Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome

 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Analysis
2.2. RT-qPCR
2.3. Generation of CHIKV RNA Synthetic Transcript
2.4. Sensitivity
2.5. Specificity
3. Results
3.1. CHIKV RT-qPCR Assays and Selection
3.2. Analytical Sensitivity of Duo CHIKV RT-qPCR Assay
3.3. Specificity of Duo CHIKV RT-qPCR Assay
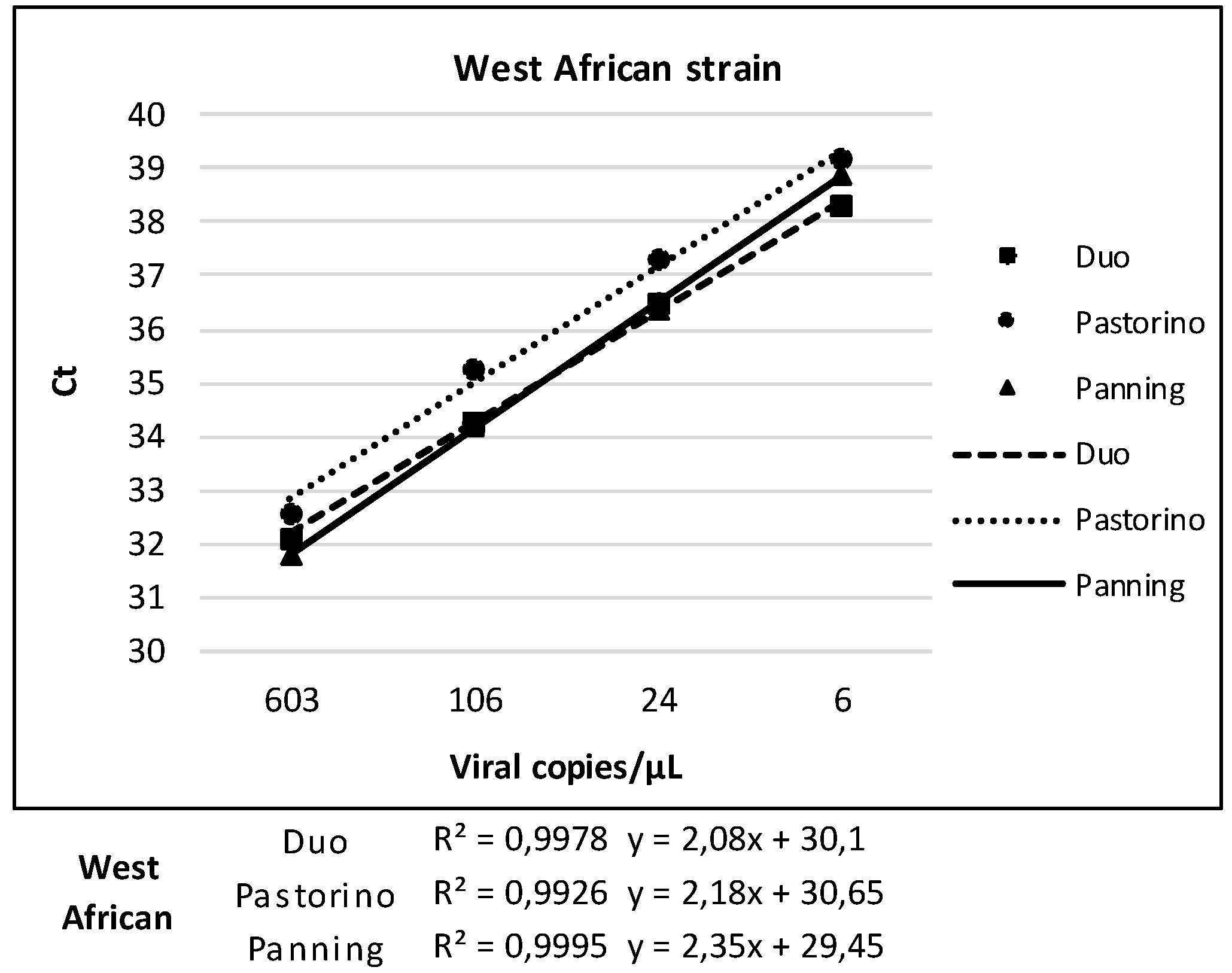
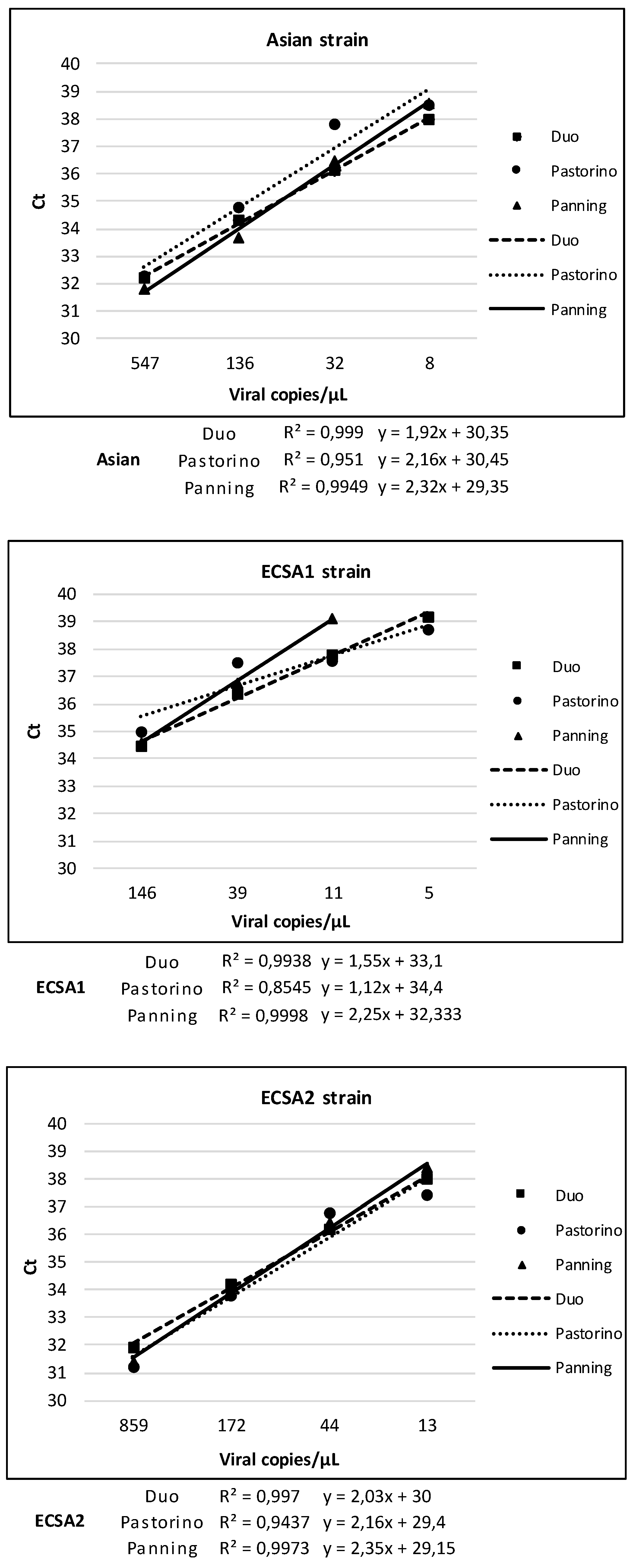
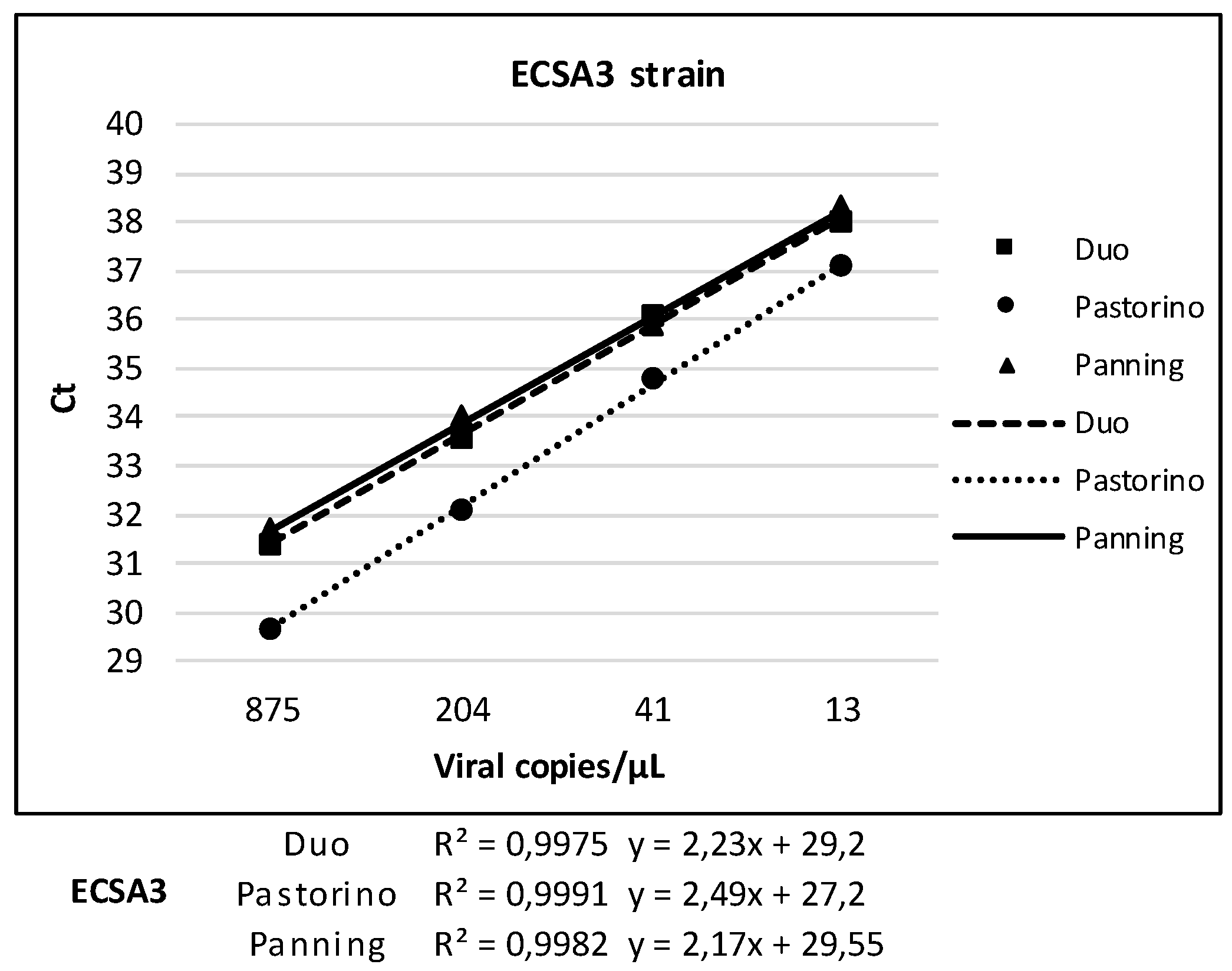
3.4. Linearity of Duo CHIKV RT-qPCR vs. Pastorino and Panning Mono-Target Assays
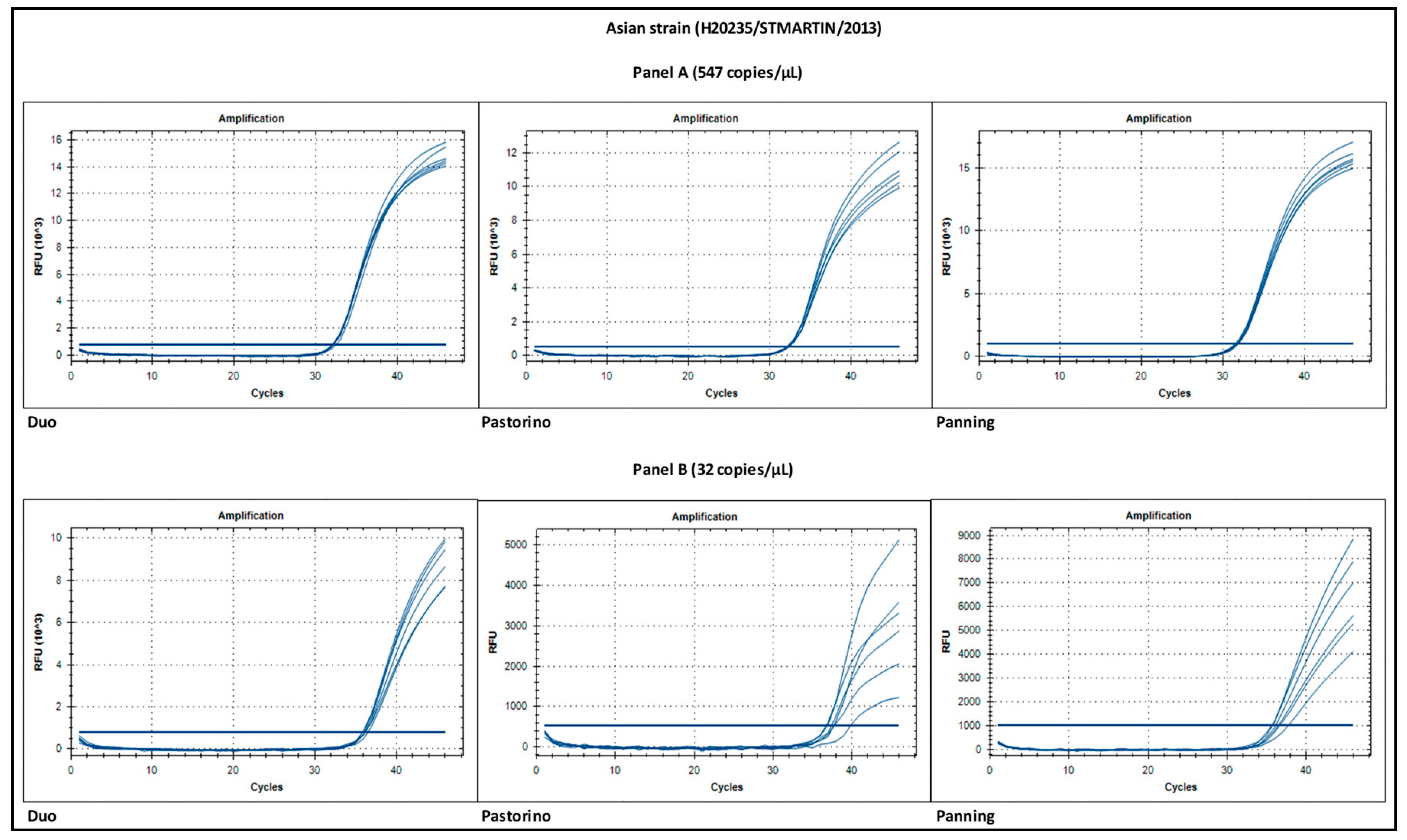
3.5. Signal Intensity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thiberville, S.D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; De Lamballerie, X. Chikungunya fever: Epidemiology, clinical syndrome, pathogenesis and therapy. Antivir. Res. 2013, 99, 345–370. [Google Scholar] [CrossRef] [PubMed]
- Chikungunya. Available online: https://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 26 April 2019).
- PAHO; WHO. Chikungunya: Data, Maps and Statistics. Available online: https://www.paho.org/hq/index.php?option=com_topics&view=rdmore&cid=5927&item=chikungunya&type=statistics&Itemid=40931&lang=en (accessed on 28 January 2019).
- Delisle, E.; Rousseau, C.; Broche, B.; Leparc-Goffart, I.; L’ambert, G.; Cochet, A.; Prat, C.; Foulongne, V.; Ferre, J.B.; Catelinois, O.; et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Eurosurveillance 2014, 20, 21108. [Google Scholar] [CrossRef]
- Calba, C.; Guerbois-Galla, M.; Franke, F.; Jeannin, C.; Auzet-Caillaud, M.; Grard, G.; Pigaglio, L.; Decoppet, A.; Weicherding, J.; Savaill, M.-C.; et al. Preliminary report of an autochthonous chikungunya outbreak in France, July to September 2017. Eurosurveillance 2017, 22, 17–00647. [Google Scholar] [CrossRef] [PubMed]
- Angelini, P.; Macini, P.; Finarelli, A.C.; Pol, C.; Venturelli, C.; Bellini, R.; Dottori, M. Chikungunya epidemic outbreak in Emilia-Romagna (Italy) during summer 2007. Parassitologia 2008, 50, 97–98. [Google Scholar] [PubMed]
- Rezza, G. Chikungunya is back in Italy: 2007–2017. J. Travel. Med. 2018, 25, tay004. [Google Scholar] [CrossRef] [PubMed]
- Pezzi, L.; Reusken, C.B.; Weaver, S.C.; Drexler, J.F.; Busch, M.; LaBeaud, A.D.; Diamond, M.S.; Vasilakis, N.; Drebot, M.A.; Siqueira, A.M.; et al. GloPID-R report on Chikungunya, O’nyong-nyong and Mayaro virus, part I: Biological diagnostics. Antivir. Res. 2019, 166, 66–81. [Google Scholar] [CrossRef]
- Pastorino, B.; Bessaud, M.; Grandadam, M.; Murri, S.; Tolou, H.J.; Peyrefitte, C.N. Development of a TaqMan RT-PCR assay without RNA extraction step for the detection and quantification of African Chikungunya viruses. J. Virol. Methods 2005, 124, 65–71. [Google Scholar] [CrossRef]
- Panning, M.; Grywna, K.; Van Esbroeck, M.; Emmerich, P.; Drosten, C. Chikungunya Fever in Travelers Returning to Europe from the Indian Ocean Region, 2006. Emerg. Infect. Dis. 2008, 14, 416–422. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya Virus in US Travelers Returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Ballesteros, G.; Gresh, L.; Mohamed-Hadley, A.; Tellez, Y.; Sahoo, M.K.; Abeynayake, J.; Balmaseda, A.; Harris, E.; Pinsky, B.A. Clinical evaluation of a single-reaction real-time RT-PCR for pan-dengue and chikungunya virus detection. J. Clin. Virol. 2016, 78, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Cecilia, D.; Kakade, M.; Alagarasu, K.; Patil, J.; Salunke, A.; Parashar, D.; Shah, P.S. Development of a multiplex real-time RT-PCR assay for simultaneous detection of dengue and chikungunya viruses. Arch. Virol. 2015, 160, 323–327. [Google Scholar] [CrossRef]
- Smith, D.R.; Lee, J.S.; Jahrling, J.; Kulesh, D.A.; Turell, M.J.; Groebner, J.L.; O’Guinn, M.L. Development of Field-Based Real-Time Reverse Transcription-Polymerase Chain Reaction Assays for Detection of Chikungunya and O’nyong-nyong Viruses in Mosquitoes. Am. J. Trop. Med. Hyg. 2009, 81, 679–684. [Google Scholar] [CrossRef]
- Edwards, C.J.; Welch, S.R.; Chamberlain, J.; Hewson, R.; Tolley, H.; Cane, P.A.; Lloyd, G. Molecular diagnosis and analysis of Chikungunya virus. J. Clin. Virol. 2007, 39, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Templer, S.P.; Seiverth, B.; Baum, P.; Stevens, W.; Seguin-Devaux, C.; Carmona, S. Improved Sensitivity of a Dual-Target HIV-1 Qualitative Test for Plasma and Dried Blood Spots. J. Clin. Microbiol. 2016, 54, 1877–1882. [Google Scholar] [CrossRef] [Green Version]
- Sizmann, D.; Glaubitz, J.; Simon, C.O.; Goedel, S.; Buergisser, P.; Drogan, D.; Hesse, M.; Kröh, M.; Simmler, P.; Dewald, M.; et al. Improved HIV-1 RNA quantitation by COBAS®AmpliPrep/COBAS TaqMan HIV-1 Test, v2.0 using a novel dual-target approach. J. Clin. Virol. 2010, 49, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Damond, F.; Avettand-Fenoel, V.; Collin, G.; Roquebert, B.; Plantier, J.C.; Ganon, A.; Sizmann, D.; Babiel, R.; Glaubitz, J.; Chaix, M.L.; et al. Evaluation of an Upgraded Version of the Roche Cobas AmpliPrep/Cobas TaqMan HIV-1 Test for HIV-1 Load Quantification. J. Clin. Microbiol. 2010, 48, 1413–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chudy, M.; Kress, J.; Halbauer, J.; Heiden, M.; Funk, M.B.; Nübling, C.M. Risk Minimization Measures for Blood Screening HIV-1 Nucleic Acid Amplification Technique Assays in Germany. Transfus. Med. Hemother. 2014, 41, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Korn, K.; Nübling, C.M.; Chudy, M.; Kress, J.; Horst, H.A.; Geusendam, G.; Hennig, H.; Sireis, W.; Rabenau, H.F.; et al. First transmission of human immunodeficiency virus Type 1 by a cellular blood product after mandatory nucleic acid screening in Germany. Transfusion 2009, 49, 1836–1844. [Google Scholar] [CrossRef]
- Edelmann, A.; Kalus, U.; Oltmann, A.; Stein, A.; Unbehaun, A.; Drosten, C.; Krüger, D.H.; Hofmann, J. Improvement of an ultrasensitive human immunodeficiency virus type 1 real-time reverse transcriptase-polymerase chain reaction targeting the long terminal repeat region. Transfusion 2010, 50, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Chudy, M.; Weber-Schehl, M.; Pichl, L.; Jork, C.; Kress, J.; Heiden, M.; Funk, M.B.; Nübling, C.M. Blood screening nucleic acid amplification tests for human immunodeficiency virus Type 1 may require two different amplification targets. Transfusion 2012, 52, 431–439. [Google Scholar] [CrossRef]
- Müller, B.; Nübling, C.M.; Kress, J.; Roth, W.K.; De Zolt, S.; Pichl, L. How safe is safe: new human immunodeficiency virus Type 1 variants missed by nucleic acid testing. Transfusion 2013, 53, 2422–2430. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; VanLandingham, D.L.; McGee, C.E.; Higgs, S. A Single Mutation in Chikungunya Virus Affects Vector Specificity and Epidemic Potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome Microevolution of Chikungunya Viruses Causing the Indian Ocean Outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.K.; Chen, H.L.; Yang, C.F.; Hsieh, S.C.; Juan, C.C.; Chang, S.M.; Yu, C.C.; Lin, L.H.; Huang, J.H.; King, C.C. Slower Rates of Clearance of Viral Load and Virus-Containing Immune Complexes in Patients with Dengue Hemorrhagic Fever. Clin. Infect. Dis. 2006, 43, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klitting, R.; Fischer, C.; Drexler, J.; Gould, E.; Roiz, D.; Paupy, C.; de Lamballerie, X. What Does the Future Hold for Yellow Fever Virus? (II). Genes 2018, 9, 425. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Primer/Probe | 5′ → 3′ Sequence | Target | Position c | Amplicon Size (nts) | Concentration |
|---|---|---|---|---|---|---|
| Pastorino et al. [9] | R-CHIK | CCAAATTGTCCYGGTCTTCCT | E1 | 10,568–10,588 | 208 | 900 nM |
| F-CHIK | AAGCTYCGCGTCCTTTACCAAG | 10,380–10,401 | 900 nM | |||
| P-CHIK | FAM-CCAATGTCYTCMGCCTGGACACCTTT-TAMRA | 10,479–10,504 | 200 nM | |||
| Panning et al. [10] | ChikAsI a | GGCAAACGCAGTGGTACTTCCT | nsp1 | 295–316 | 82 | 600 nM |
| ChikSI a | TGATCCCGACTCAACCATCCT | 234–254 | 600 nM | |||
| ChikAsII b | GGCAGACGCAGTGGTACTTCCT | 295–316 | 77 | 600 nM | ||
| ChikSII b | CCGACTCAACCATCCTGGAT | 239–258 | 600 nM | |||
| ChikP $ | FAM-TCCGACATCATCCTCCTTGCTGGC-TAMRA | 270–293 | 200 nM |
| Genus | Virus Species and Acronyms | Strain (Viral Load TCID50/mL) | Reference on EVAg or NCPV Catalog | |
|---|---|---|---|---|
| Alphavirus | Chikungunya virus | CHIKV-West African | UVE/CHIKV/1983/SN/WA 37997 (10 6.22) | 001V-02448 (EVAg) |
| CHIKV-Asian | H20235/STMARTIN/2013 (10 5.57) | 001N-EVAg1583 (EVAg) | ||
| CHIKV-ECSA1 | UVE/CHIKV/UNK/XX/ROSS (10 6.49) | 001v-EVA1455 (EVAg) | ||
| CHIKV-ECSA2 | UVE/CHIKV/2011/CD/Brazza_MRS1 (10 6.57) | 001v-EVA960 (EVAg) | ||
| CHIKV-ECSA3 | UVE/CHIKV/2006/RE/LR2006_OPY1 (10 8.49) | 001v-EVA83 (EVAg) | ||
| Mayaro virus | MAYV | UVE/MAYV/1954/TT/TC625 (10 8.82) | 001v-EVA502 (EVAg) | |
| O’nyong-nyong virus | ONNV | UVE/ONNV/UNK/SN/Dakar 234 (10 4.22) | 001v-EVA1044 (EVAg) | |
| Semliki Forest virus | SFV | UVE/SFV/UNK/XX/1745 (10 4.42) | 001V-02468 (EVAg) | |
| Ross River virus | RRV | 0005281v (10 8.16) | 0005281v (NCPV) | |
| Sindbis virus | SINV | UVE/SINV/UNK/EG/Egypt 339 (10 4.32) | 001V-02469 (EVAg) | |
| Western equine encephalitis virus | WEEV | UVE/WEEV/UNK/XX/47a (10 8.32) | 001v-EVA1479 (EVAg) | |
| Venezuelan equine encephalitis virus | VEEV | UVE/VEEV/UNK/XX/TC83 vaccine (10 9.42) | 001v-EVA1459 (EVAg) | |
| Eastern equine encephalitis virus | EEEV | UVE/EEEV/1999/XX/H178_99 (10 7.82) | 001v-EVA1480 (EVAg) | |
| Flavivirus | Zika virus | ZIKV | UVE/ZIKV/1947/UG/MR766 (10 4.32) | 001v-EVA143 (EVAg) |
| Dengue virus-1 | DENV-1 | UVE/DENV-1/2013/NC/CNR_17132 (10 7.57) | 001V-03151 (EVAg) | |
| Dengue virus-2 | DENV-2 | UVE/DENV-2/1996/PF/Papeete 341.175 (10 7.82) | 001V-03178 (EVAg) | |
| Dengue virus-3 | DENV-3 | UVE/DENV-3/UNK/RE/CNR_14448 (10 7.22) | 001V-03346 (EVAg) | |
| Dengue virus-4 | DENV-4 | UVE/DENV-4/2012/GF/CNR_16008 (10 8.49) | 001V-03366 (EVAg) | |
| Japanese encephalitis virus | JEV | UVE/JEV/2009/LA/CNS769 (10 5.57) | 001V-02217(EVAg) | |
| West Nile virus | WNV | UVE/WNV/2008/US/R94224 (10 7.32) | 001V-02224 (EVAg) | |
| Tick-borne encephalitis virus | TBEV | UVE/TBEV/2013/FR/32.11 WT-PCR (10 8.82) | 001V-02355 (EVAg) | |
| Yellow fever virus | YFV | UVE/YFV/UNK/XX/Vaccine strain 17D (10 6.32) | 001v-EVA67 (EVAg) | |
| Saint Louis encephalitis virus | SLEV | UVE/SLEV/UNK/US/MSI-7 (10 4.82) | 001v-EVA128 (EVAg) | |
| Usutu virus | USUV | UVE/USUV/1959/ZA/SAAR-1776 (10 5.32) | 001v-EVA138 (EVAg) | |
| Phlebovirus | Toscana virus | TOSV | UVE/TOSV/2013/FR/113 (10 7.42) | 001V-02461 (EVAg) |
| Rift Valley fever virus | RVFV | UVE/RVFV/UNK/XX/Smithburn vaccine (10 7.32) | 001V-02385 (EVAg) | |
| Sandly fever Sicilian virus | SFSV | UVE/SFSV/1943/IT/Sabin (10 6.82) | 001v-EVA77 (EVAg) | |
| West African | Duo | Pastorino | Panning | ||||
| Dilution | Viral RNA copies/µL | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) |
| E-6 | 603 | 6/6 | 32.1 (0.1) | 6/6 | 32.6 (0.2) | 6/6 | 31.8 (0.2) |
| 2E-7 | 106 | 6/6 | 34.3 (0.25) | 6/6 | 35.3 (0.2) | 6/6 | 34.2 (0.1) |
| 4E-8 | 24 | 6/6 | 36.5 (0.43) | 6/6 | 37.3 (0.2) | 6/6 | 36.4 (0.4) |
| 8E-9 | 6 | 5/6 | 38.3 (0.36) | 1/6 | 39.2 | 4/6 | 38.9 (0.2) |
| 1.6E-9 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| 3.2E-10 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| Asian | Duo | Pastorino | Panning | ||||
| Dilution | Viral RNA copies/µL | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) |
| E-6 | 547 | 6/6 | 32.2 (0.2) | 6/6 | 32.3 (0.1) | 6/6 | 31.8 (0.2) |
| 2E-7 | 136 | 6/6 | 34.3 (0.3) | 6/6 | 34.8 (0.3) | 6/6 | 33.7 (0.1) |
| 4E-8 | 32 | 6/6 | 36.1 (0.2) | 6/6 | 37.8 (1.1) | 6/6 | 36.5 (0.8) |
| 8E-9 | 8 | 6/6 | 38 (0.5) | 4/6 | 38.5 (1) | 2/6 | 38.6 (0.9) |
| 1.6E-9 | 2 | 0/6 | - | 3/6 | 39.5 (0.3) | 0/6 | - |
| 3.2E-10 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| ECSA1 | Duo | Pastorino | Panning | ||||
| Dilution | Viral RNA copies/µL | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) |
| E-6 | 146 | 6/6 | 34. 5 (0.2) | 6/6 | 35 (0.2) | 6/6 | 34.6 (0.5) |
| 2E-7 | 39 | 6/6 | 36.4 (0.4) | 6/6 | 37.5 (0.3) | 6/6 | 36.8 (0.1) |
| 4E-8 | 11 | 4/6 | 37.8 (0.6) | 4/6 | 37.6 (0.5) | 1/6 | 39.1 |
| 8E-9 | 5 | 1/6 | 39.2 | 1/6 | 38.7 | 0/6 | - |
| 1.6E-9 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| 3.2E-10 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| ECSA2 | Duo | Pastorino | Panning | ||||
| Dilution | Viral RNA copies/µL | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) |
| E-6 | 859 | 6/6 | 31.9 (0.2) | 6/6 | 31.2 (0.2) | 6/6 | 31.4 (0.1) |
| 2E-7 | 172 | 6/6 | 34.2 (0.1) | 6/6 | 33.8 (0.5) | 6/6 | 33.9 (0.3) |
| 4E-8 | 44 | 6/6 | 36.2 (0.2) | 5*/5 | 36.8 (0.7) | 6/6 | 36.4 (0.8) |
| 8E-9 | 13 | 6/6 | 38 (0.3) | 1/6 | 37.4 | 4/6 | 38.4 (0.6) |
| 1.6E-9 | 6 | 2/6 | 39.1 (0.03) | 0/6 | - | 0/6 | - |
| 3.2E-10 | - | 0/6 | - | 0/6 | - | 0/6 | - |
| ECSA3 | Duo | Pastorino | Panning | ||||
| Dilution | Viral RNA copies/µL | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) | Positive/tested | Ct, Mean (SD) |
| E-6 | 875 | 6/6 | 31.4 (0.1) | 6/6 | 29.7 (0.1) | 5*/5 | 31.7 (0.2) |
| 2E-7 | 204 | 6/6 | 33.6 (0.2) | 6/6 | 32.1 (0.2) | 6/6 | 34 (0.3) |
| 4E-8 | 41 | 6/6 | 36.1 (0.2) | 6/6 | 34.8 (0.5) | 6/6 | 35.9 (0.4) |
| 8E-9 | 13 | 5/6 | 38 (0.9) | 5/6 | 37.1 (0.7) | 5/6 | 38.3 (0.8) |
| 1.6E-9 | 5 | 1/6 | 39.5 | 2/6 | 39.2 (0.03) | 1/6 | 39,3 |
| 3.2E-10 | - | 0/6 | - | 0/6 | - | 0/6 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thirion, L.; Pezzi, L.; Corcostegui, I.; Dubot-Pérès, A.; Falchi, A.; de Lamballerie, X.; Charrel, R.N. Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Viruses 2019, 11, 755. https://doi.org/10.3390/v11080755
Thirion L, Pezzi L, Corcostegui I, Dubot-Pérès A, Falchi A, de Lamballerie X, Charrel RN. Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Viruses. 2019; 11(8):755. https://doi.org/10.3390/v11080755
Chicago/Turabian StyleThirion, Laurence, Laura Pezzi, Iban Corcostegui, Audrey Dubot-Pérès, Alessandra Falchi, Xavier de Lamballerie, and Remi N. Charrel. 2019. "Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome" Viruses 11, no. 8: 755. https://doi.org/10.3390/v11080755
APA StyleThirion, L., Pezzi, L., Corcostegui, I., Dubot-Pérès, A., Falchi, A., de Lamballerie, X., & Charrel, R. N. (2019). Development and Evaluation of a Duo Chikungunya Virus Real-Time RT-PCR Assay Targeting Two Regions within the Genome. Viruses, 11(8), 755. https://doi.org/10.3390/v11080755