Viruses in Horses with Neurologic and Respiratory Diseases

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Samples
2.2. Viral Metagenomics
2.3. Generation of Full Genomes of Novel Horse Parvovirus
2.4. Real-Time PCR
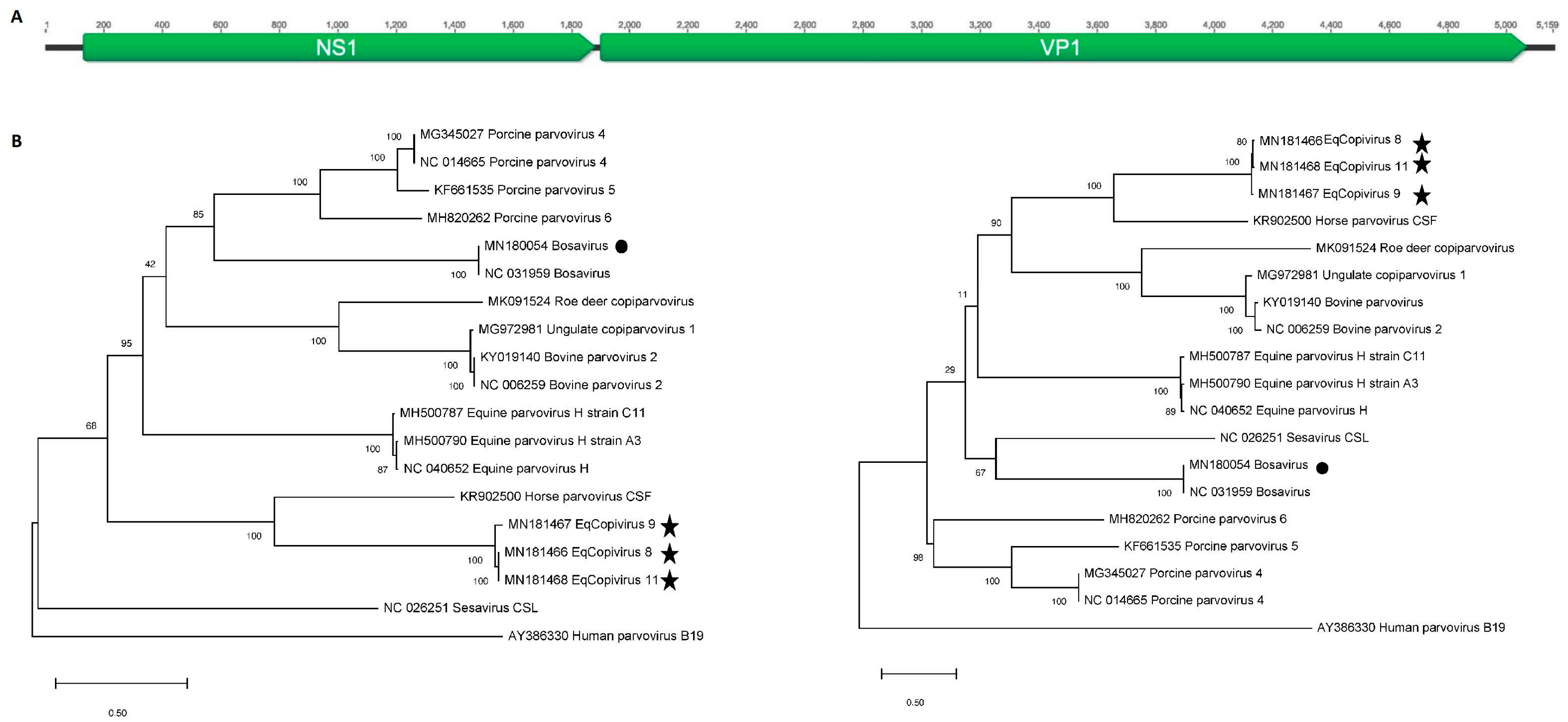
2.5. Phylogenetic Analysis
3. Results
3.1. Viral Metagenomics
3.2. Generation of Near-Full Length Genomes of Novel Equine Copiparvovirus (Eqcopivirus)
3.3. Real-Time PCR Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Horse Population Report. Food and Agriculture Organization (FAO) of the United Nations. Available online: http://faostat.fao.org/fastat/serviet (accessed on 28 August 2019).
- Divers, T.J.; Tennant, B.C.; Kumar, A.; McDonough, S.; Cullen, J.; Bhuva, N.; Jain, K.; Chauhan, L.S.; Scheel, T.K.H.; Lipkin, W.I. , et al. New parvovirus associated with serum hepatitis in horses after inoculation of common biological product. Emerg. Infect. Dis 2018, 24, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Gilkerson, J.R.; Bailey, K.E.; Diaz-Mendez, A.; Hartley, C.A. Update on viral diseases of the equine respiratory tract. Vet. Clin. North. Am. Equine Pract 2015, 31, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Manuja, A.; Gulati, B.R.; Virmani, N.; Tripathi, B.N. Zoonotic viral diseases of equines and their impact on human and animal health. Open Virol. J. 2018, 12, 80–98. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D. Hendra virus. Vet. Clin. North. Am. Equine Pract. 2014, 30, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Vin, R.; Leutenegger, C.M.; Mittel, L.D.; Divers, T.J. Enteric coronavirus infection in adult horses. Vet. J. 2018, 231, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Timoney, P.J.; McCollum, W.H. Equine viral arteritis. Vet. Clin. North. Am. Equine Pract 1993, 9, 295–309. [Google Scholar] [CrossRef]
- Nowotny, N.; Kolodziejek, J.; Jehle, C.O.; Suchy, A.; Staeheli, P.; Schwemmle, M. Isolation and characterization of a new subtype of borna disease virus. J. Virol. 2000, 74, 5655–5658. [Google Scholar] [CrossRef] [PubMed]
- Chapman, G.E.; Baylis, M.; Archer, D.; Daly, J.M. The challenges posed by equine arboviruses. Equine Vet. J. 2018, 50, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Chandriani, S.; Skewes-Cox, P.; Zhong, W.; Ganem, D.E.; Divers, T.J.; Van Blaricum, A.J.; Tennant, B.C.; Kistler, A.L. Identification of a previously undescribed divergent virus from the flaviviridae family in an outbreak of equine serum hepatitis. Proc. Nat. Acad. Sci. USA 2013, 110, E1407–E1415. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Tennant, B.C.; Struzyna, A.; Mrad, D.; Browne, N.; Whelchel, D.; Johnson, P.J.; Jamieson, C.; Lohr, C.V.; Bildfell, R.; et al. Viral testing of 10 cases of theiler’s disease and 37 in-contact horses in the absence of equine biologic product administration: A prospective study (2014-2018). J. Vet. Intern. Med. 2019, 33, 258–265. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Kapoor, A.; Kumar, A.; Tennant, B.C.; Laverack, M.A.; Beard, L.; Delph, K.; Davis, E.; Schott Ii, H.; Lascola, K.; et al. Viral testing of 18 consecutive cases of equine serum hepatitis: A prospective study (2014-2018). J. Vet. Intern. Med. 2019, 33, 251–257. [Google Scholar] [CrossRef]
- Lu, G.; Sun, L.; Ou, J.; Xu, H.; Wu, L.; Li, S. Identification and genetic characterization of a novel parvovirus associated with serum hepatitis in horses in china. Emerg. Microbes. Infect. 2018, 7, 170. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Simmonds, P.; Gerold, G.; Qaisar, N.; Jain, K.; Henriquez, J.A.; Firth, C.; Hirschberg, D.L.; Rice, C.M.; Shields, S.; et al. Characterization of a canine homolog of hepatitis c virus. Proc. Nat. Acad. Sci. USA 2011, 108, 11608–11613. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.; Kapoor, A.; Schneider, B.S.; Wolfe, N.D.; Culshaw, G.; Corcoran, B.; Durham, A.E.; Burden, F.; McGorum, B.C.; Simmonds, P. Viraemic frequencies and seroprevalence of non-primate hepacivirus and equine pegiviruses in horses and other mammalian species. J. Gen. Virol. 2014, 95, 1701–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, S.; Kapoor, A.; Sharp, C.; Schneider, B.S.; Wolfe, N.D.; Culshaw, G.; Corcoran, B.; McGorum, B.C.; Simmonds, P. Nonprimate hepaciviruses in domestic horses, united kingdom. Emerg. Infect. Dis. 2012, 18, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Dubovi, E.J.; Simmonds, P.; Medina, J.L.; Henriquez, J.A.; Mishra, N.; Wagner, J.; Tokarz, R.; Cullen, J.M.; Iadarola, M.J.; et al. Serology-enabled discovery of genetically diverse hepaciviruses in a new host. J. Virol. 2012, 86, 6171–6178. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.H.; Quan, P.L.; Briese, T.; Street, C.; Jabado, O.; Conlan, S.; Ali Khan, S.; Verdugo, D.; Hossain, M.J.; Hutchison, S.K.; et al. Identification of gbv-d, a novel gb-like flavivirus from old world frugivorous bats (pteropus giganteus) in bangladesh. PLoS Path. 2010, 6, e1000972. [Google Scholar] [CrossRef]
- Kapoor, A.; Simmonds, P.; Cullen, J.M.; Scheel, T.K.; Medina, J.L.; Giannitti, F.; Nishiuchi, E.; Brock, K.V.; Burbelo, P.D.; Rice, C.M.; et al. Identification of a pegivirus (gb virus-like virus) that infects horses. J. Virol. 2013, 87, 7185–7190. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Giannitti, F.; Low, J.; Keyes, C.; Ullmann, L.S.; Deng, X.; Aleman, M.; Pesavento, P.A.; Pusterla, N.; Delwart, E. Exploring the virome of diseased horses. J. Gen. Virol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Paim, W.P.; Weber, M.N.; Cibulski, S.P.; da Silva, M.S.; Puhl, D.E.; Budaszewski, R.F.; Varela, A.P.M.; Mayer, F.Q.; Canal, C.W. Characterization of the viral genomes present in commercial batches of horse serum obtained by high-throughput sequencing. Biologicals 2019. [Google Scholar] [CrossRef]
- Victoria, J.G.; Kapoor, A.; Li, L.; Blinkova, O.; Slikas, B.; Wang, C.; Naeem, A.; Zaidi, S.; Delwart, E. Metagenomic analyses of viruses in stool samples from children with acute flaccid paralysis. J. Virol. 2009, 83, 4642–4651. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Deng, X.; Mee, E.T.; Collot-Teixeira, S.; Anderson, R.; Schepelmann, S.; Minor, P.D.; Delwart, E. Comparing viral metagenomics methods using a highly multiplexed human viral pathogens reagent. J. Virol. Methods 2015, 213, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Naccache, S.N.; Ng, T.; Federman, S.; Li, L.; Chiu, C.Y.; Delwart, E.L. An ensemble strategy that significantly improves de novo assembly of microbial genomes from metagenomic next-generation sequencing data. Nucl. Acids Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Kapusinszky, B.; Yugo, D.M.; Phan, T.G.; Deng, X.; Kanevsky, I.; Opriessnig, T.; Woolums, A.R.; Hurley, D.J.; Meng, X.J.; et al. Virome of us bovine calf serum. Biologicals 2017, 46, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Tattersall, P. Parvoviruses: Small does not mean simple. An. Rev. Virol 2014, 1, 517–537. [Google Scholar] [CrossRef]
- Mietzsch, M.; Penzes, J.J.; Agbandje-McKenna, M. Twenty-five years of structural parvovirology. Viruses 2019, 11, 362. [Google Scholar] [CrossRef]
- Zadori, Z.; Szelei, J.; Lacoste, M.C.; Li, Y.; Gariepy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A viral phospholipase a2 is required for parvovirus infectivity. Develop. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef]
- Zhang, L.; Dailey, P.J.; He, T.; Gettie, A.; Bonhoeffer, S.; Perelson, A.S.; Ho, D.D. Rapid clearance of simian immunodeficiency virus particles from plasma of rhesus macaques. J. Virol 1999, 73, 855–860. [Google Scholar]
- Ramratnam, B.; Bonhoeffer, S.; Binley, J.; Hurley, A.; Zhang, L.; Mittler, J.E.; Markowitz, M.; Moore, J.P.; Perelson, A.S.; Ho, D.D. Rapid production and clearance of hiv-1 and hepatitis c virus assessed by large volume plasma apheresis. Lancet 1999, 354, 1782–1785. [Google Scholar] [CrossRef]
- Zhang, L.; Dailey, P.J.; Gettie, A.; Blanchard, J.; Ho, D.D. The liver is a major organ for clearing simian immunodeficiency virus in rhesus monkeys. J. Virol. 2002, 76, 5271–5273. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Penzes, J.J.; et al. Ictv virus taxonomy profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Disease and Sample Types | Total Reads | Equine Hepacivirus | Equine Pegivirus D | Equid Gamma Herpesvirus2 | Equid Gammaherpes Virus5 | Horse Parvovirus-CSF | Novel Parvovirus: Eqcopivirus | Bosavirus |
|---|---|---|---|---|---|---|---|---|
| Neuro Pool1 Plasma | 2,575,048 | 1023 | 17,058 | |||||
| Neuro Pool4 CSF | 1,106,824 | |||||||
| Neuro Pool2 Plasma | 1,612,796 | 7 | ||||||
| Neuro Pool5 CSF | 1,028,398 | |||||||
| Neuro Pool3 Plasma | 2,080,966 | |||||||
| Neuro Pool6 CSF | 1,057,992 | |||||||
| Respiratory Pool7 Plasma | 1,509,792 | 317 | 36,069 | |||||
| Respiratory Pool10 Swab | 622,376 | 1719 | ||||||
| Respiratory Pool8 Plasma | 603,470 | 1087 | 16,355 | |||||
| Respiratory Pool11 Swab | 949,260 | 120 | 25 | 880 | ||||
| Respiratory Pool9 Plasma | 1,520,736 | |||||||
| Respiratory Pool12 Swab | 629,274 | 395 |
| Sample Type | Pool | Equine Hepacivirus | EqPV-H | Eqcopivirus | Horse Parvovirus-CSF | Bosavirus | |
|---|---|---|---|---|---|---|---|
| Neurological Signs | Plasma 1 | Pool1 | 30.32 | ||||
| Plasma 2 | |||||||
| Plasma 3 | |||||||
| Plasma 4 | 27.7 | 20 | |||||
| Plasma 5 | Pool2 | ||||||
| Plasma 6 | 34.65 | 36.73 | |||||
| Plasma 7 | 23.6 | ||||||
| Plasma 8 | |||||||
| Plasma 9 | Pool3 | ||||||
| Plasma 10 | 33.82 | ||||||
| Plasma 11 | |||||||
| Plasma 12 | |||||||
| Plasma 13 | |||||||
| CSF 1 | Pool4 | ||||||
| CSF 2 | |||||||
| CSF 3 | |||||||
| CSF 4 | |||||||
| CSF 5 | Pool5 | ||||||
| CSF 6 | |||||||
| CSF 7 | |||||||
| CSF 8 | |||||||
| CSF 9 | Pool6 | 30 | |||||
| CSF 10 | |||||||
| CSF 11 | |||||||
| CSF 12 | 36.5 | ||||||
| CSF 13 | |||||||
| Respiratory Signs | Plasma 1 | Pool7 | 37.78 | ||||
| Plasma 2 | |||||||
| Plasma 3 | |||||||
| Plasma 4 | 34 | ||||||
| Plasma 5 | 27.53 | ||||||
| Plasma 6 | Pool8 | ||||||
| Plasma 7 | 30.16 | ||||||
| Plasma 8 | |||||||
| Plasma 9 | 34.6 | 34.01 | 21 | ||||
| Plasma 10 | Pool9 | ||||||
| Plasma 11 | |||||||
| Plasma 12 | |||||||
| Plasma 13 | |||||||
| Plasma 14 | |||||||
| Swab 1 | Pool10 | 26.85 | |||||
| Swab 2 | 25.65 | ||||||
| Swab 3 | 25.28 | 26.4 | |||||
| Swab 4 | 36.7 | 26.82 | |||||
| Swab 6 | Pool11 | 25.02 | |||||
| Swab 7 | 32.62 | 27.75 | |||||
| Swab 8 | 25.57 | ||||||
| Swab 9 | 26.24 | ||||||
| Swab 10 | Pool12 | 21.2 | 24.92 | ||||
| Swab 11 | 23.1 | 23.37 | |||||
| Swab 12 | 26.63 | ||||||
| Swab 13 | 32.41 | 23.29 | |||||
| Swab 14 | 37.2 | 26 | 25.3 | ||||
| Healthy Control Group | Plasma 1 | ||||||
| Plasma 2 | 16.8 | ||||||
| Plasma 3 | 21.8 | 37.3 | |||||
| Plasma 4 | 26.5 | ||||||
| Plasma 5 | 36.5 | ||||||
| Plasma 6 | |||||||
| Plasma 7 | |||||||
| Plasma 8 | |||||||
| Plasma 9 | |||||||
| Plasma 10 | |||||||
| Plasma 11 | 28.08 | ||||||
| Plasma 12 | |||||||
| Plasma 13 | |||||||
| Plasma 14 | 30.21 | ||||||
| Plasma 15 | |||||||
| Plasma 16 | |||||||
| Plasma 17 | 26.08 | ||||||
| Plasma 18 | |||||||
| Plasma 19 | 23.02 | 36.9 | |||||
| Plasma 20 | |||||||
| Plasma 21 | |||||||
| Plasma 22 | |||||||
| Plasma 23 | 23.53 | ||||||
| Plasma 24 | |||||||
| Plasma 25 | 25.11 | ||||||
| Plasma 26 | |||||||
| Plasma 27 | 33.83 | ||||||
| Plasma 28 | |||||||
| Plasma 29 | |||||||
| Plasma 30 | |||||||
| Plasma 31 | 33.99 | 30.09 | |||||
| Plasma 32 | 37.93 | 26.34 | 30.1 | ||||
| Plasma 33 | |||||||
| Plasma 34 | |||||||
| Plasma 35 | |||||||
| Plasma 36 | 33.22 | ||||||
| Plasma 37 | |||||||
| Plasma 38 | |||||||
| Plasma 39 | |||||||
| Plasma 40 | 34.69 | 28.63 | |||||
| Plasma 41 | 30.12 |
| Neuro Plasma n = 13 | Neuro CSF n = 13 | Respiratory Plasma n = 14 | Respiratory Swab n = 13 | Healthy Control Plasma n = 41 | Total Samples n = 94 | P Value = Plasma Neurological Versus Healthy | P Value = Plasma Respiratory Versus Healthy | |
|---|---|---|---|---|---|---|---|---|
| Equine flavivirus | 3 (23%) | 0 | 0 | 2 (15%) | 6 (15%) | 11 (12%) | 0.32 | 0.67 |
| EqPV-H | 2 (15%) | 0 | 2 (15%) | 0 | 7 (17%) | 11 (12%) | 1 | 1 |
| Eqcopivirus | 1 (7%) | 1 (7%) | 4 (30%) | 3 (23%) | 7 (17%) | 15 (16%) | 0.443 | 0.663 |
| Horse parvovirus-CSF | 1 (7%) | 1 (7%) | 1 (7%) | 3 (23%) | 2 (5%) | 8 (8.5%) | 1 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altan, E.; Li, Y.; Sabino-Santos Jr, G.; Sawaswong, V.; Barnum, S.; Pusterla, N.; Deng, X.; Delwart, E. Viruses in Horses with Neurologic and Respiratory Diseases. Viruses 2019, 11, 942. https://doi.org/10.3390/v11100942
Altan E, Li Y, Sabino-Santos Jr G, Sawaswong V, Barnum S, Pusterla N, Deng X, Delwart E. Viruses in Horses with Neurologic and Respiratory Diseases. Viruses. 2019; 11(10):942. https://doi.org/10.3390/v11100942
Chicago/Turabian StyleAltan, Eda, Yanpeng Li, Gilberto Sabino-Santos Jr, Vorthon Sawaswong, Samantha Barnum, Nicola Pusterla, Xutao Deng, and Eric Delwart. 2019. "Viruses in Horses with Neurologic and Respiratory Diseases" Viruses 11, no. 10: 942. https://doi.org/10.3390/v11100942
APA StyleAltan, E., Li, Y., Sabino-Santos Jr, G., Sawaswong, V., Barnum, S., Pusterla, N., Deng, X., & Delwart, E. (2019). Viruses in Horses with Neurologic and Respiratory Diseases. Viruses, 11(10), 942. https://doi.org/10.3390/v11100942