
Robust and Persistent B- and T-Cell Responses after COVID-19 in Immunocompetent and Solid Organ Transplant Recipient Patients

 , , , , ,
, , , , ,  , , ,
, , ,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Antibody Assays
2.3. Protein Peptide Pools
2.4. PBMC Isolation
2.5. Activation Induction Marker Assay
2.6. Antigen-Specific Cytokine Production
2.7. Detection of Antigen-Specific CD4+and CD8+ T-Cell Proliferative Response
2.8. Statistical Analysis
3. Results
3.1. Characteristics of the Patients Analysed
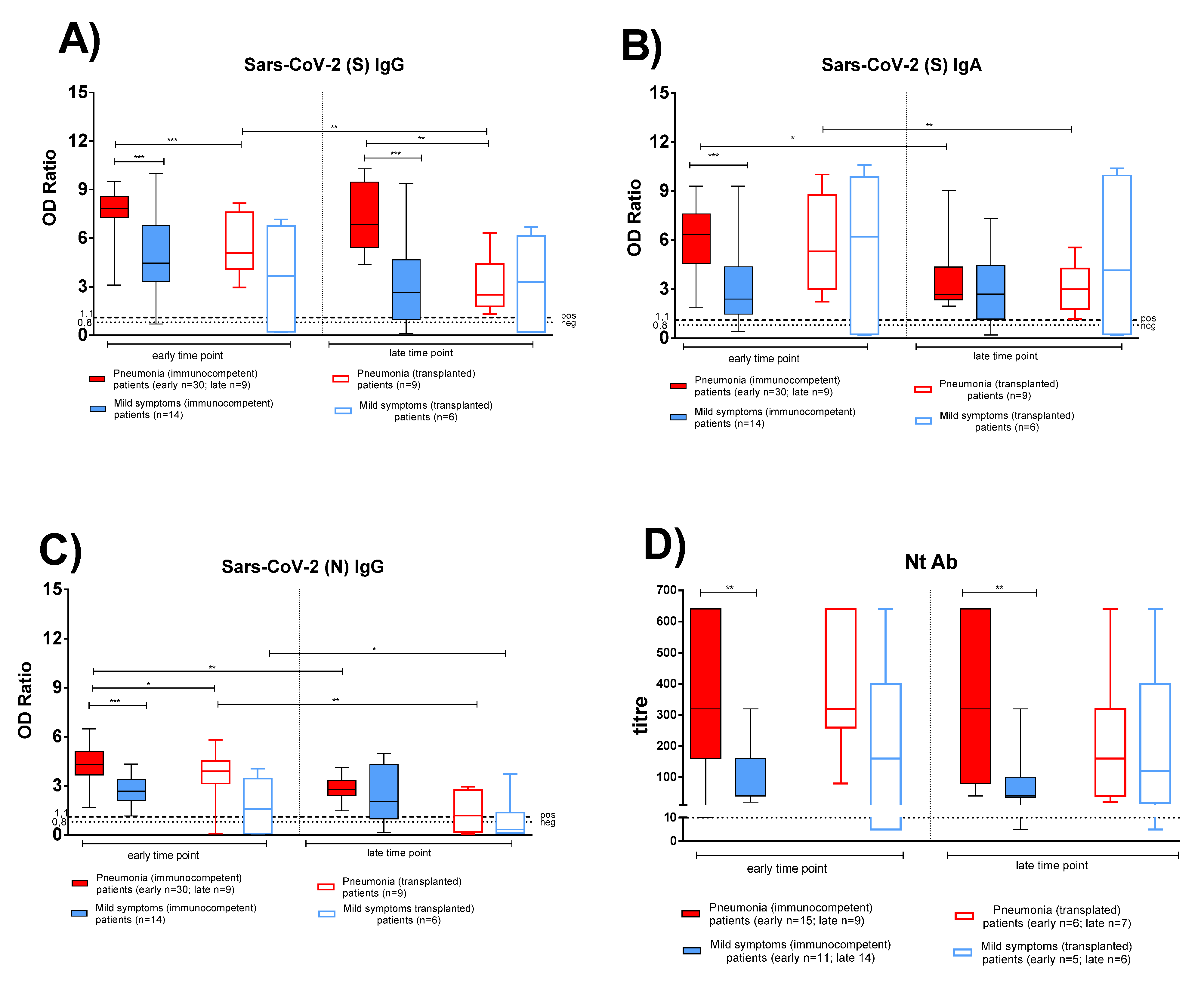
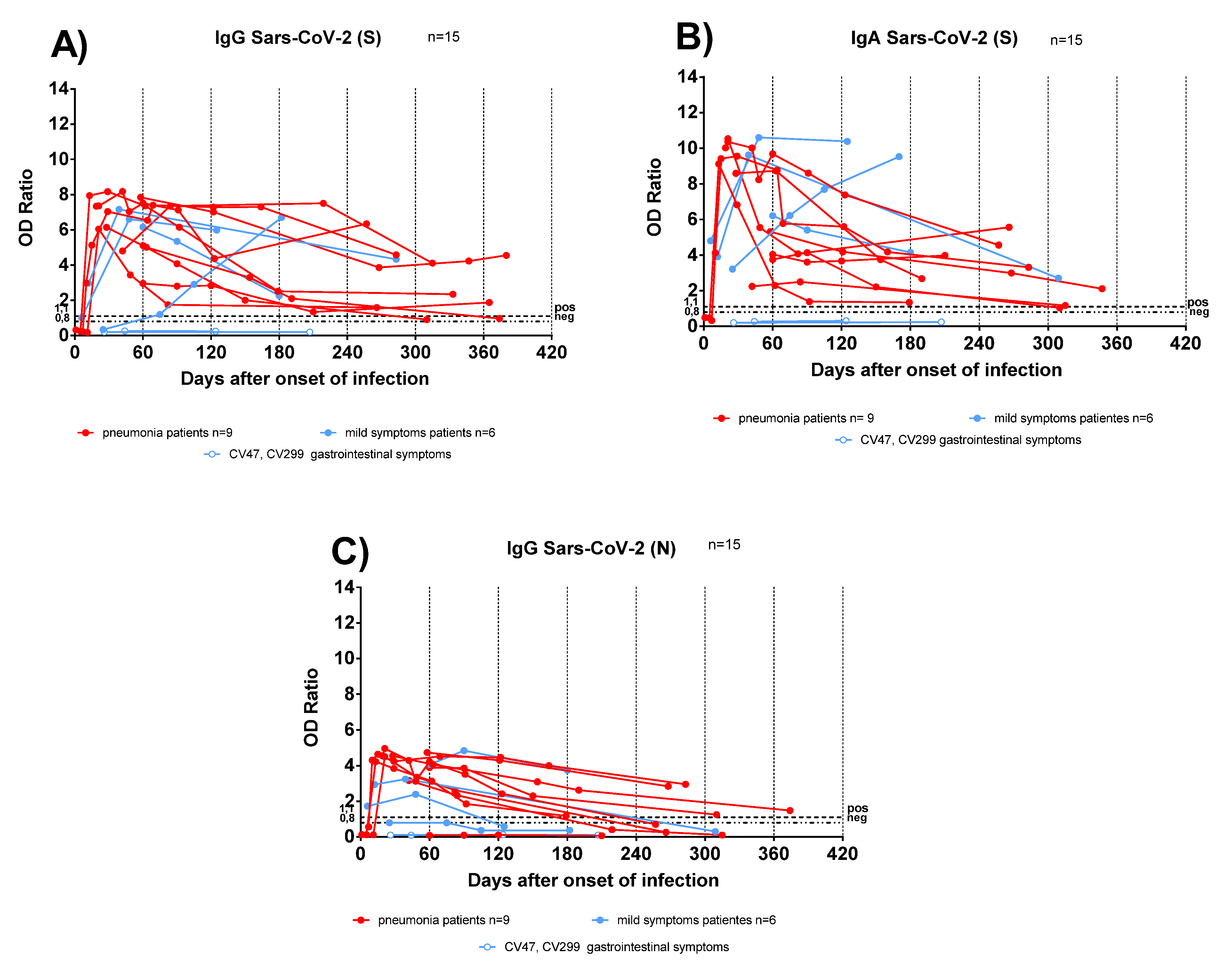
3.2. Antigen-Specific Antibody Response in Convalescent COVID-19 IC and TX Patients
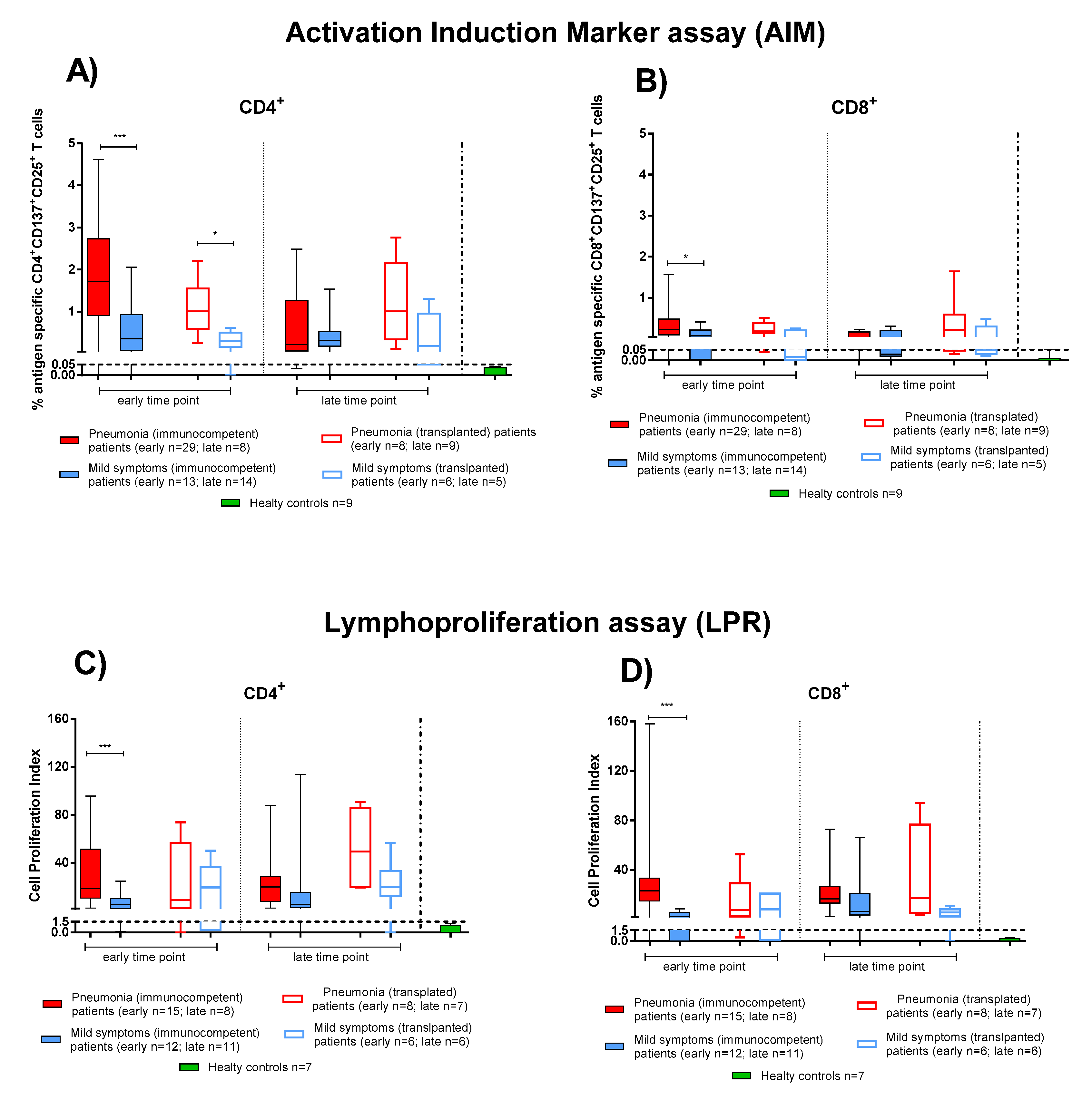
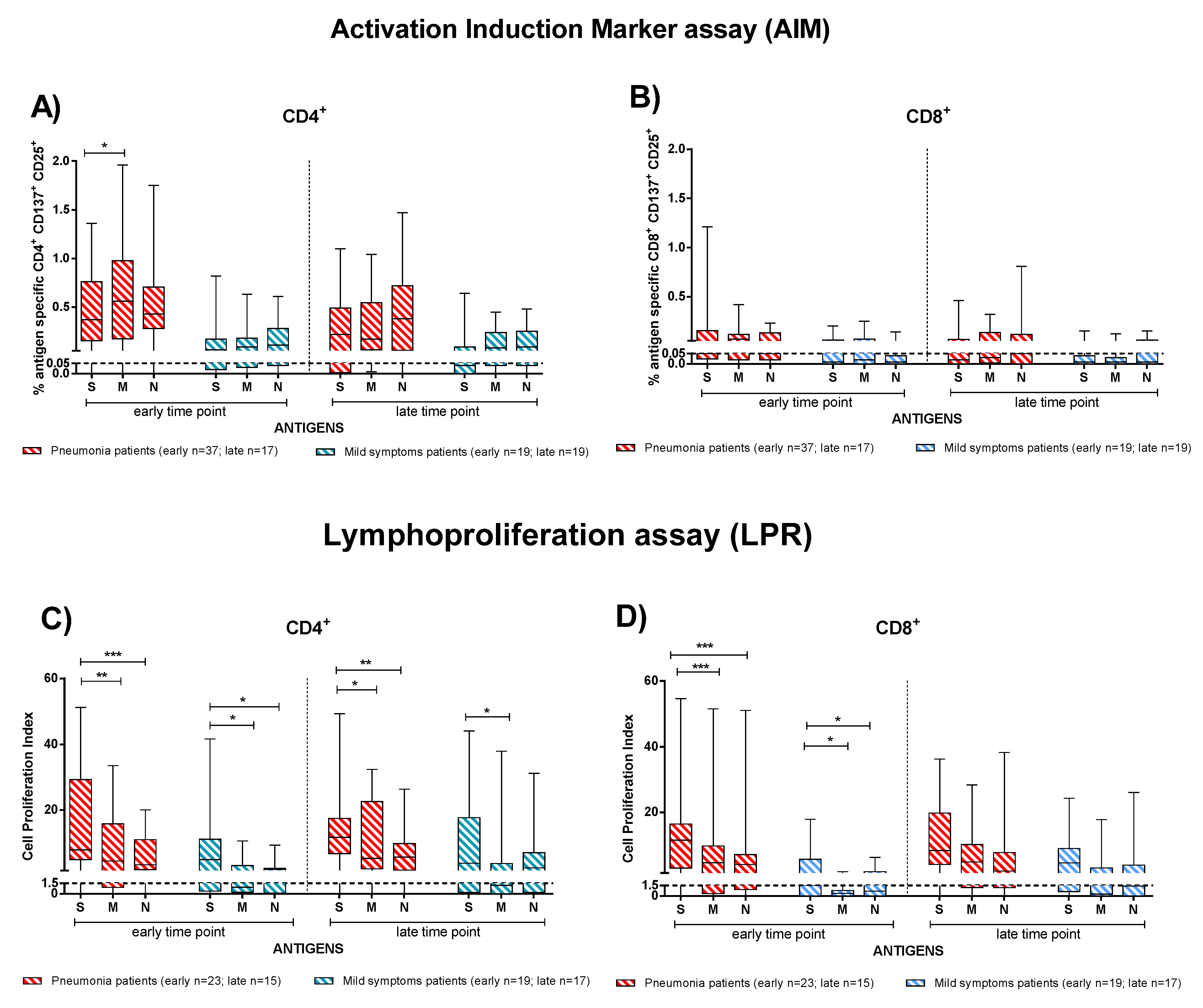
3.3. Effector and Memory T-Cell Responses in IC and TX Patients after COVID-19
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chan, J.F.; Yuan, S.; Kok, K.H.; Kai-Wang To, K.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Chik-Yan Yip, C.; Wing-Shan Poon, R.; et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [Green Version]
- Candel González, F.J.; Viñuela-Prieto, J.M.; Del Castillo, J.G.; García, P.B.; Saavedra, M.F.; Píriz, A.H.; Jiménez Virumbrales, D.; Canora Lebrato, J.; García de Casasola, G.; Gil Prieto, R.; et al. Utility of lateral flow tests in SARS-CoV-2 infection monitorization. Rev. Esp. Quimioter. 2020, 33, 258. [Google Scholar] [CrossRef]
- Carsetti, R.; Zaffina, S.; Mortari, E.P.; Terreri, S.; Corrente, F.; Capponi, C.; Palomba, P.; Mirabella, M.; Cascioli, S.; Palange, P.; et al. Different innate and adaptive immune response to SARS-CoV-2 infection of asymptomatic, mild and severe cases. Front. Immunol. 2020, 11, 610300. [Google Scholar] [CrossRef] [PubMed]
- Borremans, B.; Gamble, A.; Prager, K.; Helman, S.; McClain, A.; Cox, C.; Savage, V.; Lloyd-Smith, J.O. Quantifying antibody kinetics and RNA shedding during early-phase SARS-CoV-2 infection by time since symptom onset. eLife 2020, 9, e60122. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Tan, M.; He, H.; Tan, H.; Mai, J.; Long, Y.; Jiang, X.; He, Q.; Huang, Y.; Li, Y.; et al. Study on the expression levels of antibodies against SARS-CoV-2 at different period of disease and its related factors in 192 cases of COVID-19 patients. MedRxiv 2020. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of immune response in patients with (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.T.; Garcia-Carreras, B.; Hitchings, M.D.T.; Yang, B.; Katzelnick, L.; Rattigan, S.M.; Borgert, B.A.; Moreno, C.A.; Solomon, B.D.; Rodriguez-Barraquer, I.; et al. A systematic review of antibody mediated immunity to coronaviruses: Antibody kinetics, correlates of protection, and association of antibody responses with severity of disease. Nat. Commun. 2020, 11, 4704. [Google Scholar] [CrossRef]
- Zhang, J.; Qu, X.; Liu, Z.; Wang, Q.; Wu, J.; Hu, Y.; Bai, T.; Xie, T.; Huang, M.; Wu, T.; et al. Spike-specific circulating T follicular helper cell and cross-neutralizing antibody responses in COVID-19 convalescent individuals. Nat. Microbiol. 2021, 6, 51–58. [Google Scholar] [CrossRef]
- Fafi-Kremer, S.; Bruel, T.; Madec, Y.; Grant, R.; Tondeur, L.; Grzelak, L.; Staropoli, I.; Anna, F.; Souque, P.; Fernandes-Pellerin, S.; et al. Serologic responses to SARSCoV-2 infection among hospital staff with mild disease in eastern France. medRxiv 2020. [Google Scholar] [CrossRef]
- Brochot, E.; Demey, B.; Touze, A.; Belouzard, S.; Dubuisson, J.; Schmit, J.L.; Duverlie, G.; Francois, C.; Castelain, S.; Helle, F.; et al. Anti-Spike anti-Nucleocapsid and neutralizing antibodies in SARS-CoV-2 inpatients and asymptomatic individuals. Front. Microbiol. 2020, 11, 584251. [Google Scholar] [CrossRef]
- Wang, X.; Guo, X.; Xin, Q.; Pan, Y.; Hu, Y.; Li, J.; Chu, Y.; Feng, Y.; Wang, Q. Neutralizing Antibody Responses to Severe Acute Respiratory Syndrome Coronavirus 2 in Coronavirus Disease 2019 Inpatients and Convalescent Patients. Clin. Infect. Dis. 2020, 71, 2688–2694. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, L.; Nair, M.S.; Yin, M.T.; Luo, Y.; Wang, Q.; Yuan, T.; Mori, K.; Guzman Solis, A.; Yamashita, M.; et al. SARS-CoV-2 neutralizing antibody responses are more robust in patients with severe disease. Emerg. Microbes Infect. 2020, 9, 2091–2093. [Google Scholar] [CrossRef]
- Thieme, C.J.; Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seiberto, F.S.; Hoelzer, B.; Konik, M.J.; Moritz Berger, M.; Brenner, T.; et al. Robust T cell response towards spike, membrane, and nucleocapsid SARS-CoV-2proteins is not associated with recovery in critical COVID-19 patients. Cell Rep. Med. 2020, 1, 100092. [Google Scholar] [CrossRef]
- Weiskopf, D.; Schmitz, K.S.; Raadsen, M.P.; Grifoni, A.; Okba, N.M.A.; Henrik Endeman, H.; van den Akker, J.P.C.; Molenkamp, R.; Koopmans, M.P.G.; van Gorp, E.C.M.; et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci. Immunol. 2020, 5, eabd2071. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Rydyznski Moderbacher, C.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Marcos, A.; Salcedo, M.; Alonso-Fernández, R.; Rodríguez-Perálvarez, M.; Olmedo, M.; Graus Morales, J.; Cuervas-Mons, V.; Cachero, A.; Loinaz-Segurola, C.; Iñarrairaegui, M.; et al. Changes in humoral immune response after SARS-CoV-2 infection in liver transplant recipients compared to immunocompetent patients. Am. J. Transplant. 2021, 21, 2876–2884. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seibert, F.S.; Hoelzer, B.; Konik, M.J.; Meister, T.L.; Pfaender, S.; et al. The Magnitude and Functionality of SARS-CoV-2 Reactive Cellular and Humoral Immunity in Transplant Population Is Similar to the General Population Despite Immunosuppression. Transplantation 2021, 105, 2156. [Google Scholar] [CrossRef]
- Percivalle, E.; Cambiè, G.; Cassaniti, I.; Vecchio Nepita, E.; Maserati, R.; Ferrari, A.; Di Martino, R.; Isernia, P.; Mojoli, F.; Bruno, R.; et al. Prevalence of SARS-CoV-2 specific neutralising antibodies in blood donors from the Lodi Red Zone in Lombardy, Italy, as at 06 April 2020. Eurosurveill 2020, 25, 2001031. [Google Scholar] [CrossRef] [PubMed]
- Li, C.K.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.J.; Weiss, R.A.; Brenchley, J.M.; et al. T cell responses to whole SARS coronavirus in humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favà, A.; Donadeu, L.; Sabé, N.; Pernin, V.; González-Costello, J.; Lladó, L.; Meneghini, M.; Charmetant, X.; García-Romero, E.; Cachero, A.; et al. SARS-CoV-2-Specific serological and functional T cell immune responses during acute and early COVID-19 convalescence in solid organ transplant patients. Am. J. Transplant. 2021, 21, 2749–2761. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P.; et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Favà, A.; Cucchiari, D.; Montero, N.; Toapanta, N.; Centellas, F.J.; Vila-Santandreu, A.; Coloma, A.; Meneghini, M.; Manonelles, A.; Sellarés, J.; et al. Clinical characteristics and risk factors for severe COVID-19 in hospitalized kidney transplant recipients: A multicentric cohort study. Am. J. Transplant. 2020, 20, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Alberici, F.; Delbarba, E.; Manenti, C.; Econimo, L.; Francesca Valerio, F.; Pola, A.; Maffei, C.; Possenti, S.; Zambetti, N.; Moscato, M.; et al. A single center observational study of the clinical characteristics and short-term outcome of 20 kidney transplant patients admitted for SARS-CoV2 pneumonia. Kidney Int. 2020, 97, 1083–1088. [Google Scholar] [CrossRef]
- Coll, E.; Fernandez-Ruiz, M.; Padilla, M.; Moreso, F.; Hernández-Vicente, A.; Yañez, I.; Molina, M.; Vázquez-Sánchez, T.; Crespo, M.; Facund, C.; et al. COVID-19 in Solid Organ Transplant Recipients in Spain throughout 2020: Catching the Wave? Transplantation 2021, 105, 2146. [Google Scholar] [CrossRef]
- Grupper, A.; Rabinowich, L.; Schwartz, D.; Schwartz, I.F.; Ben-Yehoyada, M.; Shashar, M.; Katchman, E.; Halperin, T.; Turner, D.; Goykhman, Y.; et al. Reduced humoral response to mRNA SARS-CoV-2 BNT162b2 vaccine in kidney transplant recipients without prior exposure to the virus. Am. J. Transplant. 2021, 21, 2719–2726. [Google Scholar] [CrossRef]
- Marinaki, S.; Adamopoulos, S.; Degiannis, D.; Roussos, S.; Pavlopoulou, I.D.; Hatzakis, A.; Boletis, I.N. Immunogenicity of SARS-CoV-2 BNT162b2 vaccine in solid organ transplant recipients. Am. J. Transplant. 2021. [Google Scholar] [CrossRef]
- Bertrand, D.; Hamzaoui, M.; Lemée, V.; Lamulle, J.; Hanoy, M.; Laurent, C.; Lebourg, L.; Etienne, I.; Lemoine, M.; Le Roy, F.; et al. Antibody and T Cell Response to SARS-CoV-2 Messenger RNA BNT162b2 Vaccine in Kidney Transplant Recipients and Hemodialysis Patients. J. Am. Soc. Nephrol. 2021, 32, 2147–2152. [Google Scholar] [CrossRef]
- Kamar, N.; Abravanel, F.; Marion, O.; Couat, C.; Izopet, J.; Del Bello, A. Three Doses of an mRNA Covid-19 Vaccine in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2021, 385, 661–662. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, J.; Tonnus, W.; Paliege, A.; Rettig, R.; Steglich, A.; Gembardt, F.; Kessel, F.; Kröger, H.; Arndt, P.; Sradnick, J.; et al. Cellular and Humoral Immune Responses after Three Doses of BNT162b2 mRNA SARS-Cov-2 Vaccine in Kidney Transplant. Transplantation 2021, 105, e267–e269. [Google Scholar] [CrossRef] [PubMed]
- Cavagna, L.; Seminare, E.; Zanframundo, G.; Gregorini, M.; Di Matteo, A.; Rampino, T.; Montecucco, C.; Pelenghi, S.; Cattadori, B.; Pattonieri, E.F.; et al. Calcineurin Inhibitor-Based Immunosuppression and COVID-19: Results from a Multidisciplinary Cohort of Patients in Northern Italy. Microorganisms 2020, 8, 977. [Google Scholar] [CrossRef] [PubMed]
- Rovida, F.; Cassaniti, I.; Percivalle, E.; Sarasini, A.; Paolucci, S.; Klersy, C.; Cutti, S.; Novelli, V.; Marena, C.; Luzzaro, F.; et al. Incidence of SARS-CoV-2 infection in health care workers from Northern Italy based on antibody status: Immune protection from secondary infection—A retrospective observational case-control study. Int. J. Infect. Dis. 2021, 109, 199–202. [Google Scholar] [CrossRef]
- OMurchu, E.; Byrne, P.; Carty, P.G.; De Gascun, C.; Keogan, M.; O’Neill, M.; Harrington, P.; Ryan, M. Quantifying the risk of SARS-CoV-2 reinfection over time. Rev. Med. Virol. 2021, e2260. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunocompetent (IC) (n = 57) | p Value | Transplanted (TX) (n = 15) | p Value | p Value IC vs. TX | |||
|---|---|---|---|---|---|---|---|
| Pneumonia | Mild | Pneumonia | Mild | ||||
| Subjects % (n) | 53% (30) | 47% (27) | 60% (9) | 40% (6) | |||
| Transplanted Organ: | |||||||
| Kidney % (n) | na | na | 78% (7) | 33% (2) | |||
| Heart % (n) | na | na | 11% (1) | 33% (2) | |||
| Lung % (n) | na | na | 11% (1) | 33% (2) | |||
| Time After Transplant, Median [range] months | na | na | 89 [3–288] | 20 [1–43] | |||
| Age, Median [range] | 62 [44–81] | 45 [21–61] | p < 0.001 | 58 [48–71] | 59 [39–65] | p = 0.556 | p = 0.119 |
| Sex, M/F % (n) | 63% (19)/37% (11) | 48% (13)/52% (14) | p = 0.358 | 78% (7)/22% (2) | 83% (5)/18% (1) | p = 1 | p = 0.079 |
| Symptoms: | |||||||
| Fever % (n) | 90% (27) | 70% (19) | 100% (9) | 33% (2) | |||
| Rhinitis %(n) | 0 | 24% (8) | 0 | 0 | |||
| Cough % (n) | 43% (13) | 26% (7) | 50% (4) | 33% (2) | |||
| Sore Throat % (n) | 0 | 11% (3) | 13% (1) | 0 | |||
| Conjunctivitis % (n) | 0 | 0 | 13% (1) | 0 | |||
| Ageusia % (n) | 7% (2) | 56% (15) | 0 | 33% (2) | |||
| Anosmia % (n) | 3% (1) | 56% (15) | 0 | 0 | |||
| Gastrointestinal % (n) | 17% (5) | 19% (5) | 50% (4) | 50% (3) | |||
| Headache % (n) | 3% (1) | 44% (12) | 13% (1) | 33% (2) | |||
| O2 supply, % patients (n): | |||||||
| no | 3% (1) | 100% (27) | 56% (5) | 100% (6) | |||
| <5 mL/min | 27% (8) | 0 | 33% (3) | 0 | |||
| ≥5 mL/min | 70% (21) | 0 | 11% (1) | 0 | p < 0.001 | ||
| Duration of SARS-CoV-2 infection, Median [range] Days | 20 [4–38] | 20 [12–29] | 17 [10–36] | 7 [4–25] | p = 0.773 | ||
| Outcome: | |||||||
| Live % (n) | 100% (30) | 100% (27) | 100% (9) | 83% (5) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zavaglio, F.; Frangipane, V.; Morosini, M.; Gabanti, E.; Zelini, P.; Sammartino, J.C.; Ferrari, A.; Gregorini, M.; Rampino, T.; Asti, A.; et al. Robust and Persistent B- and T-Cell Responses after COVID-19 in Immunocompetent and Solid Organ Transplant Recipient Patients. Viruses 2021, 13, 2261. https://doi.org/10.3390/v13112261
Zavaglio F, Frangipane V, Morosini M, Gabanti E, Zelini P, Sammartino JC, Ferrari A, Gregorini M, Rampino T, Asti A, et al. Robust and Persistent B- and T-Cell Responses after COVID-19 in Immunocompetent and Solid Organ Transplant Recipient Patients. Viruses. 2021; 13(11):2261. https://doi.org/10.3390/v13112261
Chicago/Turabian StyleZavaglio, Federica, Vanessa Frangipane, Monica Morosini, Elisa Gabanti, Paola Zelini, Josè Camilla Sammartino, Alessandro Ferrari, Marilena Gregorini, Teresa Rampino, Annalia Asti, and et al. 2021. "Robust and Persistent B- and T-Cell Responses after COVID-19 in Immunocompetent and Solid Organ Transplant Recipient Patients" Viruses 13, no. 11: 2261. https://doi.org/10.3390/v13112261
APA StyleZavaglio, F., Frangipane, V., Morosini, M., Gabanti, E., Zelini, P., Sammartino, J. C., Ferrari, A., Gregorini, M., Rampino, T., Asti, A., Seminari, E., Di Matteo, A., Cattadori, B., Pellegrini, C., Tonello, S., Mallela, V. R., Minisini, R., Rizzi, M., Sainaghi, P. P., ... Baldanti, F. (2021). Robust and Persistent B- and T-Cell Responses after COVID-19 in Immunocompetent and Solid Organ Transplant Recipient Patients. Viruses, 13(11), 2261. https://doi.org/10.3390/v13112261