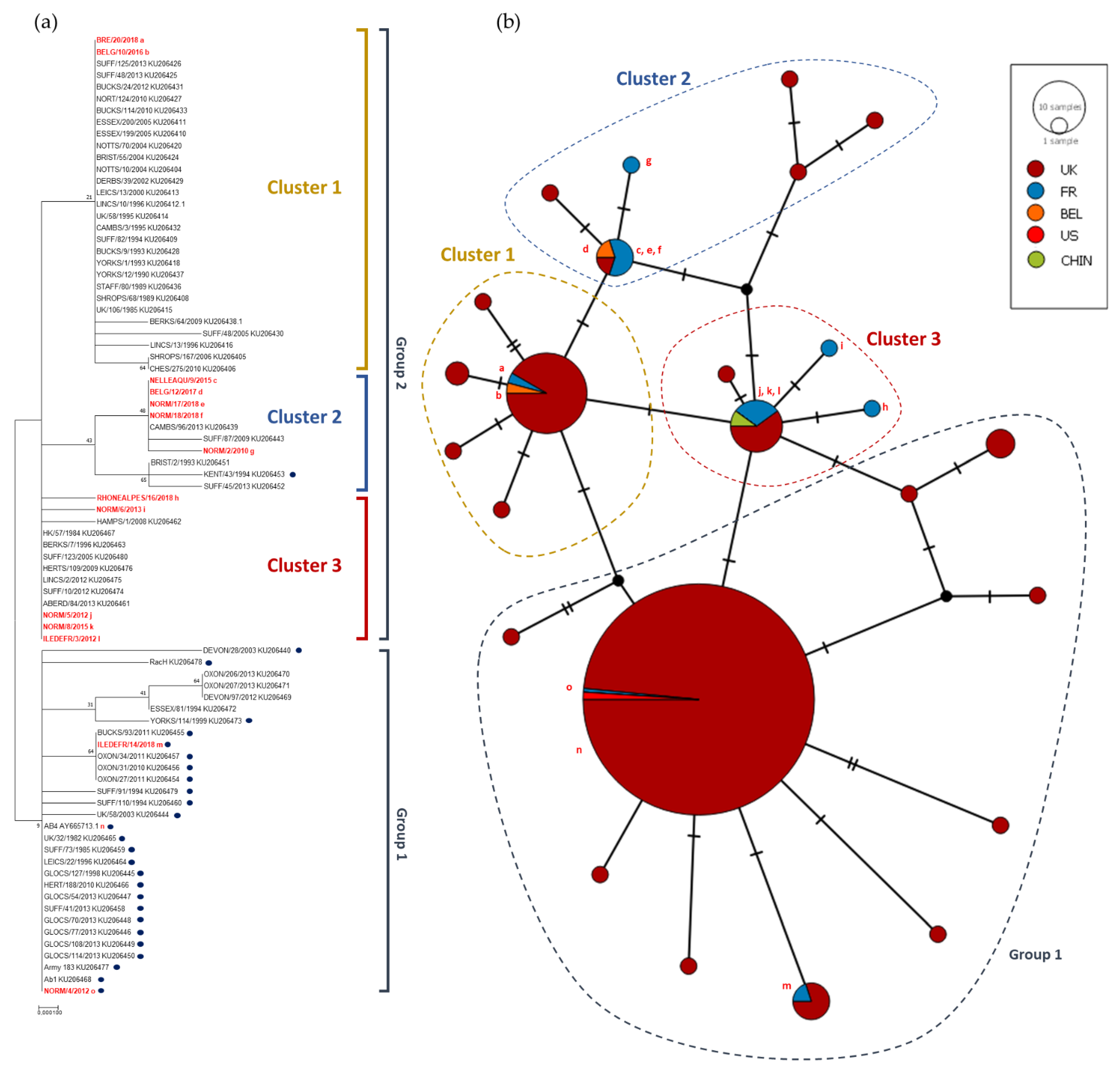
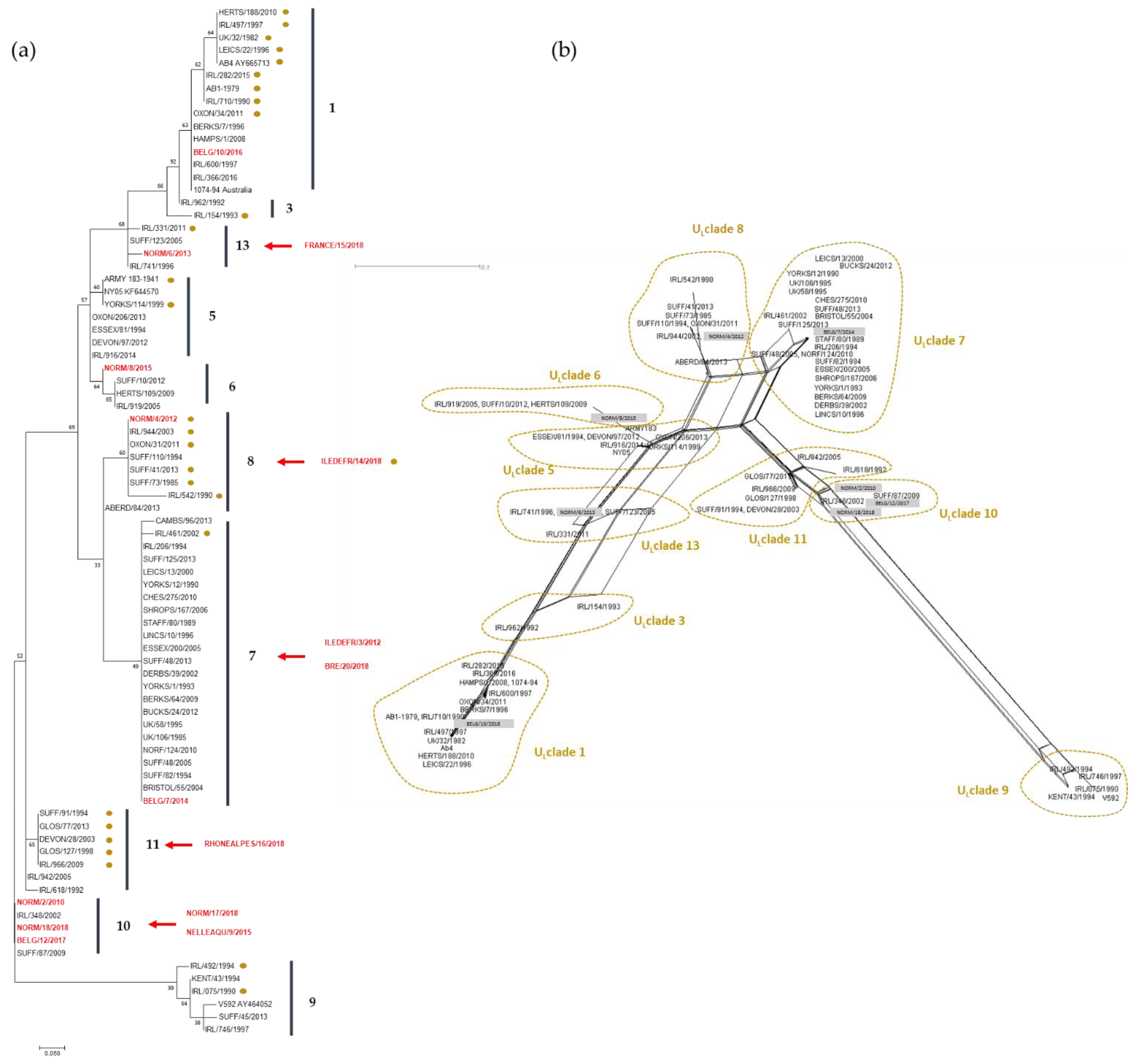
3.2.3. Phylogeny and Multi-locus Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The Order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Abdelgawad, A.; Azab, W.; Damiani, A.M.; Baumgartner, K.; Will, H.; Osterrieder, N.; Greenwood, A.D. Zebra-borne equine herpesvirus type 1 (EHV-1) infection in non-African captive mammals. Vet. Microbiol. 2014, 169, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Wohlsein, P.; Lehmbecker, A.; Spitzbarth, I.; Algermissen, D.; Baumgärtner, W.; Böer, M.; Kummrow, M.; Haas, L.; Grummer, B. Fatal epizootic equine herpesvirus 1 infections in new and unnatural hosts. Vet. Microbiol. 2011, 149, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.I.; Ludwig, H. Molecular biological characterization of equine herpesvirus type 1 (EHV-1) isolates from ruminant hosts. Virus Res. 1988, 11, 127–139. [Google Scholar] [CrossRef]
- Ghanem, Y.M.; Fukushi, H.; Ibrahim, E.S.M.; Ohya, K.; Yamaguchi, T.; Kennedy, M. Molecular phylogeny of equine herpesvirus 1 isolates from onager, zebra and Thomson’s gazelle. Arch. Virol. 2008, 153, 2297–2302. [Google Scholar] [CrossRef]
- Dayaram, A.; Franz, M.; Schattschneider, A.; Damiani, A.M.; Bischofberger, S.; Osterrieder, N.; Greenwood, A.D. Long term stability and infectivity of herpesviruses in water. Sci. Rep. UK 2017, 7. [Google Scholar] [CrossRef]
- Paillot, R.; Case, R.; Ross, J.; Newton, R.; Nugent, J. Equine Herpes Virus-1: Virus, Immunity and Vaccines. Open Vet. Sci. J. 2008, 2, 68–91. [Google Scholar] [CrossRef]
- Lunn, D.P.; Davis-Poynter, N.; Flaminio, M.J.B.F.; Horohov, D.W.; Osterrieder, K.; Pusterla, N.; Townsend, H.G.G. Equine Herpesvirus-1 Consensus Statement. J. Vet. Intern. Med. 2009, 23, 450–461. [Google Scholar] [CrossRef]
- Chesters, P.M.; Allsop, R.; Purewal, A.; Edington, N. Detection of Latency-Associated Transcripts of Equid Herpesvirus 1 in Equine Leukocytes but Not in Trigeminal Ganglia. J. Virol. 1997, 71, 7. [Google Scholar]
- Aleman, M.; Pickles, K.J.; Simonek, G.; Madigan, J.E. Latent Equine Herpesvirus-1 in Trigeminal Ganglia and Equine Idiopathic Headshaking. J. Vet. Intern. Med. 2012, 26, 192–194. [Google Scholar] [CrossRef]
- Rock, D.L. The molecular basis of latent infections by alphaherpesviruses. Semin. Virol. 1993, 4, 157–165. [Google Scholar] [CrossRef]
- Gulati, B.R.; Sharma, H.; Riyesh, T.; Khurana, S.K.; Kapoor, S. Viral and Host Strategies for Regulation of Latency and Reactivation in Equid Herpesviruses. Asian J. Anim. Vet. Adv. 2015, 10, 669–689. [Google Scholar] [CrossRef] [Green Version]
- Telford, E.A.R.; Watson, M.S.; McBride, K.; Davison, A.J. The DNA sequence of equine herpesvirus-1. Virology 1992, 189, 304–316. [Google Scholar] [CrossRef]
- Bryant, N.A.; Wilkie, G.S.; Russell, C.A.; Compston, L.; Grafham, D.; Clissold, L.; McLay, K.; Medcalf, L.; Newton, R.; Davison, A.J.; et al. Genetic diversity of equine herpesvirus 1 isolated from neurological, abortigenic and respiratory disease outbreaks. Transbound. Emerg. Dis. 2018, 65, 817–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuxson, J.L.; Hartley, C.A.; Ficorilli, N.P.; Symes, S.J.; Devlin, J.M.; Gilkerson, J.R. Comparing the genetic diversity of ORF30 of Australian isolates of 3 equid alphaherpesviruses. Vet. Microbiol. 2014, 169, 50–57. [Google Scholar] [CrossRef]
- Crowhurst, F.A.; Dickinson, G.; Burrows, R. An outbreak of paresis in mares and geldings associated with equid herpesvirus 1. Vet. Rec. 1981, 109, 527–528. [Google Scholar]
- Mumford, J.A.; Rossdale, P.D.; Jessett, D.M.; Gann, S.J.; Ousey, J.; Cook, R.F. Serological and virological investigations of an equid herpesvirus 1 (EHV-1) abortion storm on a stud farm in 1985. J. Reprod. Fertil. 1987, 35, 509–518. [Google Scholar]
- Anagha, G.; Gulati, B.R.; Riyesh, T.; Virmani, N. Genetic characterization of equine herpesvirus 1 isolates from abortion outbreaks in India. Arch. Virol. 2017, 162, 157–163. [Google Scholar] [CrossRef]
- Nugent, J.; Birch-Machin, I.; Smith, K.C.; Mumford, J.A.; Swann, Z.; Newton, J.R.; Bowden, R.J.; Allen, G.P.; Davis-Poynter, N. Analysis of Equid Herpesvirus 1 Strain Variation Reveals a Point Mutation of the DNA Polymerase Strongly Associated with Neuropathogenic versus Nonneuropathogenic Disease Outbreaks. J. Virol. 2006, 80, 4047–4060. [Google Scholar] [CrossRef] [Green Version]
- Goodman, L.B.; Loregian, A.; Perkins, G.A.; Nugent, J.; Buckles, E.L.; Mercorelli, B.; Kydd, J.H.; Palù, G.; Smith, K.C.; Osterrieder, N.; et al. A Point Mutation in a Herpesvirus Polymerase Determines Neuropathogenicity. PLoS Pathog. 2007, 3, e160. [Google Scholar] [CrossRef]
- Pronost, S.; Léon, A.; Legrand, L.; Fortier, C.; Miszczak, F.; Freymuth, F.; Fortier, G. Neuropathogenic and non-neuropathogenic variants of equine herpesvirus 1 in France. Vet. Microbiol. 2010, 145, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.P. Development of a Real-Time Polymerase Chain Reaction Assay for Rapid Diagnosis of Neuropathogenic Strains of Equine Herpesvirus-1. J. Vet. Diagn. Investig. 2007, 19, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Van de Walle, G.R.; Goupil, R.; Wishon, C.; Damiani, A.; Perkins, G.A.; Osterrieder, N. A Single-Nucleotide Polymorphism in a Herpesvirus DNA Polymerase Is Sufficient to Cause Lethal Neurological Disease. J. Infect. Dis. 2009, 200, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.; Goodman, L.; Van de Walle, G.; Osterrieder, N.; Greenwood, A. A Point Mutation in a Herpesvirus Co-Determines Neuropathogenicity and Viral Shedding. Viruses 2017, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Gryspeerdt, A.; Vandekerckhove, A.; Van Doorsselaere, J.; Van de Walle, G.; Nauwynck, H. Description of an unusually large outbreak of nervous system disorders caused by equine herpesvirus 1 (EHV1) in 2009 in Belgium. Vlaams Diergeneeskundig Tijdschrif 2011, 80, 147–153. [Google Scholar]
- Garvey, M.; Lyons, R.; Hector, R.; Walsh, C.; Arkins, S.; Cullinane, A. Molecular Characterisation of Equine Herpesvirus 1 Isolates from Cases of Abortion, Respiratory and Neurological Disease in Ireland between 1990 and 2017. Pathogens 2019, 8, 7. [Google Scholar] [CrossRef]
- Pronost, S.; Legrand, L.; Pitel, P.-H.; Wegge, B.; Lissens, J.; Freymuth, F.; Richard, E.; Fortier, G. Outbreak of Equine Herpesvirus Myeloencephalopathy in France: A Clinical and Molecular Investigation: Outbreak of EHV-1 myeloencephalopathy. Transbound. Emerg. Dis. 2012, 59, 256–263. [Google Scholar] [CrossRef]
- Diallo, I.S.; Hewitson, G.; Wright, L.; Rodwell, B.J.; Corney, B.G. Detection of equine herpesvirus type 1 using a real-time polymerase chain reaction. J. Virol. Methods 2006, 131, 92–98. [Google Scholar] [CrossRef]
- Thieulent, C.J.; Hue, E.S.; Fortier, C.I.; Dallemagne, P.; Zientara, S.; Munier-Lehmann, H.; Hans, A.; Fortier, G.D.; Pitel, P.-H.; Vidalain, P.-O.; et al. Screening and evaluation of antiviral compounds against Equid alpha-herpesviruses using an impedance-based cellular assay. Virology 2019, 526, 105–116. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- CodonCode Aligner. Available online: https://www.codoncode.com (accessed on 27 September 2019).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- PopART. Available online: http://popart.otago.ac.nz (accessed on 27 September 2019).
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Bryant, D. Neighbor-Net: An Agglomerative Method for the Construction of Phylogenetic Networks. Mol. Biol. Evol. 2003, 21, 255–265. [Google Scholar] [CrossRef]
- Walter, J.; Seeh, C.; Fey, K.; Bleul, U.; Osterrieder, N. Clinical observations and management of a severe equine herpesvirus type 1 outbreak with abortion and encephalomyelitis. Acta Vet. Scand. 2013, 55, 19. [Google Scholar] [CrossRef]
- Vissani, M.A.; Becerra, M.L.; Olguín Perglione, C.; Tordoya, M.S.; Miño, S.; Barrandeguy, M. Neuropathogenic and non-neuropathogenic genotypes of Equid Herpesvirus type 1 in Argentina. Vet. Microbiol. 2009, 139, 361–364. [Google Scholar] [CrossRef]
- Pronost, S.; Cook, R.F.; Fortier, G.; Timoney, P.J.; Balasuriya, U.B.R. Relationship between equine herpesvirus-1 myeloencephalopathy and viral genotype: EHV-1, genotype and EHM. Equine Vet. J. 2010, 42, 672–674. [Google Scholar] [CrossRef]
- Lechmann, J.; Schoster, A.; Ernstberger, M.; Fouché, N.; Fraefel, C.; Bachofen, C. A novel PCR protocol for detection and differentiation of neuropathogenic and non-neuropathogenic equid alphaherpesvirus 1. J. Vet. Diagn. Investig. 2019, 31, 696–703. [Google Scholar] [CrossRef]
- Liu, S.; Knafels, J.D.; Chang, J.S.; Waszak, G.A.; Baldwin, E.T.; Deibel, M.R.; Thomsen, D.R.; Homa, F.L.; Wells, P.A.; Tory, M.C.; et al. Crystal Structure of the Herpes Simplex Virus 1 DNA Polymerase. J. Biol. Chem. 2006, 281, 18193–18200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrouk, K.; Piret, J.; Boivin, G. Herpesvirus DNA polymerases: Structures, functions and inhibitors. Vir. Res. 2017, 234, 177–192. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Respiratory | Abortion | Neurological | ND 1 | Total |
|---|---|---|---|---|---|
| 2012 | MD 2 | MD | MD | MD | MD |
| 2013 | 1 | 1 | 3 | 0 | 5 |
| 2014 | 14 | 10 | 3 | 0 | 27 |
| 2015 | 11 | 16 | 1 | 0 | 28 |
| 2016 | 11 | 8 | 7 | 3 | 32 |
| 2017 | 5 | 9 | 6 | 3 | 27 |
| 2018 | 15 | 16 | 5 | 20 | 56 |
| Mares | Foals | ||||||
|---|---|---|---|---|---|---|---|
| Group | Total Number | Respiratory Disease | Abortion 1 | EHM | Total Number | Sex | Respiratory Disease |
| 1 | 13 | 0 | 3 1 | 2 | 13 2 | Female | 6 |
| 2 | 9 | 0 | 1 | 0 | 9 2 | Male | 4 |
| 3 | 10 | 0 | 0 | 0 | 10 3 | Female | 0 |
| 4 | 12 | 4 | 2 1 | 0 | 12 3 | Male | 9 |
| 5 | 6 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6 | 10 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| D7 | D14 | D21 | D28 | D35 | D42 | D49 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Horse | NS | Blood | NS | Blood | NS | Blood | NS | Blood | NS | Blood | NS | Blood | NS | Blood | |
| Group 1 and Group 2 | 1 | 2.14E + 08 | nd | 6.75E + 05 | 1.05E + 03 | 7.04E + 02 | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| 2 | 3.02E + 08 | nd | 4.52E + 04 | NEG | 7.04E + 02 | NEG | 6.05E + 03 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 3 | 9.29E + 07 | nd | NEG | NEG | NEG | NEG | 8.09E + 02 | NEG | 2.14E + 03 | 4.58E + 02 | 1.11E + 05 | NEG | NEG | NEG | |
| 4 | 1.91E + 06 | nd | 1.81E + 05 | 2.98E + 03 | 7.34E + 04 | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG | |
| 5 | 1.66E + 06 | nd | NEG | NEG | 3.67E + 04 | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | |
| 6 | 1.08E + 07 | nd | 1.30E + 04 | NEG | 1.23E + 03 | NEG | 6.05E + 03 | NEG | POS | NEG | NEG | NEG | NEG | NEG | |
| 7 | 2.23E + 05 | nd | NEG | POS | POS | NEG | NEG | POS | NEG | POS | NEG | NEG | NEG | NEG | |
| 8 | 1.28E + 05 | nd | 3.93E + 04 | POS | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 9 | 4.84E + 04 | nd | NEG | NEG | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 10 | NEG | nd | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 11 | NEG | nd | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 12 | POS | nd | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 13 | NEG | nd | NEG | NEG | NEG | NEG | NEG | NEG | NEG | 5.64E + 02 | NEG | NEG | NEG | NEG | |
| 14 | NEG | nd | 6.75E + 05 | NEG | 1.31E + 08 | NEG | POS | NEG | 6.48E + 03 | NEG | NEG | NEG | NEG | NEG | |
| 15 | nd | nd | nd | nd | POS | NEG | POS | NEG | NEG | NEG | POS | NEG | NEG | NEG | |
| Group 4 | 16 | 2.26E + 04 | nd | NEG | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG |
| 17 | 1.18E + 06 | nd | 4.27E + 03 | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| 18 | 1.97E + 04 | nd | POS | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | NEG | |
| Respiratory Syndrome | Abortion | Neurological Syndrome | ND 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Strains | A2254 | G2254 | A2254 | G2254 | A2254 | G2254 | A2254 | G2254 | No A/G Typing | Outbreaks (RESPE) |
| 1 major outbreak 2009 | 30 | 24 | 0 | 2 | 2 | 0 | 2 | 0 | 0 | 0 | / |
| 2012 | 8 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 5 * | Missing data |
| 2013 | 19 | 0 | 0 | 6 | 7 | 2 | 1 | 0 | 1 | 2 * | 4 |
| 2014 | 10 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 2 * | 26 |
| 2015 | 32 | 0 | 0 | 17 | 0 | 1 | 1 | 1 | 0 | 12 * | 27 |
| 2016 | 18 | 1 | 0 | 8 | 0 | 2 | 4 | 0 | 1 | 2 * | 32 |
| 2017 | 22 | 1 | 0 | 2 | 1 | 0 | 7 | 1 | 2 | 8 * | 23 |
| 2018 | 71 | 9 | 4 | 17 | 1 | 10 | 2 | 5 | 11 | 12 * | 56 |
| Type | Respiratory | Abortion | Neurological | Information Missing | Total | |
|---|---|---|---|---|---|---|
| A2254 | Non-neuro. | 11 (73%) | 58 (84%) | 15 (48%) | 7 (32%) | 91(66%) |
| G2254 | Neuro. | 4 (27%) | 11 (16%) | 16 (52%) | 15 (68%) | 46 (34%) |
| Chi square p-value | Resp./Ab. | 0.325642 | ||||
| Resp./Neu. | 0.109608 | |||||
| Ab./Neu. | 0.000202 * | |||||
| All three | 0.000996 * |
| ORF | UL/MLST Clades | 2 | 5 | 8 | 11 | 11 | 11 | 13 | 13 | 13 | 13 | 13 | 13 | 14 | 14 | 14 | 15 | 22 | 29 | 30 | 30 | 31 | 32 | 33 | 33 | 34 | 36 | 37 | 39 | 40 | 42 | 45 | 46 | 50 | 52 | 57 | 73 | 76 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LOCI | 59 | 114 | 114 | 189 | 235 | 250 | 305 | 405 | 460 | 492 | 493 | 499 | 618/20 | 628 | 692 | 166 | 430 | 12 | 752 | 990 | 90 | 42 | 15 | 976 | 66 | 47 | 265 | 440 | 196 | 1275 | 427 | 140 | 367 | 386 | 804 | 122 | 128 | |
| Ab4 AY665713 | 1 | G | G | D | Q | R | A | S | A | A | E | T | A | --- | R | S | D | S | T | D | E | N | S | N | N | D | S | A | S | R | K | E | F | P | A | K | A | F |
| NORM/2/2010 | 10 | D | V | D | Q | R | A | L | A | T | E | T | A | PSR | R | S | N | S | T | N | K | S | S | H | D | D | S | V | S | R | K | G | S | P | V | R | A | S |
| ILEDEFR/3/2012 | 7 | Q | M | A | S | A | T | E | T | A | N | E | V | V | R | S | ||||||||||||||||||||||
| NORM/4/2012 | 8 | D | G | D | Q | R | A | S | A | T | E | T | T | PSR | K | N | N | S | T | D | E | N | S | H | D | D | S | V | S | R | K | G | S | P | V | K | A | S |
| NORM/6/2013 | 13 | D | G | D | Q | R | A | S | A | A | E | T | A | PSR | R | S | D | S | T | N | E | N | S | N | D | D | S | V | S | R | K | G | S | P | A | K | A | S |
| BELG/7/2014 | 7 | D | G | D | Q | M | A | S | A | T | E | T | A | PSR | K | N | N | S | T | N | E | S | S | H | D | D | S | V | S | R | K | G | S | P | V | R | A | S |
| NORM/8/2015 | 6 | D | G | D | Q | R | S * | S | T | A | E | T | A | PSR | R | S | N | S | T | N | E | N | S | H | D | D | S | V | S | R | K | G | S | P | V | K | A | S |
| NELLEAQU/9/2015 | 10 | Q | R | A | L | A | T | E | T | A | N | K | V | V | R | S | ||||||||||||||||||||||
| BELG/10/2016 | 1 | D | G | D | Q | R | A | S | A | A | E | T | A | --- | R | S | D | S | T | N | E | N | S | N | N | D | S | A | S | R | K | E | F | P | A | K | A | F |
| BELG/12/2017 | 10 | D | V | D | Q | R | A | L | A | T | E | T | A | PSR | R | S | N | S | T | N | K | S | S | H | D | D | S | V | S | R | K | G | S | P | V | R | A | S |
| ILEDEFR/14/2018 | 8 | Q | R | A | S | A | T | E | T | T | D | E | V | V | K | S | ||||||||||||||||||||||
| FRANCE/15/2018 | 13 | Q | R | A | S | A | A | E | T | A | N | E | V | A | K | S | ||||||||||||||||||||||
| RHONEALPES/16/2018 | 11 | Q | R | A | L | A | T | E | T | A | N | E | V | V | R | S | ||||||||||||||||||||||
| NORM/17/2018 | 10 | Q | R | A | L | A | T | E | T | A | N | K | V | V | R | S | ||||||||||||||||||||||
| NORM/18/2018 | 10 | D | V | D | Q | R | A | L | A | T | E | T | A | PSR | R | S | N | S | T | N | K | S | S | H | D | D | S | V | S | R | K | G | S | P | V | R | A | S |
| BRE/20/2018 | 7 | Q | M | A | S | A | T | E | T | A | N | E | V | V | R | S | ||||||||||||||||||||||
| V592 AY464052 | 9 | D | V | N | K | R | A | L | A | T | E | T | A | PSR | R | S | N | P | K | N | K | S | L | H | D | G | R | V | L | H | R | G | S | S | V | R | V | S |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutton, G.; Garvey, M.; Cullinane, A.; Jourdan, M.; Fortier, C.; Moreau, P.; Foursin, M.; Gryspeerdt, A.; Maisonnier, V.; Marcillaud-Pitel, C.; et al. Molecular Surveillance of EHV-1 Strains Circulating in France during and after the Major 2009 Outbreak in Normandy Involving Respiratory Infection, Neurological Disorder, and Abortion. Viruses 2019, 11, 916. https://doi.org/10.3390/v11100916
Sutton G, Garvey M, Cullinane A, Jourdan M, Fortier C, Moreau P, Foursin M, Gryspeerdt A, Maisonnier V, Marcillaud-Pitel C, et al. Molecular Surveillance of EHV-1 Strains Circulating in France during and after the Major 2009 Outbreak in Normandy Involving Respiratory Infection, Neurological Disorder, and Abortion. Viruses. 2019; 11(10):916. https://doi.org/10.3390/v11100916
Chicago/Turabian StyleSutton, Gabrielle, Marie Garvey, Ann Cullinane, Marion Jourdan, Christine Fortier, Peggy Moreau, Marc Foursin, Annick Gryspeerdt, Virginie Maisonnier, Christel Marcillaud-Pitel, and et al. 2019. "Molecular Surveillance of EHV-1 Strains Circulating in France during and after the Major 2009 Outbreak in Normandy Involving Respiratory Infection, Neurological Disorder, and Abortion" Viruses 11, no. 10: 916. https://doi.org/10.3390/v11100916
APA StyleSutton, G., Garvey, M., Cullinane, A., Jourdan, M., Fortier, C., Moreau, P., Foursin, M., Gryspeerdt, A., Maisonnier, V., Marcillaud-Pitel, C., Legrand, L., Paillot, R., & Pronost, S. (2019). Molecular Surveillance of EHV-1 Strains Circulating in France during and after the Major 2009 Outbreak in Normandy Involving Respiratory Infection, Neurological Disorder, and Abortion. Viruses, 11(10), 916. https://doi.org/10.3390/v11100916