Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Cases
2.2. Sample Preparation and Quantification of PRRSV RNA
2.3. Serology
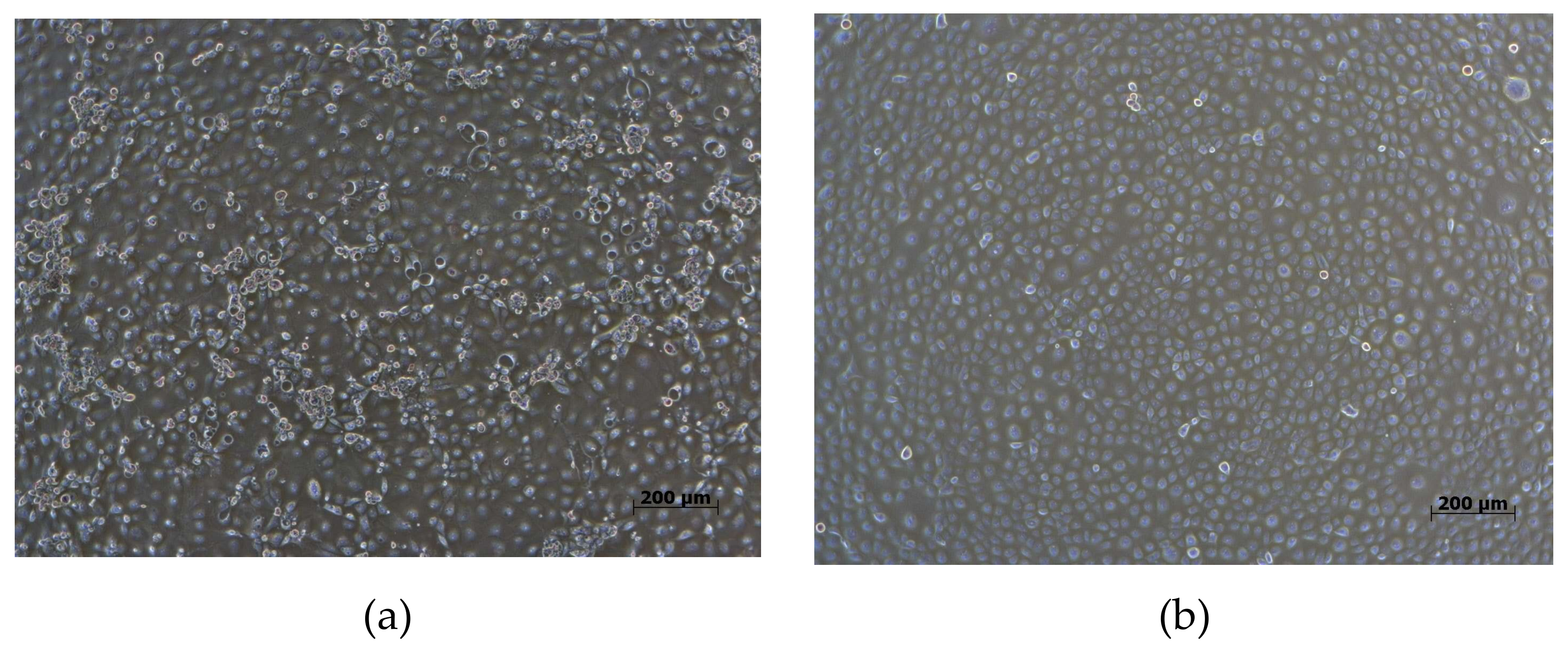
2.4. Virus Isolation and Whole Genome Sequencing of PRRSV
2.5. Open Reading Frame 5 Amplification and Sequencing
2.6. Sequence and Phylogenetic Analysis
3. Results
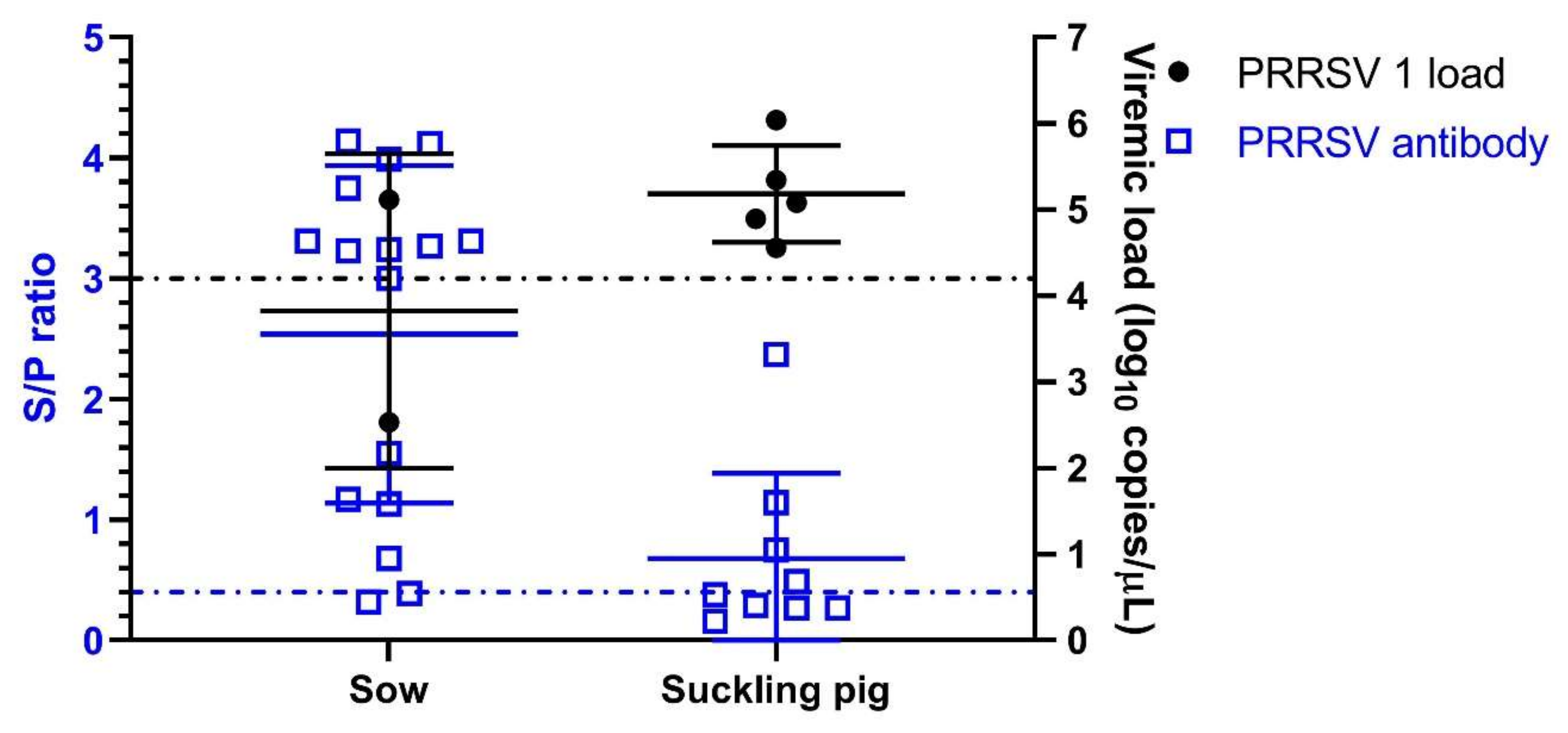
3.1. Emergence of PRRSV 1
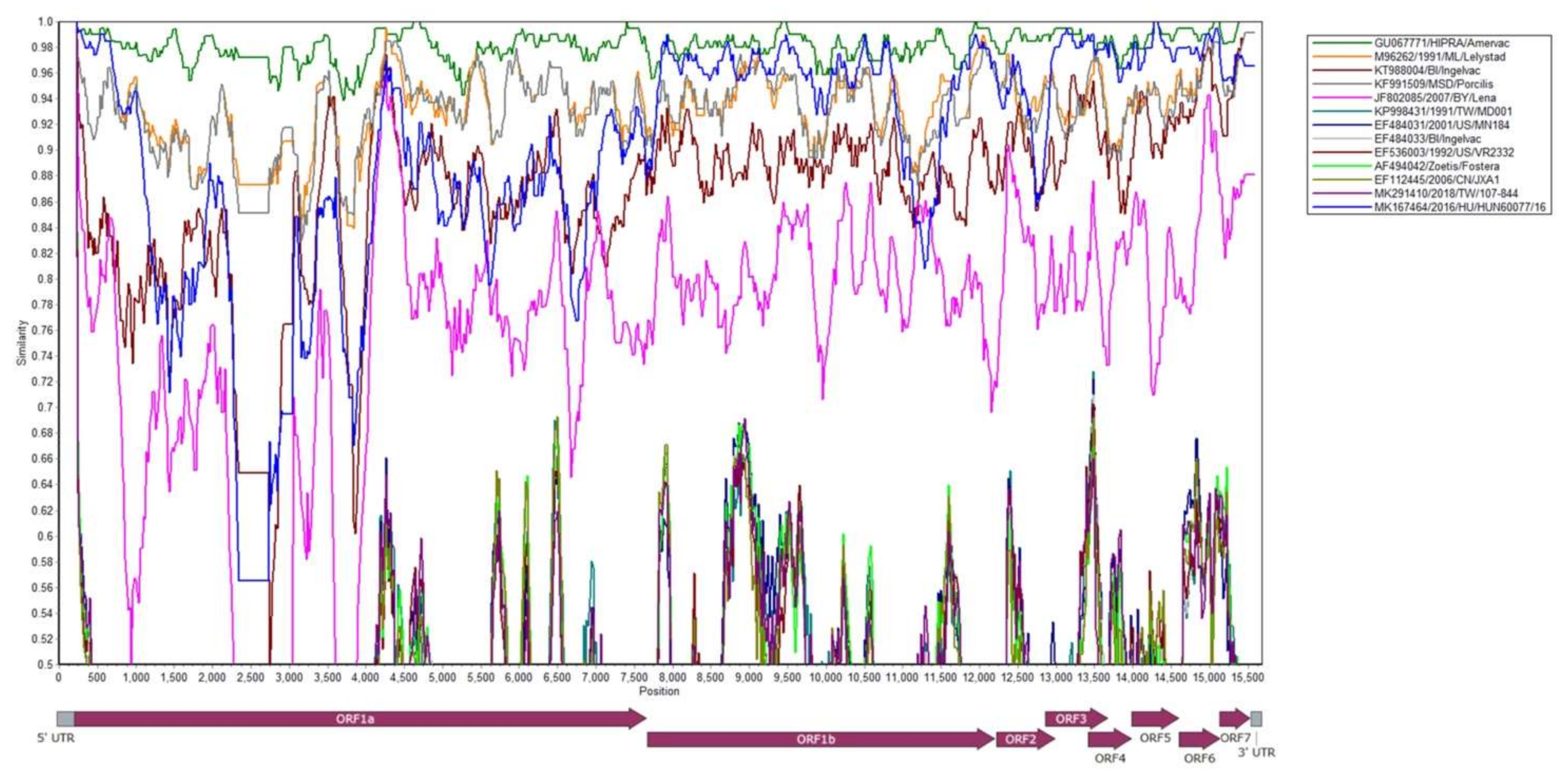
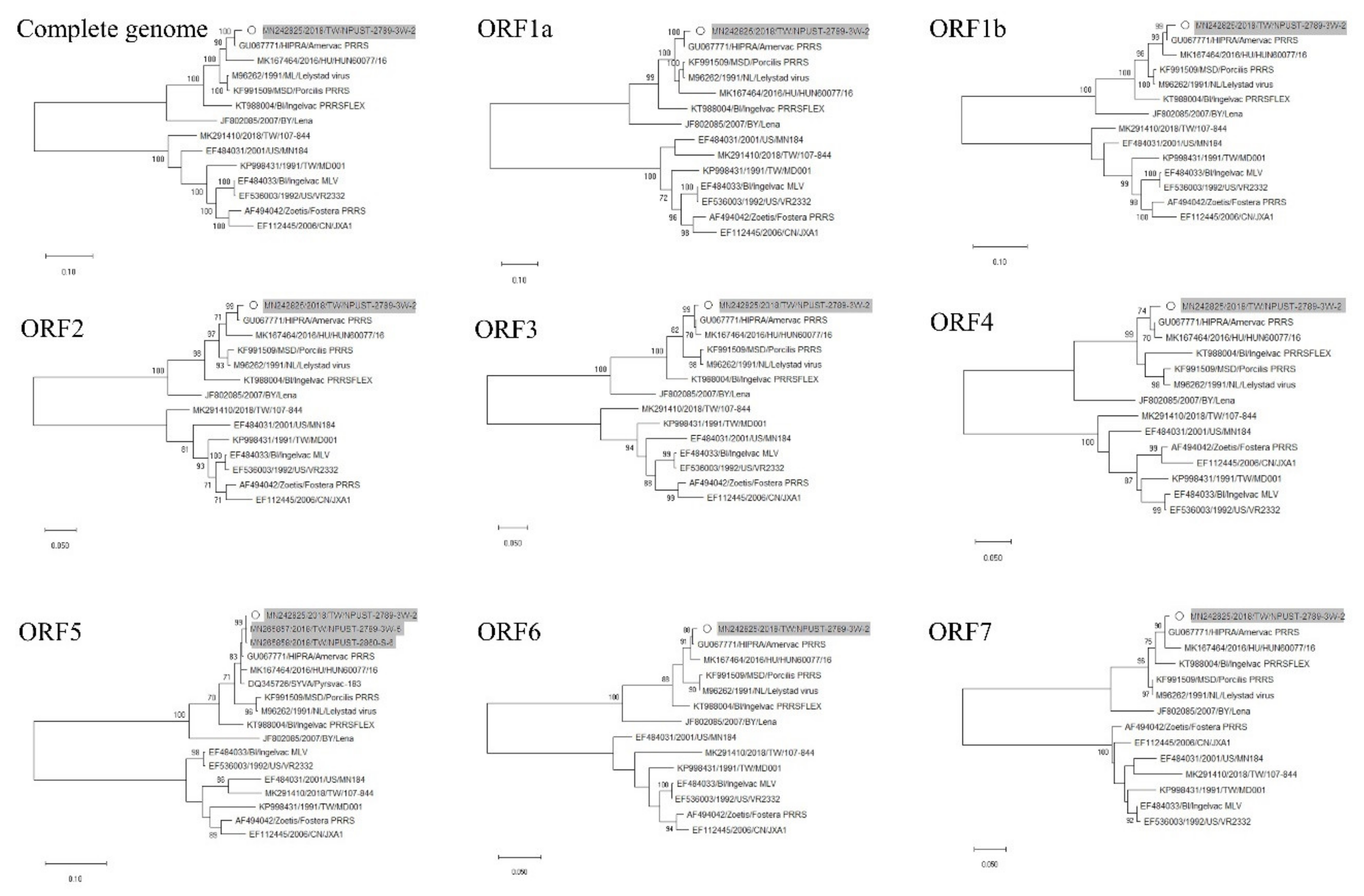
3.2. Phylogenetic Analysis of Whole Genome Sequences
3.3. Phylogenetic, Amino Acid and Nucleotide Sequence Analysis of ORF5
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; McCullough, S.; Morrison, R.B.; Joo, H.S.; et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Investig. 1992, 4, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Carman, S.; Sanford, S.E.; Dea, S. Assessment of seropositivity to porcine reproductive and respiratory syndrome (PRRS) virus in swine herds in Ontario-1978 to 1982. Can. Vet. J. 1995, 36, 776–777. [Google Scholar] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Elam, M.R.; Kakach, L.T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch. Virol. 1995, 140, 1451–1460. [Google Scholar] [CrossRef]
- Nelsen, C.J.; Murtaugh, M.P.; Faaberg, K.S. Porcine reproductive and respiratory syndrome virus comparison: Divergent evolution on two continents. J. Virol. 1999, 73, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Wensvoort, G.; de Kluyver, E.P.; Luijtze, E.A.; den Besten, A.; Harris, L.; Collins, J.E.; Christianson, W.T.; Chladek, D. Antigenic comparison of Lelystad virus and swine infertility and respiratory syndrome (SIRS) virus. J. Vet. Diagn. Investig. 1992, 4, 134–138. [Google Scholar] [CrossRef]
- Zimmerman, J.J.; Dee, S.A.; Holtkamp, D.J.; Murtaugh, M.P.; Stadejek, T.; Stevenson, G.W.; Torremorell, M.; Yang, H.; Zhang, J. Porcine Reproductive and Respiratory Syndrome Viruses (Porcine Arteriviruses). In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 685–708. [Google Scholar]
- Murtaugh, M.P.; Genzow, M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS). Vaccine 2011, 29, 8192–8204. [Google Scholar] [CrossRef]
- Nielsen, H.S.; Oleksiewicz, M.B.; Forsberg, R.; Stadejek, T.; Botner, A.; Storgaard, T. Reversion of a live porcine reproductive and respiratory syndrome virus vaccine investigated by parallel mutations. J. Gen. Virol. 2001, 82, 1263–1272. [Google Scholar] [CrossRef]
- Marton, S.; Szalay, D.; Kecskemeti, S.; Forro, B.; Olasz, F.; Zadori, Z.; Szabo, I.; Molnar, T.; Banyai, K.; Balint, A. Coding-complete sequence of a vaccine-derived recombinant porcine reproductive and respiratory syndrome virus strain isolated in Hungary. Arch. Virol. 2019, 164, 2605–2608. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Chen, Q.; Wang, L.; Madson, D.; Harmon, K.; Gauger, P.; Zhang, J.; Li, G. Recombination between Vaccine and Field Strains of Porcine Reproductive and Respiratory Syndrome Virus. Emerg. Infect. Dis. 2019, 25, 2335–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Kang, R.; Yu, J.; Xie, B.; Chen, C.; Li, X.; Xie, J.; Ye, Y.; Xiao, L.; Zhang, J.; et al. Genetic Characterization and Pathogenicity of a Novel Recombined Porcine Reproductive and Respiratory Syndrome Virus 2 among Nadc30-Like, Jxa1-Like, and Mlv-Like Strains. Viruses 2018, 10, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eclercy, J.; Renson, P.; Lebret, A.; Hirchaud, E.; Normand, V.; Andraud, M.; Paboeuf, F.; Blanchard, Y.; Rose, N.; Bourry, O. A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs. Viruses 2019, 11, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dooren, K. Denmark: New PRRS strain emerged. Pig Progress, 16 November 2019. Available online: https://www.pigprogress.net/Piglets/Articles/2019/11/Denmark-New-PRRS-strain-emerged-in-July-499756E/.
- Chen, N.; Cao, Z.; Yu, X.; Deng, X.; Zhao, T.; Wang, L.; Liu, Q.; Li, X.; Tian, K. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J. Gen. Virol. 2011, 92, 880–892. [Google Scholar] [CrossRef]
- Iseki, H.; Takagi, M.; Kawashima, K.; Shibahara, T.; Kuroda, Y.; Tsunemitsu, H.; Yamakawa, M. Type 1 porcine reproductive and respiratory syndrome virus emerged in Japan. In Proceedings of the 22nd International Pig Veterinary Society Congress, Jeju, Korea, 10–13 June 2012; p. 978. [Google Scholar]
- Kim, S.H.; Roh, I.S.; Choi, E.J.; Lee, C.; Lee, C.H.; Lee, K.H.; Lee, K.K.; Song, Y.K.; Lee, O.S.; Park, C.K. A molecular analysis of European porcine reproductive and respiratory syndrome virus isolated in South Korea. Vet. Microbiol. 2010, 143, 394–400. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Vuong, T.H.V.; Vo, T.H. PRRS in Vietnam and its diagnosis. J. Life Sci. 2015, 9, 272–276. [Google Scholar]
- Thanawongnuwech, R.; Amonsin, A.; Tatsanakit, A.; Damrongwatanapokin, S. Genetics and geographical variation of porcine reproductive and respiratory syndrome virus (PRRSV) in Thailand. Vet. Microbiol. 2004, 101, 9–21. [Google Scholar] [CrossRef]
- Yoshii, M.; Kaku, Y.; Murakami, Y.; Shimizu, M.; Kato, K.; Ikeda, H. Polymerase chain reaction-based genetic typing of Japanese porcine reproductive and respiratory syndrome viruses. J. Vet. Diagn. Investig. 2004, 16, 342–347. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Yang, H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010, 154, 31–37. [Google Scholar] [CrossRef]
- Wu, H.-H.; Lin, W.-H.; Su, G.-S.; Lin, C.-N.; Chiou, M.-T. Comparison of viremia of type I and II porcine reproductive and respiratory syndrome virus in Taiwan. In 24th International Pig Veterinary Society Congress; Kirwan, P., Ed.; Pat Kirwan: Dublin, Ireland, 2016; p. 547. [Google Scholar]
- Holtkamp, D.J.; Polson, D.D.; Torremorell, M.; Morrison, B.; Classen, D.M.; Becton, L.; Henry, S.; Rodibaugh, M.T.; Rowland, R.R.; Snelson, H.; et al. Terminology for classifying the porcine reproductive and respiratory syndrome virus (PRRSV) status of swine herds. Tierarztl Prax Ausg G Grosstiere Nutztiere 2011, 39, 101–112. [Google Scholar]
- Lin, C.N.; Lin, W.H.; Hung, L.N.; Wang, S.Y.; Chiou, M.T. Comparison of viremia of type II porcine reproductive and respiratory syndrome virus in naturally infected pigs by zip nucleic acid probe-based real-time PCR. BMC Vet. Res. 2013, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.H.; Shih, H.C.; Wang, S.Y.; Lin, C.F.; Yang, C.Y.; Chiou, M.T.; Lin, C.N. Emergence of a virulent porcine reproductive and respiratory syndrome virus in Taiwan in 2018. Transbound. Emerg. Dis. 2019, 66, 1138–1141. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, K.G.; Hansen, C.M.; Madsen, E.S.; Strandbygaard, B.; Botner, A.; Sorensen, K.J. Sequence analysis of porcine reproductive and respiratory syndrome virus of the American type collected from Danish swine herds. Arch. Virol. 1998, 143, 1683–1700. [Google Scholar] [CrossRef]
- Chen, N.; Liu, Q.; Qiao, M.; Deng, X.; Chen, X.; Sun, M. Whole genome characterization of a novel porcine reproductive and respiratory syndrome virus 1 isolate: Genetic evidence for recombination between Amervac vaccine and circulating strains in mainland China. Infect. Genet. Evol. 2017, 54, 308–313. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Species | Year | Country/Company | Sequence Identity (%) | GenBank Accession Number | |
|---|---|---|---|---|---|---|
| Whole Genome | ORF5 | |||||
| Amervac PRRS 1 | PRRSV 1 | N/A | HIPRA | 98.2 | 99.0 | GU067771 |
| Porcilis PRRS | PRRSV 1 | N/A | MSD | 93.8 | 94.2 | KF991509 |
| Ingelvac PRRSFLEX | PRRSV 1 | N/A | Boehringer Ingelheim | 87.4 | 92.6 | KT988004 |
| Pyrsvac-183 1 | PRRSV 1 | N/A | SYVA | N/A | 98.5 | DQ345726 |
| Ingelvac PRRS 2 | PRRSV 2 | N/A | Boehringer Ingelheim | 57.8 | 59.7 | EF484033 |
| Fostera PRRS 2 | PRRSV 2 | N/A | Zoetis | 57.5 | 59.2 | AF494042 |
| Lelystad virus | PRRSV 1 | 1991 | Netherlands | 94.1 | 94.5 | M96262 |
| Lena | PRRSV 1 | 2007 | Belarus | 80.2 | 82.8 | JF802085 |
| HUN60077/16 | PRRSV 1 | 2016 | Hungary | 91.7 | 98.2 | MK167464 |
| NPUST-2789-3W-2 | PRRSV 1 | 2018 | Taiwan | 100 | 100 | MN242825 |
| NPUST-2789-3W-5 | PRRSV 1 | 2018 | Taiwan | N/A | 99.8 | MN265857 |
| NPUST-2860-S-6 | PRRSV 1 | 2018 | Taiwan | N/A | 99.8 | MN265858 |
| VR2332 | PRRSV 2 | 1992 | USA | 57 | 59.9 | EF536003 |
| MD001 | PRRSV 2 | 1991 | Taiwan | 56.1 | 58.4 | KP998431 |
| MN184 | PRRSV 2 | 2001 | USA | 57.9 | 58.4 | EF484031 |
| JXA1 | PRRSV 2 | 2006 | China | 57.3 | 60.7 | EF112445 |
| 107-844 | PRRSV 2 | 2018 | Taiwan | 57.8 | 56.8 | MK291410 |
| ORF | Amervac PRRS | Lelystad Virus | HUN60077/16 | |||
|---|---|---|---|---|---|---|
| Nucleotide (%) | Amino Acid (%) | Nucleotide (%) | Amino Acid (%) | Nucleotide (%) | Amino Acid (%) | |
| ORF1a | 98.1 | 98.1 | 93.6 | 94.4 | 87.2 | 87.6 |
| ORF1b | 98.2 | 99.1 | 94.3 | 98.2 | 95.4 | 98.6 |
| ORF2 | 98.7 | 97.7 | 94.9 | 94.6 | 93.6 | 94.6 |
| ORF3 | 98.9 | 99.6 | 94.5 | 94.4 | 98 | 98.5 |
| ORF4 | 97.8 | 96.8 | 93.3 | 94.1 | 96.9 | 94.6 |
| ORF5 | 99.8 | 100 | 98.5 | 97.1 | 98.2 | 97.1 |
| ORF6 | 99.2 | 98.3 | 96.4 | 96.6 | 98.1 | 97.1 |
| ORF7 | 99.2 | 98.5 | 96.2 | 96.2 | 96.7 | 96.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-H.; Kaewprom, K.; Wang, S.-Y.; Lin, C.-F.; Yang, C.-Y.; Chiou, M.-T.; Lin, C.-N. Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan. Viruses 2020, 12, 316. https://doi.org/10.3390/v12030316
Lin W-H, Kaewprom K, Wang S-Y, Lin C-F, Yang C-Y, Chiou M-T, Lin C-N. Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan. Viruses. 2020; 12(3):316. https://doi.org/10.3390/v12030316
Chicago/Turabian StyleLin, Wei-Hao, Kraijak Kaewprom, Sheng-Yuan Wang, Chuen-Fu Lin, Cheng-Yao Yang, Ming-Tang Chiou, and Chao-Nan Lin. 2020. "Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan" Viruses 12, no. 3: 316. https://doi.org/10.3390/v12030316
APA StyleLin, W. -H., Kaewprom, K., Wang, S. -Y., Lin, C. -F., Yang, C. -Y., Chiou, M. -T., & Lin, C. -N. (2020). Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan. Viruses, 12(3), 316. https://doi.org/10.3390/v12030316