Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons



Abstract
:1. Introduction
2. Materials and Methods
2.1. Neuronal Cultures
2.2. ZIKV Infection and Plaque Assay
2.3. Indirect-Immunofluorescence Microscopy
2.4. NanoString nCounter® Gene Expression
2.5. qRT-PCR
2.6. Measurement of Cytokines and Chemokines
2.7. Ingenuity Pathways Analysis (IPA)
3. Results
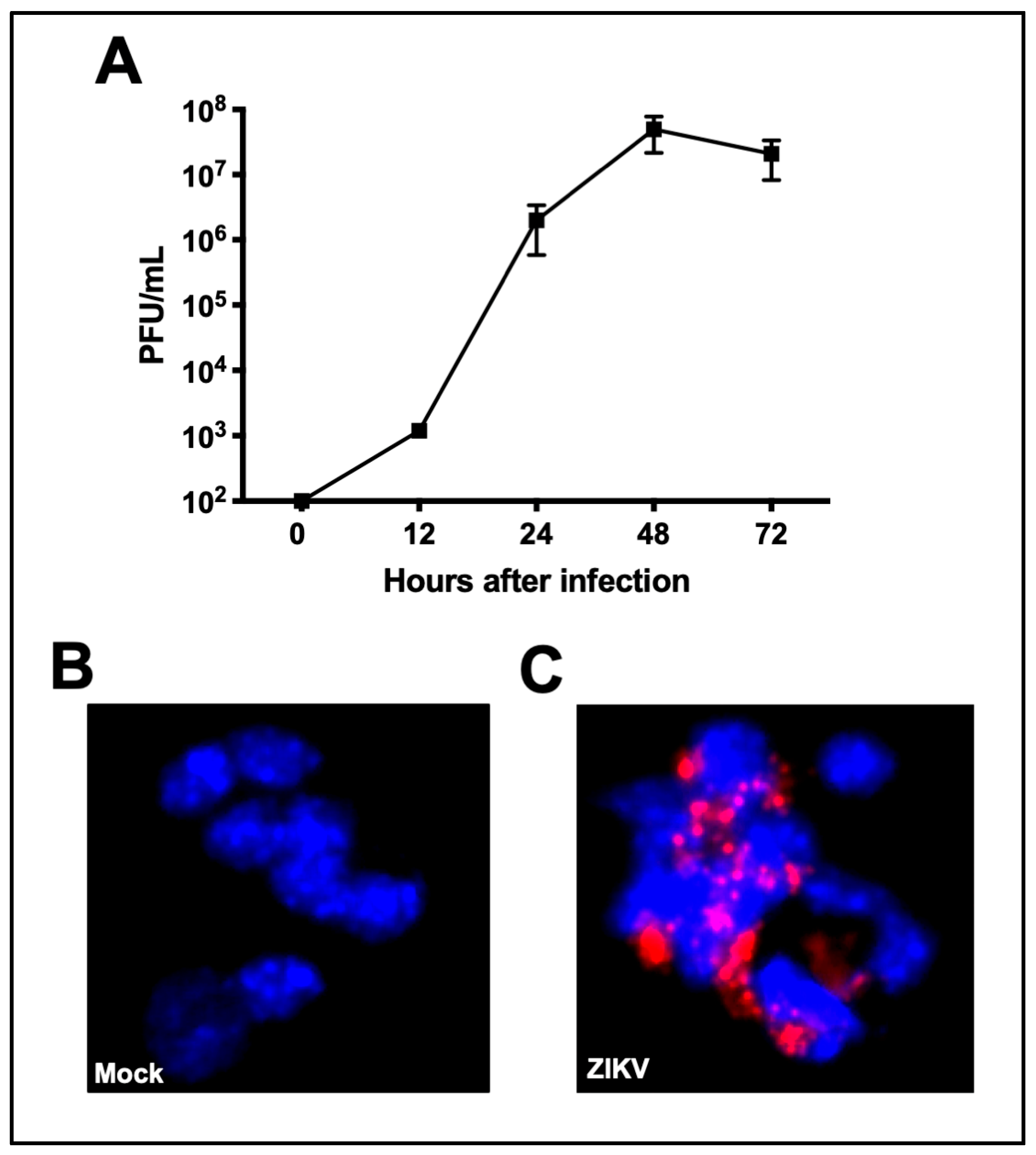
3.1. ZIKV Can Infect Primary Mouse Cortical Neurons
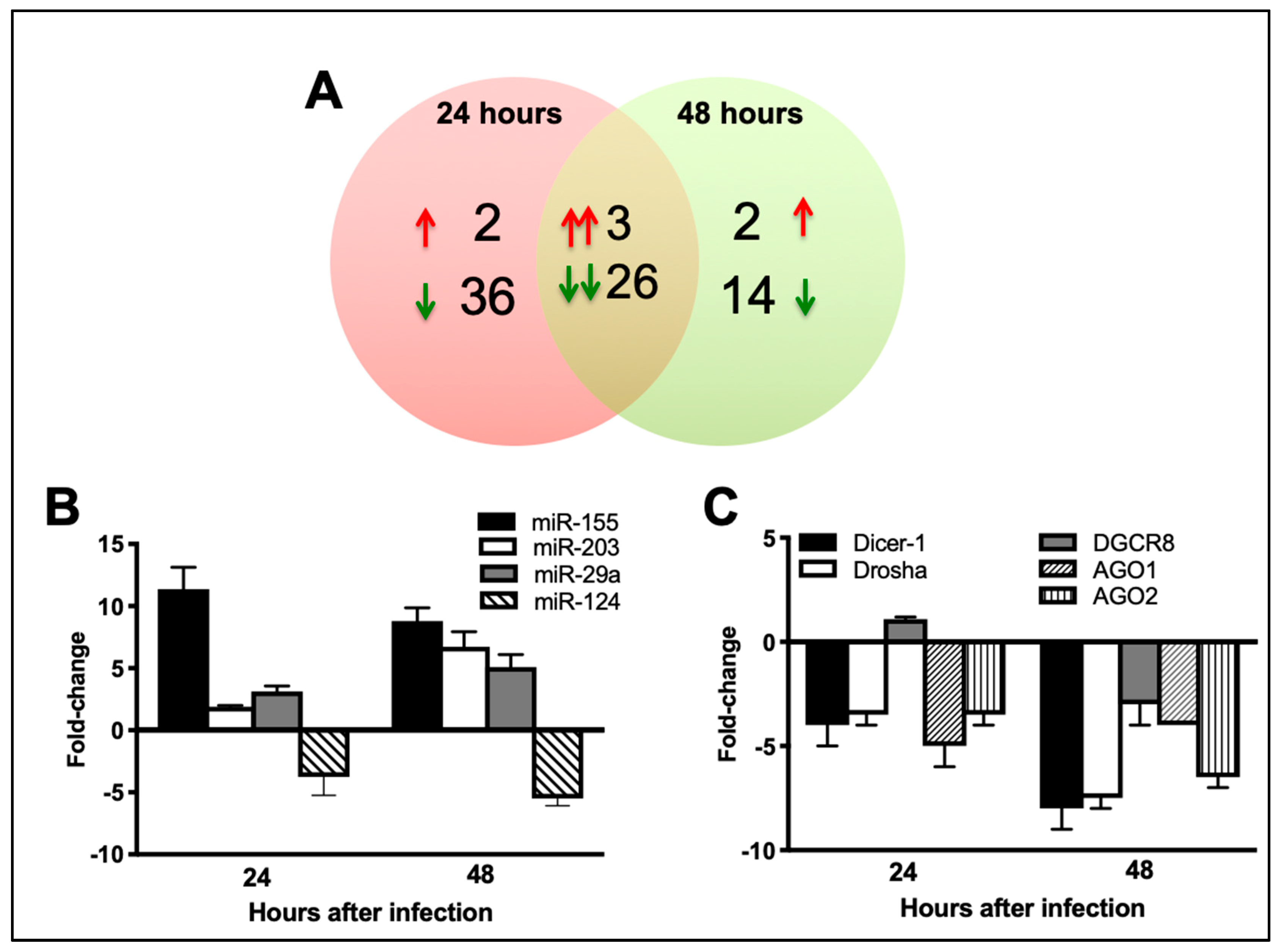
3.2. ZIKV Infection Modulates Cellular miRNA Expression
3.3. ZIKV Infection Results in Downregulation of miRNA Processing Enzymes
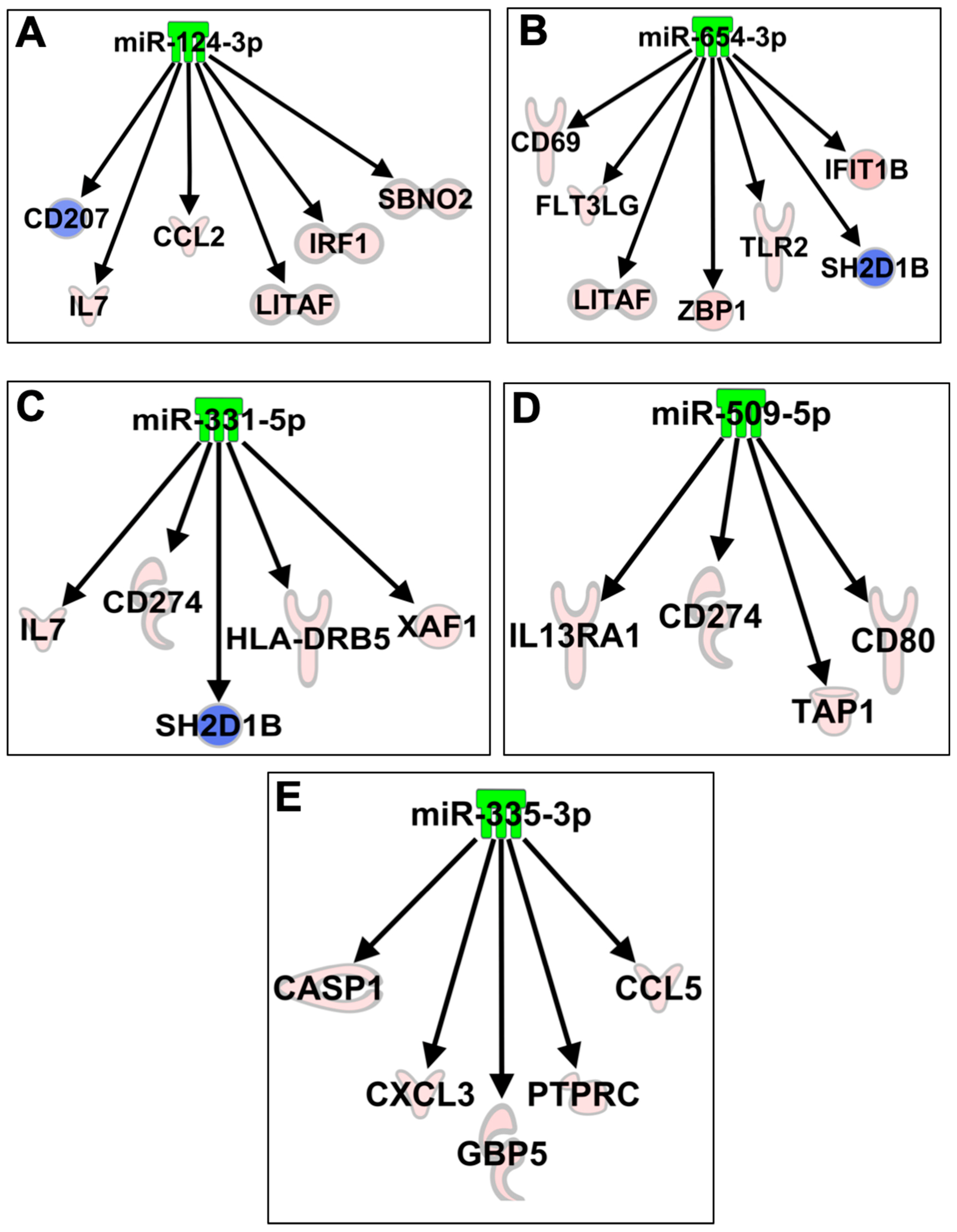
3.4. Functional Analysis of ZIKV-Modulated miRNAs and Their Predicted Targets
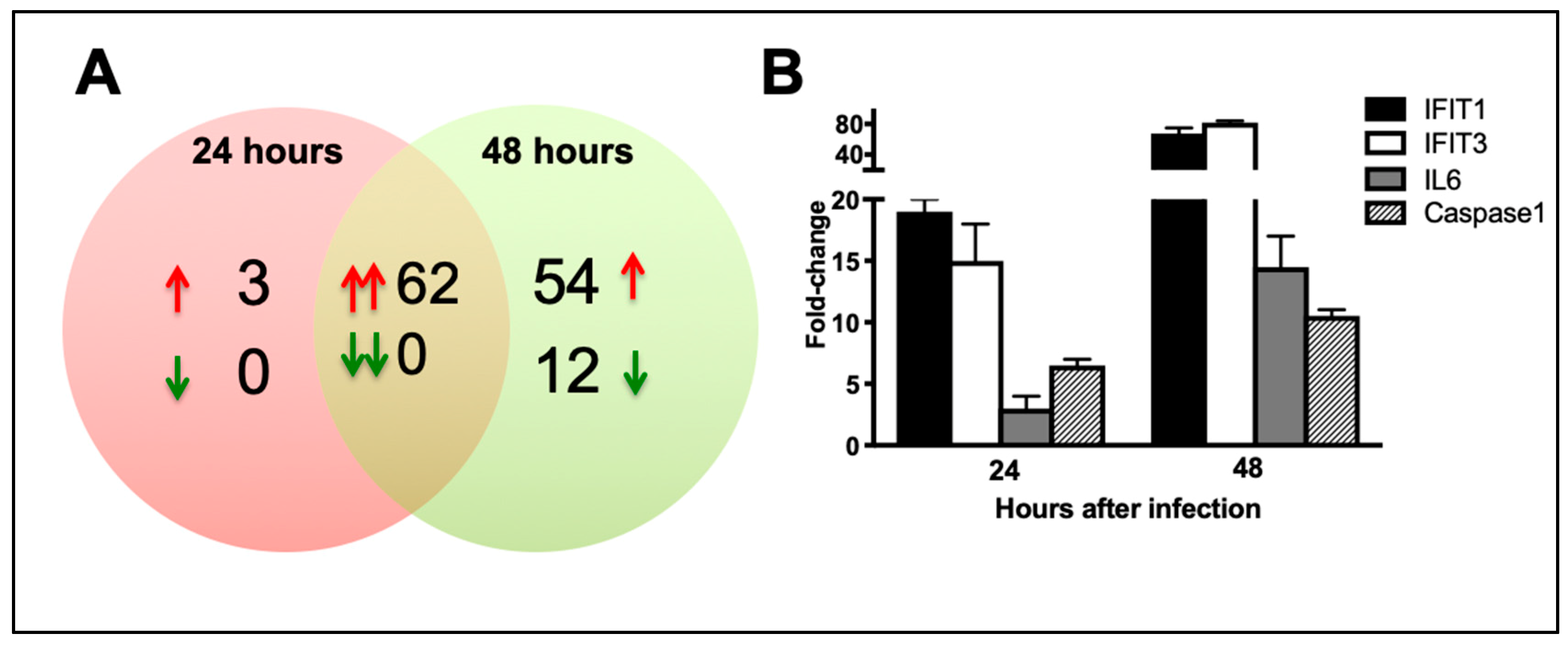
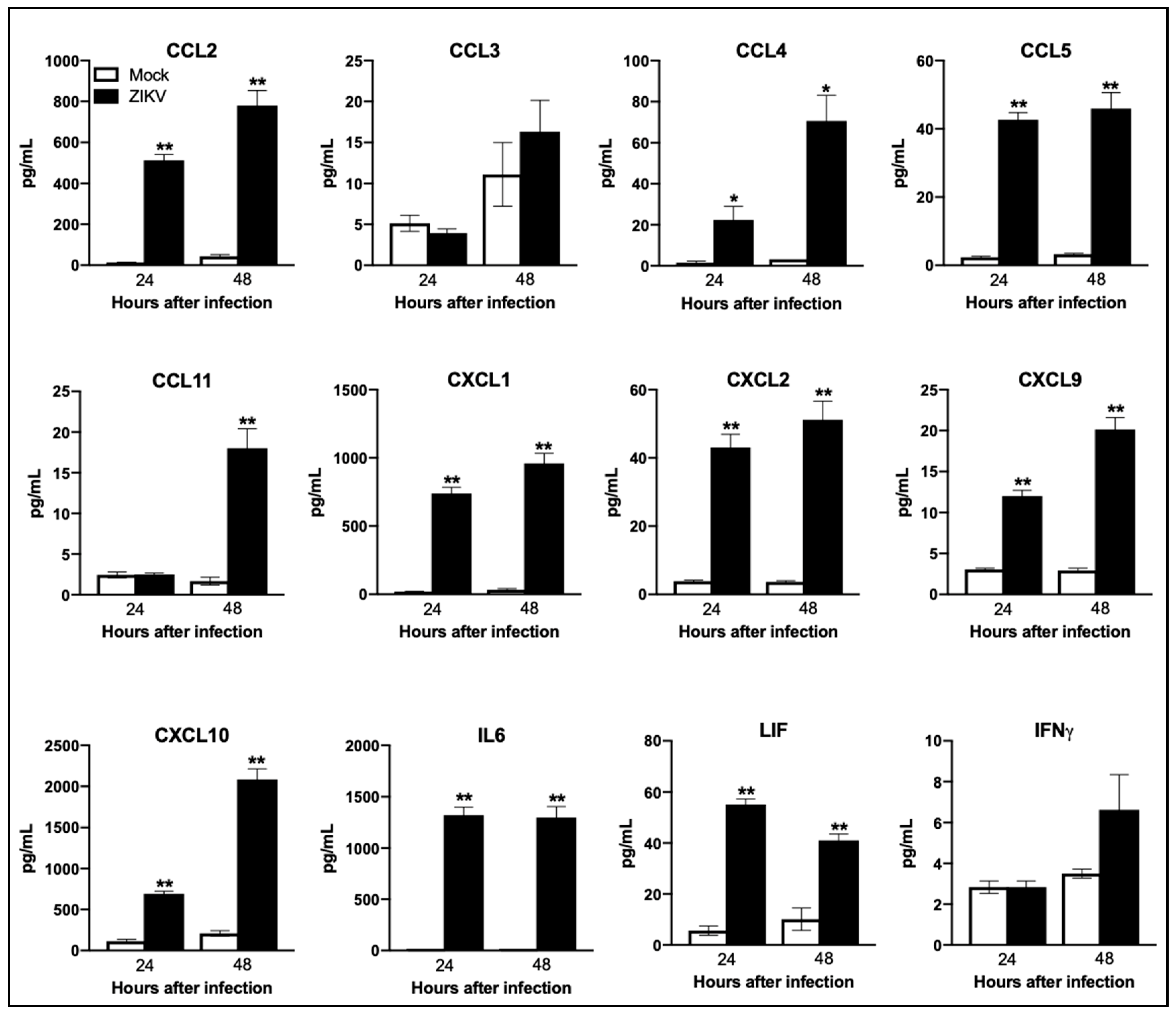
3.5. ZIKV Infection Induces Dramatic Upregulation of Anti-Viral, Inflammatory, and Apoptotic Genes in Neurons
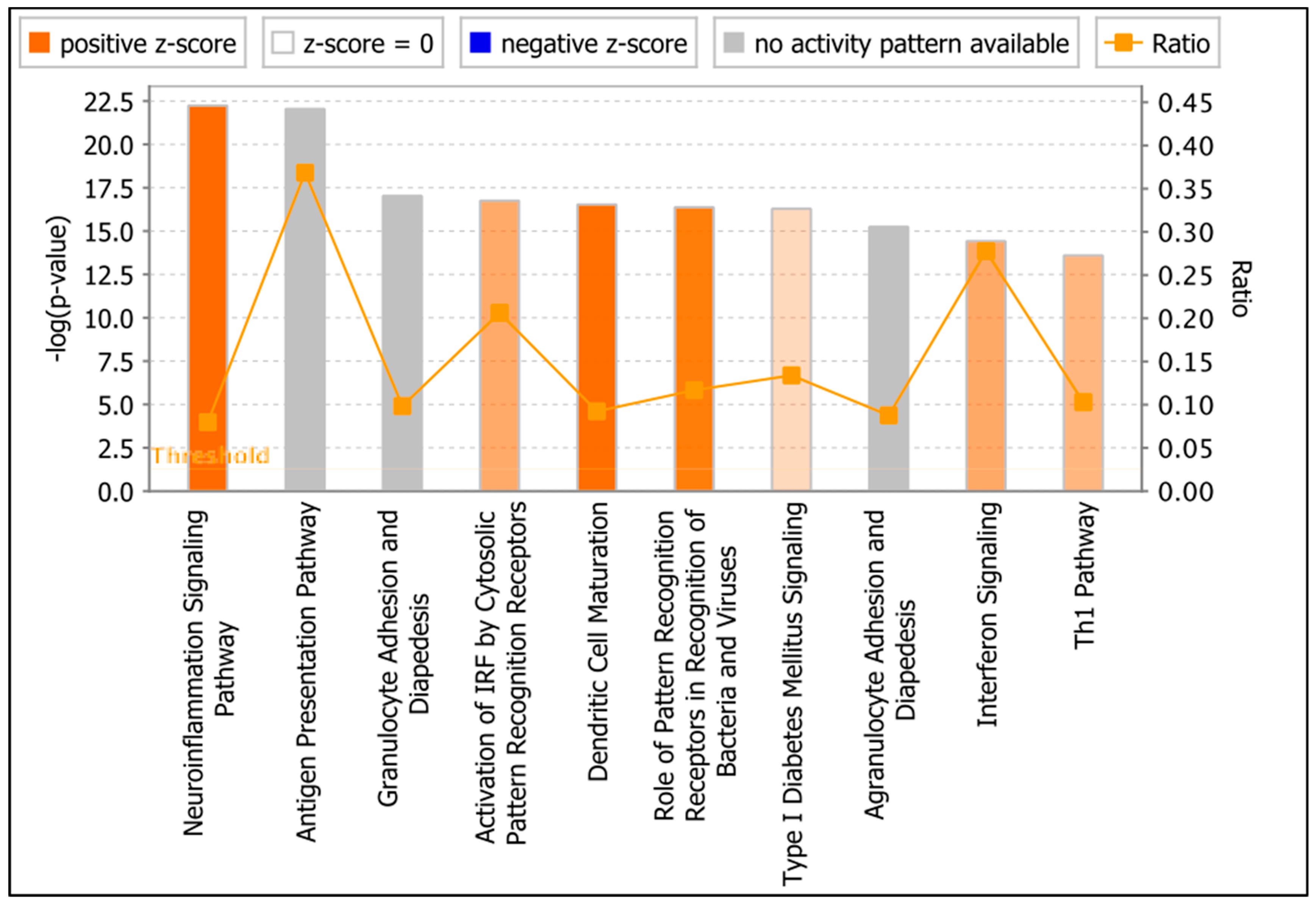
3.6. Functional Analysis of ZIKV-Modulated mRNAs
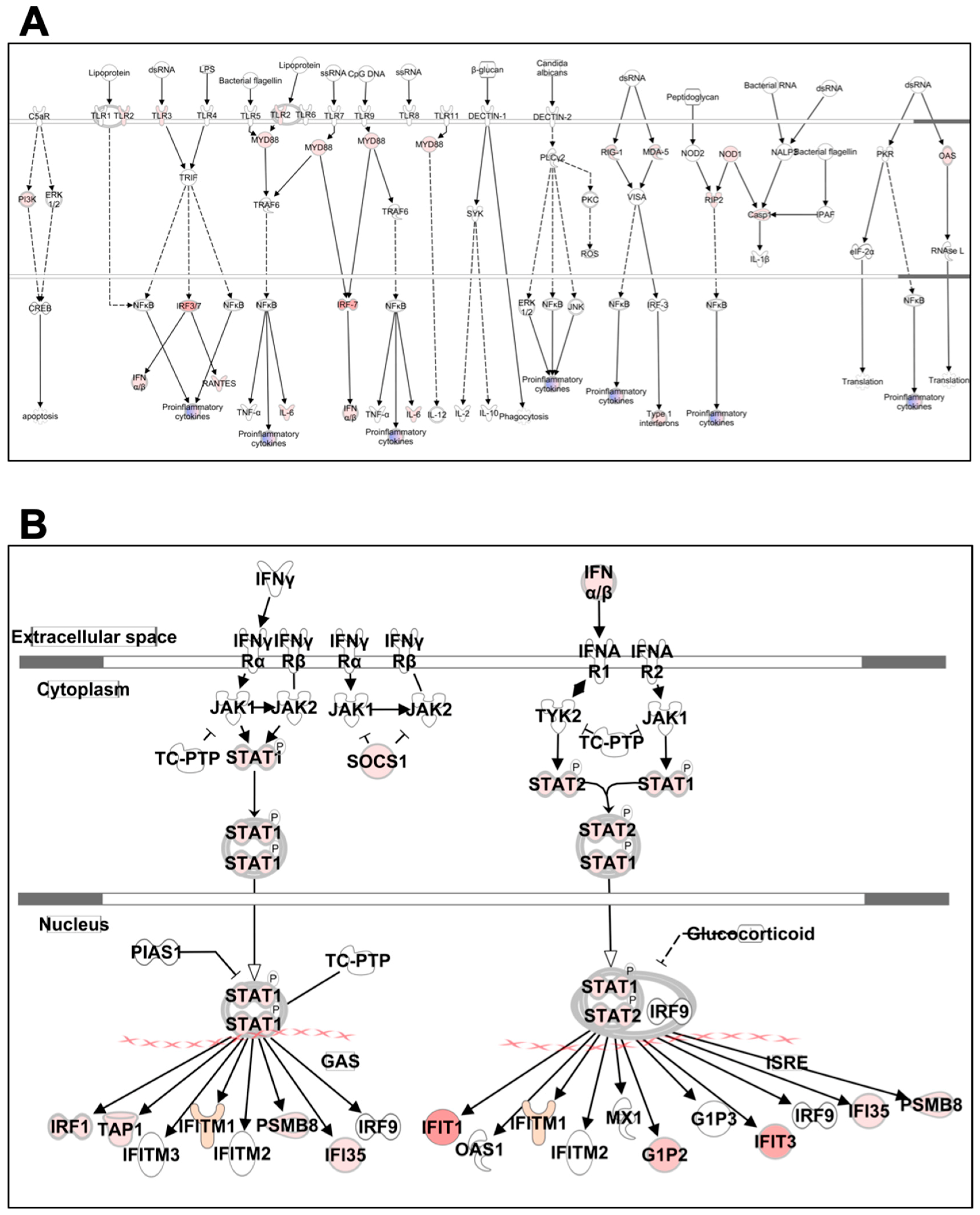
3.6.1. Pattern Recognition Receptors
3.6.2. IFN Signaling
3.7. Network Analysis of Expression of miRNAs and mRNAs From ZIKV-Infected Neurons
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Musso, D.; Gubler, D.J. Zika virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed]
- Klase, Z.A.; Khakhina, S.; Schneider Ade, B.; Callahan, M.V.; Glasspool-Malone, J.; Malone, R. Zika fetal neuropathogenesis: Etiology of a viral syndrome. PLoS Negl. Trop. Dis. 2016, 10, e0004877. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré syndrome outbreak associated with zika virus infection in french polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Tang, H.; Hammack, C.; Ogden, S.C.; Wen, Z.; Qian, X.; Li, Y.; Yao, B.; Shin, J.; Zhang, F.; Lee, E.M.; et al. Zika virus infects human cortical neural progenitors and attenuates their growth. Cell Stem Cell 2016, 18, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Anfasa, F.; Siegers, J.Y.; van der Kroeg, M.; Mumtaz, N.; Stalin Raj, V.; de Vrij, F.M.S.; Widagdo, W.; Gabriel, G.; Salinas, S.; Simonin, Y.; et al. Phenotypic differences between asian and african lineage zika viruses in human neural progenitor cells. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Gaburro, J.; Bhatti, A.; Sundaramoorthy, V.; Dearnley, M.; Green, D.; Nahavandi, S.; Paradkar, P.N.; Duchemin, J.B. Zika virus-induced hyper excitation precedes death of mouse primary neuron. Virol. J. 2018, 15, 79. [Google Scholar] [CrossRef]
- Rosenfeld, A.B.; Doobin, D.J.; Warren, A.L.; Racaniello, V.R.; Vallee, R.B. Replication of early and recent zika virus isolates throughout mouse brain development. Proc. Natl. Acad. Sci. USA 2017, 114, 12273–12278. [Google Scholar] [CrossRef]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A mouse model of zika virus pathogenesis. Cell host microbe 2016, 19, 720–730. [Google Scholar] [CrossRef]
- Rossi, S.L.; Tesh, R.B.; Azar, S.R.; Muruato, A.E.; Hanley, K.A.; Auguste, A.J.; Langsjoen, R.M.; Paessler, S.; Vasilakis, N.; Weaver, S.C. Characterization of a novel murine model to study zika virus. Am. J. Trop Med. Hyg. 2016, 94, 1362–1369. [Google Scholar] [CrossRef]
- Duggal, N.K.; Ritter, J.M.; McDonald, E.M.; Romo, H.; Guirakhoo, F.; Davis, B.S.; Chang, G.J.; Brault, A.C. Differential neurovirulence of african and asian genotype zika virus isolates in outbred immunocompetent mice. Am. J. Trop Med. Hyg. 2017, 97, 1410–1417. [Google Scholar] [CrossRef]
- Manangeeswaran, M.; Ireland, D.D.; Verthelyi, D. Zika (prvabc59) infection is associated with t cell infiltration and neurodegeneration in cns of immunocompetent neonatal c57bl/6 mice. PLoS pathog. 2016, 12, e1006004. [Google Scholar] [CrossRef] [PubMed]
- Van den Pol, A.N.; Mao, G.; Yang, Y.; Ornaghi, S.; Davis, J.N. Zika virus targeting in the developing brain. J. Neurosci. 2017, 37, 2161–2175. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, N.C.; Nogueira, J.S.; Ressio, R.A.; Cirqueira, C.S.; Kimura, L.M.; Fernandes, K.R.; Cunha, M.S.; Souza, R.P.; Guerra, J.M. Experimental zika virus infection induces spinal cord injury and encephalitis in newborn swiss mice. Exp. Toxicol. Pathol. 2017, 69, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Abraham, R.; Shim, B.S.; Choe, H.; Page, D.T. Zika virus infection during the period of maximal brain growth causes microcephaly and corticospinal neuron apoptosis in wild type mice. Sci. Rep. 2016, 6, 34793. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, M.; Condorelli, G.; Croce, C.M. Micrornas in diseases and drug response. Curr. Opin. Pharmacol. 2008, 8, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Mouradian, M.M. Micrornas in neurodegenerative diseases and their therapeutic potential. Pharmacol. Ther. 2012, 133, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Pauley, K.M.; Chan, E.K. Micrornas and their emerging roles in immunology. Ann. NY Acad. Sci. 2008, 1143, 226–239. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Rao, D.S.; Baltimore, D. Microrna regulation of inflammatory responses. Annu. Rev. Immunol. 2012, 30, 295–312. [Google Scholar] [CrossRef]
- Zhai, A.; Qian, J.; Kao, W.; Li, A.; Li, Y.; He, J.; Zhang, Q.; Song, W.; Fu, Y.; Wu, J.; et al. Borna disease virus encoded phosphoprotein inhibits host innate immunity by regulating mir-155. Antiviral Res. 2013, 98, 66–75. [Google Scholar] [CrossRef]
- Bolisetty, M.T.; Dy, G.; Tam, W.; Beemon, K.L. Reticuloendotheliosis virus strain t induces Mir-155, which targets JARID2 and promotes cell survival. J. Virol. 2009, 83, 12009–12017. [Google Scholar] [CrossRef]
- Swaminathan, G.; Rossi, F.; Sierra, L.J.; Gupta, A.; Navas-Martin, S.; Martin-Garcia, J. A role for microrna-155 modulation in the anti-HIV-1 effects of toll-like receptor 3 stimulation in macrophages. PLoS Pathog. 2012, 8, e1002937. [Google Scholar] [CrossRef] [PubMed]
- Pareek, S.; Roy, S.; Kumari, B.; Jain, P.; Banerjee, A.; Vrati, S. Mir-155 induction in microglial cells suppresses japanese encephalitis virus replication and negatively modulates innate immune responses. J. Neuroinflammation 2014, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Fabozzi, G.; Nabel, C.S.; Dolan, M.A.; Sullivan, N.J. Ebolavirus proteins suppress the effects of small interfering rna by direct interaction with the mammalian rna interference pathway. J. Virol. 2011, 85, 2512–2523. [Google Scholar] [CrossRef]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular micrornas contribute to hiv-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis c virus rna abundance by a liver-specific microrna. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, I.M.; Cheng, G.; Wieland, S.; Volinia, S.; Croce, C.M.; Chisari, F.V.; David, M. Interferon modulation of cellular micrornas as an antiviral mechanism. Nature 2007, 449, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Bavia, L.; Mosimann, A.L.; Aoki, M.N.; Duarte Dos Santos, C.N. A glance at subgenomic flavivirus rnas and micrornas in flavivirus infections. Virol. J. 2016, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; He, Z.; Hu, Y.; Wen, W.; Lin, C.; Yu, J.; Pan, J.; Li, R.; Deng, H.; Liao, S.; et al. MicroRNA-30e* suppresses dengue virus replication by promoting NF-κB -dependent IFN production. PLoS Negl. Trop Dis. 2014, 8, e3088. [Google Scholar] [CrossRef]
- Escalera-Cueto, M.; Medina-Martinez, I.; del Angel, R.M.; Berumen-Campos, J.; Gutierrez-Escolano, A.L.; Yocupicio-Monroy, M. Let-7c overexpression inhibits dengue virus replication in human hepatoma Huh-7 cells. Virus Res. 2015, 196, 105–112. [Google Scholar] [CrossRef]
- Slonchak, A.; Shannon, R.P.; Pali, G.; Khromykh, A.A. Human microrna MiR-532-5p exhibits antiviral activity against west nile virus via suppression of host genes SESTD1 and TAB3 required for virus replication. J. virol. 2015, 90, 2388–2402. [Google Scholar] [CrossRef]
- Smith, J.L.; Grey, F.E.; Uhrlaub, J.L.; Nikolich-Zugich, J.; Hirsch, A.J. Induction of the cellular microrna, hs_154, by west nile virus contributes to virus-mediated apoptosis through repression of antiapoptotic factors. J. Virol. 2012, 86, 5278–5287. [Google Scholar] [CrossRef] [PubMed]
- Heiss, B.L.; Maximova, O.A.; Pletnev, A.G. Insertion of microrna targets into the flavivirus genome alters its highly neurovirulent phenotype. J. virol. 2011, 85, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nerurkar, V.R. Integrated analysis of micrornas and their disease related targets in the brain of mice infected with west nile virus. Virology 2014, 452–453, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Shin, O.S.; Kumar, M.; Yanagihara, R.; Song, J.W. Hantaviruses induce cell type- and viral species-specific host microrna expression signatures. Virology 2013, 446, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Geiss, G.K.; Bumgarner, R.E.; Birditt, B.; Dahl, T.; Dowidar, N.; Dunaway, D.L.; Fell, H.P.; Ferree, S.; George, R.D.; Grogan, T.; et al. Direct multiplexed measurement of gene expression with color-coded probe pairs. Nat. Biotechnol. 2008, 26, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakayama, I.; Song, K.J.; Nerurkar, V.R.; Yoshida, S.; Garruto, R.M. Slow dendritic transport of dissociated mouse hippocampal neurons exposed to aluminum. Brain Res. 1997, 748, 237–240. [Google Scholar] [CrossRef]
- Forest, K.H.; Alfulaij, N.; Arora, K.; Taketa, R.; Sherrin, T.; Todorovic, C.; Lawrence, J.L.M.; Yoshikawa, G.T.; Ng, H.L.; Hruby, V.J.; et al. Protection against beta-amyloid neurotoxicity by a non-toxic endogenous n-terminal beta-amyloid fragment and its active hexapeptide core sequence. J. Neurochem. 2018, 144, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Seong, R.K.; Kumar, M.; Shin, O.S. Favipiravir and ribavirin inhibit replication of asian and african strains of zika virus in different cell models. Viruses 2018, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Krause, K.K.; Azouz, F.; Nakano, E.; Nerurkar, V.R. A guinea pig model of zika virus infection. Virol. J. 2017, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Lo, Y.; Chapagain, M.; Lum, S.; Kumar, M.; Gurjav, U.; Luo, H.; Nakatsuka, A.; Nerurkar, V.R. West nile virus infection modulates human brain microvascular endothelial cells tight junction proteins and cell adhesion molecules: Transmigration across the in vitro blood-brain barrier. Virology 2009, 385, 425–433. [Google Scholar] [CrossRef]
- Kumar, M.; Belcaid, M.; Nerurkar, V.R. Identification of host genes leading to west nile virus encephalitis in mice brain using RNA-Seq analysis. Sci. Rep. 2016, 6, 26350. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Verma, S.; Nerurkar, V.R. Pro-inflammatory cytokines derived from west nile virus (WNV)-infected SK-N-SH cells mediate neuroinflammatory markers and neuronal death. J. Neuroinflammation 2010, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Roe, K.; Nerurkar, P.V.; Namekar, M.; Orillo, B.; Verma, S.; Nerurkar, V.R. Impaired virus clearance, compromised immune response and increased mortality in type 2 diabetic mice infected with west nile virus. PLoS ONE 2012, 7, e44682. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Roe, K.; Orillo, B.; Muruve, D.A.; Nerurkar, V.R.; Gale, M., Jr.; Verma, S. Inflammasome adaptor protein apoptosis-associated speck-like protein containing card (ASC) is critical for the immune response and survival in west nile virus encephalitis. J. Virol. 2013, 87, 3655–3667. [Google Scholar] [CrossRef] [PubMed]
- Pijlman, G.P. Flavivirus rnai suppression: Decoding non-coding rna. Curr. Opin. Virol. 2014, 7, 55–60. [Google Scholar] [CrossRef]
- Schnettler, E.; Sterken, M.G.; Leung, J.Y.; Metz, S.W.; Geertsema, C.; Goldbach, R.W.; Vlak, J.M.; Kohl, A.; Khromykh, A.A.; Pijlman, G.P. Noncoding flavivirus rna displays rna interference suppressor activity in insect and mammalian cells. J. Virol. 2012, 86, 13486–13500. [Google Scholar] [CrossRef]
- Kakumani, P.K.; Ponia, S.S.; S., R.K.; Sood, V.; Chinnappan, M.; Banerjea, A.C.; Medigeshi, G.R.; Malhotra, P.; Mukherjee, S.K.; Bhatnagar, R.K. Role of RNA interference (RNAi) in dengue virus replication and identification of ns4b as an rnai suppressor. J. Virol. 2013, 87, 8870–8883. [Google Scholar] [CrossRef]
- Moon, S.L.; Dodd, B.J.; Brackney, D.E.; Wilusz, C.J.; Ebel, G.D.; Wilusz, J. Flavivirus sfrna suppresses antiviral rna interference in cultured cells and mosquitoes and directly interacts with the rnai machinery. Virology 2015, 485, 322–329. [Google Scholar] [CrossRef]
- Fredericksen, B.L. The neuroimmune response to west nile virus. J. Neurovirol. 2014, 20, 113–121. [Google Scholar] [CrossRef]
- Fredericksen, B.L.; Keller, B.C.; Fornek, J.; Katze, M.G.; Gale, M., Jr. Establishment and maintenance of the innate antiviral response to west nile virus involves both rig-i and mda5 signaling through IPS-1. J. Virol. 2008, 82, 609–616. [Google Scholar] [CrossRef]
- Errett, J.S.; Suthar, M.S.; McMillan, A.; Diamond, M.S.; Gale, M., Jr. The essential, nonredundant roles of rig-i and mda5 in detecting and controlling west nile virus infection. J. Virol. 2013, 87, 11416–11425. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, D.K.; Gupta, M.; Kumawat, K.L.; Basu, A. Nlrp3 inflammasome: Key mediator of neuroinflammation in murine japanese encephalitis. PLoS ONE 2012, 7, e32270. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Gale, M., Jr. Cell-intrinsic innate immune control of west nile virus infection. Trends Immunol. 2012, 33, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Wang, P.Y.; Su, D.F.; Liu, X. Mirna-124 in immune system and immune disorders. Front. Immunol. 2016, 7, 406. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Broering, R.; Trippler, M.; Wu, J.; Zhang, E.; Zhang, X.; Gerken, G.; Lu, M.; Schlaak, J.F. Microrna-155 controls toll-like receptor 3- and hepatitis C virus-induced immune responses in the liver. J. Viral. Hepat. 2014, 21, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Dickey, L.L.; Hanley, T.M.; Huffaker, T.B.; Ramstead, A.G.; O’Connell, R.M.; Lane, T.E. MicroRNA 155 and viral-induced neuroinflammation. J. Neuroimmunol. 2017, 308, 17–24. [Google Scholar] [CrossRef]
- Thounaojam, M.C.; Kaushik, D.K.; Kundu, K.; Basu, A. MicroRNA-29b modulates japanese encephalitis virus-induced microglia activation by targeting tumor necrosis factor alpha-induced protein 3. J. Neurochem. 2014, 129, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Buggele, W.A.; Horvath, C.M. Microrna profiling of sendai virus-infected a549 cells identifies MiR-203 as an interferon-inducible regulator of IFIT1/ISG56. J. Virol. 2013, 87, 9260–9270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, J.; Li, J.; Yang, Y.; Kang, X.; Li, Y.; Wu, X.; Zhu, Q.; Zhou, Y.; Hu, Y. Upregulation of microrna-203 in influenza A virus infection inhibits viral replication by targeting dr1. Sci. Rep. 2018, 8, 6797. [Google Scholar] [CrossRef]
- Song, M.S.; Rossi, J.J. Molecular mechanisms of dicer: Endonuclease and enzymatic activity. Biochem. J. 2017, 474, 1603–1618. [Google Scholar] [CrossRef] [PubMed]
- Kozak, R.A.; Majer, A.; Biondi, M.J.; Medina, S.J.; Goneau, L.W.; Sajesh, B.V.; Slota, J.A.; Zubach, V.; Severini, A.; Safronetz, D.; et al. MicroRNA and mRNA dysregulation in astrocytes infected with zika virus. Viruses 2017, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Saldana, M.A.; Etebari, K.; Hart, C.E.; Widen, S.G.; Wood, T.G.; Thangamani, S.; Asgari, S.; Hughes, G.L. Zika virus alters the microrna expression profile and elicits an rnai response in aedes aegypti mosquitoes. PLoS Negl. Trop Dis. 2017, 11, e0005760. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.N.; Holanda, G.M.; Pinto Silva, E.V.; Casseb, S.M.M.; Melo, K.F.L.; Carvalho, C.A.M.; Lima, J.A.; Vasconcelos, P.F.C.; Cruz, A.C.R. Zika virus alters the expression profile of microrna-related genes in liver, lung, and kidney cell lineages. Viral. Immunol. 2018, 31, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Weidmer, A.; Liu, C.G.; Volinia, S.; Croce, C.M.; Lieberman, P.M. Epstein–Barr virus-induced mir-155 attenuates NF-KB signaling and stabilizes latent virus persistence. J.Virol. 2008, 82, 10436–10443. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Croce, C.M.; Michaille, J.J. Mir-155: On the crosstalk between inflammation and cancer. Int. Rev. Immunol. 2009, 28, 264–284. [Google Scholar] [CrossRef] [PubMed]
- Thounaojam, M.C.; Kundu, K.; Kaushik, D.K.; Swaroop, S.; Mahadevan, A.; Shankar, S.K.; Basu, A. MicroRNA 155 regulates Japanese encephalitis virus-induced inflammatory response by targeting src homology 2-containing inositol phosphatase 1. J. Virol. 2014, 88, 4798–4810. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Luo, Z.M.; Guo, X.M.; Su, D.F.; Liu, X. An updated role of microrna-124 in central nervous system disorders: A review. Front. Cell Neurosci. 2015, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Papagiannakopoulos, T.; Kosik, K.S. MicroRNA-124: Micromanager of neurogenesis. Cell Stem Cell 2009, 4, 375–376. [Google Scholar] [CrossRef]
- Roy, B.; Dunbar, M.; Shelton, R.C.; Dwivedi, Y. Identification of microRNA-124-3p as a putative epigenetic signature of major depressive disorder. Neuropsychopharmacology 2017, 42, 864–875. [Google Scholar] [CrossRef]
- Ge, Q.; Gerard, J.; Noel, L.; Scroyen, I.; Brichard, S.M. MicroRNAs regulated by adiponectin as novel targets for controlling adipose tissue inflammation. Endocrinology 2012, 153, 5285–5296. [Google Scholar] [CrossRef] [PubMed]
- Meissner, L.; Gallozzi, M.; Balbi, M.; Schwarzmaier, S.; Tiedt, S.; Terpolilli, N.A.; Plesnila, N. Temporal profile of microRNA expression in contused cortex after traumatic brain injury in mice. J. Neurotrauma. 2016, 33, 713–720. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession No.) | Primer Sequence (5’‒3’) |
|---|---|
| IFIT1 (NM_008331) | |
| Forward | GTTGTTGTTGTTGTTCGT |
| Reverse | CAGCAGGAATCAGTTGTG |
| IL6 (NM_031168) | |
| Forward | ATCCAGTTGCCTTCTTGGGACTGA |
| Reverse | TAAGCCTCCGACTTGTGAAGTGGT |
| IFIT3 (NM_010501) | |
| Forward | GTCCTCTCTACTCTTTGG |
| Reverse | CATCCTCTGTCTTCTCTC |
| Caspase1 (NM_009807) | |
| Forward | GGAAGCAATTTATCAACTCAGTG |
| Reverse | GCCTTGTCCATAGCAGTAATG |
| Dicer-1 (NM_148948) | |
| Forward | TGTCATCTTGCGATTCTA |
| Reverse | TCTCTTCCAATTCCTCTG |
| DROSHA (NM_001130149) | |
| Forward | CTTCAACAGTTACCAGAAC |
| Reverse | CCTTTGGGAGTGAGTATG |
| AGO1 (NM_001317174) | |
| Forward | CCTGTGTATGATGGAAAGA |
| Reverse | CACTTGATGGAGACCTTAA |
| AGO2 (NM_153178) | |
| Forward | GGAGAACAATCAAACTACAG |
| Reverse | CAGATTCTTCCTTCCATCA |
| DGCR8 (NM_033324) | |
| Forward | CAGATAAGAAGGATGAGGAA |
| Reverse | GCTCCAAATTGTCAGTAAA |
| miR-155 (MIMAT0000165) | |
| Forward | UUAAUGCUAAUUGUGAUAGGGGUA |
| miR-203 (MIMAT0000236) | |
| Forward | GUGAAAUGUUUAGGACCACUAG |
| miR-29a (MIMAT0000535) | |
| Forward | UAGCACCAUCUGAAAUCGGUUA |
| miR-124-3p (MIMAT0000134) | |
| Forward | UAAGGCACGCGGUGAAUGCC |
| miRNA | Fold-change (24 h) | miRNA | Fold-change (48 h) |
|---|---|---|---|
| miR-155 | 18.29 | miR-155 | 10.61 |
| miR-3471 | 4.96 | miR-203 | 4.61 |
| miR-2145 | 2.33 | miR-1902 | 2.95 |
| miR-29a | 2.09 | miR-29b | 2.1 |
| miR-29b | 2.05 | miR-29a | 2.08 |
| miRNA | Fold-change | miRNA | Fold-change |
|---|---|---|---|
| miR-124-3p | −4.02 | miR-144 | −2.15 |
| miR-M1-1 | −2.97 | miR-201 | −2.15 |
| miR-1892 | −2.85 | miR-764-5p | −2.15 |
| miR-883a-3p | −2.7 | miR-1895 | −2.13 |
| miR-879 | −2.69 | miR-871 | −2.12 |
| miR-669g | −2.65 | miR-m108-1 | −2.12 |
| miR-654-3p | −2.62 | miR-1928 | −2.11 |
| miR-339-3p | −2.52 | miR-1941-5p | −2.11 |
| miR-1960 | −2.51 | miR-759 | −2.11 |
| miR-207 | −2.51 | miR-1187 | −2.1 |
| miR-2861 | −2.51 | miR-666-3p | −2.1 |
| miR-412 | −2.48 | miR-1966 | −2.09 |
| miR-741 | −2.47 | miR-1942 | −2.08 |
| miR-770-5p | −2.41 | miR-1946a | −2.08 |
| miR-673-5p | −2.38 | miR-1967 | −2.08 |
| miR-1941-3p | −2.37 | miR-m01-1 | −2.07 |
| miR-493 | −2.33 | miR-383 | −2.06 |
| miR-465a-3p | −2.31 | miR-186 | −2.05 |
| miR-1956 | −2.26 | miR-323-5p | −2.05 |
| miR-1194 | −2.25 | miR-M1-3 | −2.05 |
| miR-709 | −2.25 | miR-433 | −2.04 |
| miR-m107-1-5p | −2.25 | miR-767 | −2.04 |
| miR-877 | −2.23 | miR-1188 | −2.03 |
| miR-331-5p | −2.22 | miR-694 | −2.03 |
| miR-483 | −2.22 | miR-665 | −2.02 |
| miR-675-5p | −2.2 | miR-2139 | −2.02 |
| miR-1957 | −2.19 | miR-883a-5p | −2.02 |
| miR-M23-1-3p | −2.19 | miR-1943 | −2 |
| miR-1898 | −2.18 | miR-346 | −2 |
| miR-1940 | −2.17 | miR-764-3p | −2 |
| miR-1894-5p | −2.16 | miR-710 | −2 |
| miRNA | Fold-change | miRNA | Fold-change |
|---|---|---|---|
| miR-124-3p | −6.68 | miR-m21-1 | −2.15 |
| miR-883a-3p | −3.93 | miR-335-3p | −2.14 |
| miR-2137 | −2.92 | miR-1957 | −2.12 |
| miR-2133 | −2.74 | miR-764-5p | −2.11 |
| miR-714 | −2.56 | miR-1194 | −2.08 |
| miR-669g | −2.54 | miR-683 | −2.08 |
| miR-467g | −2.5 | miR-509-5p | −2.07 |
| miR-1188 | −2.41 | miR-463 | −2.06 |
| miR-879 | −2.33 | miR-741 | −2.06 |
| miR-760 | −2.25 | miR-761 | −2.05 |
| miR-298 | −2.21 | miR-710 | −2.03 |
| miR-m01-3 | −2.21 | miR-346 | −2.01 |
| miR-764-3p | −-2.2 | miR-882 | −2.01 |
| miR-370 | −2.18 | miR-1898 | −2.01 |
| miR-1892 | −2.16 | miR-759 | −2 |
| miR-m01-2 | −2.16 | miR-433 | −2 |
| miR-133b | −2.15 | miR-709 | −2 |
| miR-1894-5p | −2.15 | miR-1956 | −2 |
| miR-666-3p | −2.15 | miR-483 | −2 |
| miR-877 | −2.15 | miR-1941-5p | −2 |
| Biological Process/Pathway | p-Value | Number of miRNAs |
|---|---|---|
| Cancer | 4.02 × 10−10 | 18 |
| Organismal Injury and Abnormalities | 4.02 × 10−10 | 23 |
| Reproductive System Disease | 4.02 × 10−10 | 18 |
| Immunological Disease | 6.98 × 10−10 | 13 |
| Inflammatory Disease | 6.98 × 10−10 | 13 |
| Inflammatory Response | 6.98 × 10−10 | 11 |
| Neurological Disease | 6.98 × 10−10 | 10 |
| Connective Tissue Disorders | 1.46 × 10−8 | 10 |
| Respiratory Disease | 1.46 × 10−8 | 7 |
| Nervous System Development and Function | 7.20 × 10−7 | 6 |
| mRNA | Fold-Change (24 h) | Fold-Change (48 h) | mRNA | Fold-Change (24 h) | Fold-Change (48 h) |
|---|---|---|---|---|---|
| Rsad2 | 26.44 | 244.24 | Ccl2 | 3.91 | 14.32 |
| Cxcl10 | 22.31 | 113.73 | Ifna4 | 2.85 | 14.02 |
| Ifit3 | 21.63 | 103.85 | Nlrc5 | 3.42 | 13.39 |
| Ifi44 | 18.36 | 103.12 | Psmb10 | 3.13 | 12.78 |
| Irf7 | 12.53 | 96.42 | Stat2 | 5.55 | 12.37 |
| Ifit1 | 24.52 | 75.06 | H2-K1 | 2 | 12.34 |
| Isg15 | 20.58 | 69.08 | Cfb | 2.04 | 12.04 |
| Lcn2 | 27.07 | 68.88 | Ifi35 | 4.05 | 11.3 |
| Zbp1 | 13.57 | 55.1 | Ddx58 | 3.84 | 10.56 |
| Gbp5 | 12.51 | 44.43 | Psmb9 | 2.4 | 10.03 |
| Ifit2 | 2.53 | 41.91 | Tap1 | 2.77 | 9.85 |
| Ccl5 | 24.11 | 37.65 | C3 | 3.29 | 9.22 |
| Oas2 | 10.64 | 37.13 | Casp1 | 2.76 | 7.63 |
| Oasl1 | 5.15 | 34.55 | Socs1 | 2.13 | 6.63 |
| Usp18 | 9.7 | 32.04 | H2-Ab1 | 2.21 | 5.82 |
| Isg20 | 5.57 | 30.07 | Cxcl1 | 3.49 | 5.5 |
| H2-T23 | 3.61 | 28.33 | Ifitm1 | 3.94 | 5.39 |
| Ifih1 | 6.11 | 25.37 | Ccl12 | 3.68 | 5.38 |
| Ddx60 | 7.87 | 23.88 | Pml | 2 | 5.38 |
| Stat1 | 6.92 | 21.99 | Vcam1 | 2.23 | 5.24 |
| Bst2 | 4.92 | 21.62 | Myd88 | 2.38 | 4.87 |
| Mx2 | 7.55 | 20.85 | Cxcl9 | 2.83 | 4.7 |
| Psmb8 | 4.31 | 20.39 | Il6 | 2.07 | 3.63 |
| Ccl7 | 2.71 | 19.98 | Ptgs2 | 2.02 | 3.32 |
| Cd274 | 4.02 | 19.34 | Cxcl2 | 2.68 | 3.27 |
| Cmpk2 | 4.01 | 18.95 | Sbno2 | 3.05 | 2.86 |
| Xaf1 | 8.02 | 18.06 | Ccl4 | 2.52 | 2.84 |
| Herc6 | 3.68 | 17.33 | Lif | 2.75 | 2.47 |
| Ifnb1 | 2.97 | 16.75 | Il13ra1 | 2.01 | 2.33 |
| Tlr3 | 3.84 | 16.4 | Runx1 | 2 | 2.15 |
| Irgm2 | 6.29 | 15.13 | Litaf | 2 | 2.01 |
| mRNA | Fold-Change | mRNA | Fold-Change |
|---|---|---|---|
| Cxcl11 | 17.69 | Slamf7 | 2.81 |
| C2 | 9.88 | Tnfrsf14 | 2.76 |
| Tnfsf10 | 9.53 | Fcgr4 | 2.6 |
| Cxcl13 | 9.29 | Mill2 | 2.59 |
| Cd74 | 8.49 | Ptprc | 2.59 |
| C1ra | 7.96 | H2-Dma | 2.57 |
| H2-D1 | 7.64 | Nfkbia | 2.52 |
| H2-Aa | 7.35 | Tnfaip3 | 2.51 |
| Tap2 | 6.53 | Cxcl5 | 2.47 |
| Ccl11 | 5.49 | A2m | 2.46 |
| Fcgr1 | 5.32 | Ifna1 | 2.38 |
| H2-M3 | 5.25 | Il3ra | 2.36 |
| Ccrl2 | 4.9 | Ctss | 2.34 |
| Cd47 | 4.2 | Ripk2 | 2.34 |
| Tapbp | 3.97 | H2-DMb1 | 2.28 |
| C4b | 3.96 | Lck | 2.27 |
| Ifna2 | 3.96 | Cd80 | 2.26 |
| Irf1 | 3.88 | Cxcl16 | 2.19 |
| Tlr2 | 3.87 | Cybb | 2.19 |
| C1s1 | 3.56 | Icosl | 2.18 |
| Lbp | 3.44 | Cfi | 2.14 |
| Il7 | 3.3 | Irf2 | 2.1 |
| Irf5 | 3.03 | Nod1 | 2.02 |
| Cd69 | 2.99 | Atm | 2 |
| Serping1 | 2.91 | Axl | 2 |
| Flt3l | 2.87 | H2-Eb1 | 2 |
| Bid | 2.85 | Relb | 2 |
| mRNA | Fold-Change (48 h) | mRNA | Fold-Change (48 h) |
|---|---|---|---|
| Cd36 | −2 | Cd207 | −2.19 |
| Elane | −2 | Timd4 | −2.22 |
| Il17f | −2 | Xcl1 | −2.31 |
| Ticam2 | −2 | Card9 | −2.65 |
| Pax5 | −2.02 | Sh2d1b1 | −2.76 |
| Il1rapl2 | −2.15 | Mpped1 | −2.87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azouz, F.; Arora, K.; Krause, K.; Nerurkar, V.R.; Kumar, M. Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons. Viruses 2019, 11, 162. https://doi.org/10.3390/v11020162
Azouz F, Arora K, Krause K, Nerurkar VR, Kumar M. Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons. Viruses. 2019; 11(2):162. https://doi.org/10.3390/v11020162
Chicago/Turabian StyleAzouz, Francine, Komal Arora, Keeton Krause, Vivek R. Nerurkar, and Mukesh Kumar. 2019. "Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons" Viruses 11, no. 2: 162. https://doi.org/10.3390/v11020162
APA StyleAzouz, F., Arora, K., Krause, K., Nerurkar, V. R., & Kumar, M. (2019). Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons. Viruses, 11(2), 162. https://doi.org/10.3390/v11020162