
Impact of Phage CDHS-1 on the Transcription, Physiology and Pathogenicity of a Clostridioides difficile Ribotype 027 Strain, R20291



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Culture Conditions
2.2. Phage CDHS-1 Propagation
2.3. One-Step Growth Curve of Phage CDHS-1
2.4. RNA Extraction and Analysis
2.5. RNA Sequencing and Transcript Isolation
2.6. Preparation of RT-qPCR Reactions
2.6.1. Construction of Complementary DNA (cDNA)
2.6.2. qPCR Primer Design and Reactions
2.7. Isolation of CDHS-1 Resistant and Lysogenic R20291 Strains
2.8. Characterization of the Virulence of CDHS-1 Resistant and Lysogenic R20291 Strains in G. mellonella
2.9. Assessment of Rate of Survival and Bacterial Colonization in G. mellonella Larvae
2.10. G. mellonella RNA Extraction, cDNA Synthesis and RT-qPCR of Growth and Infection Markers
3. Results
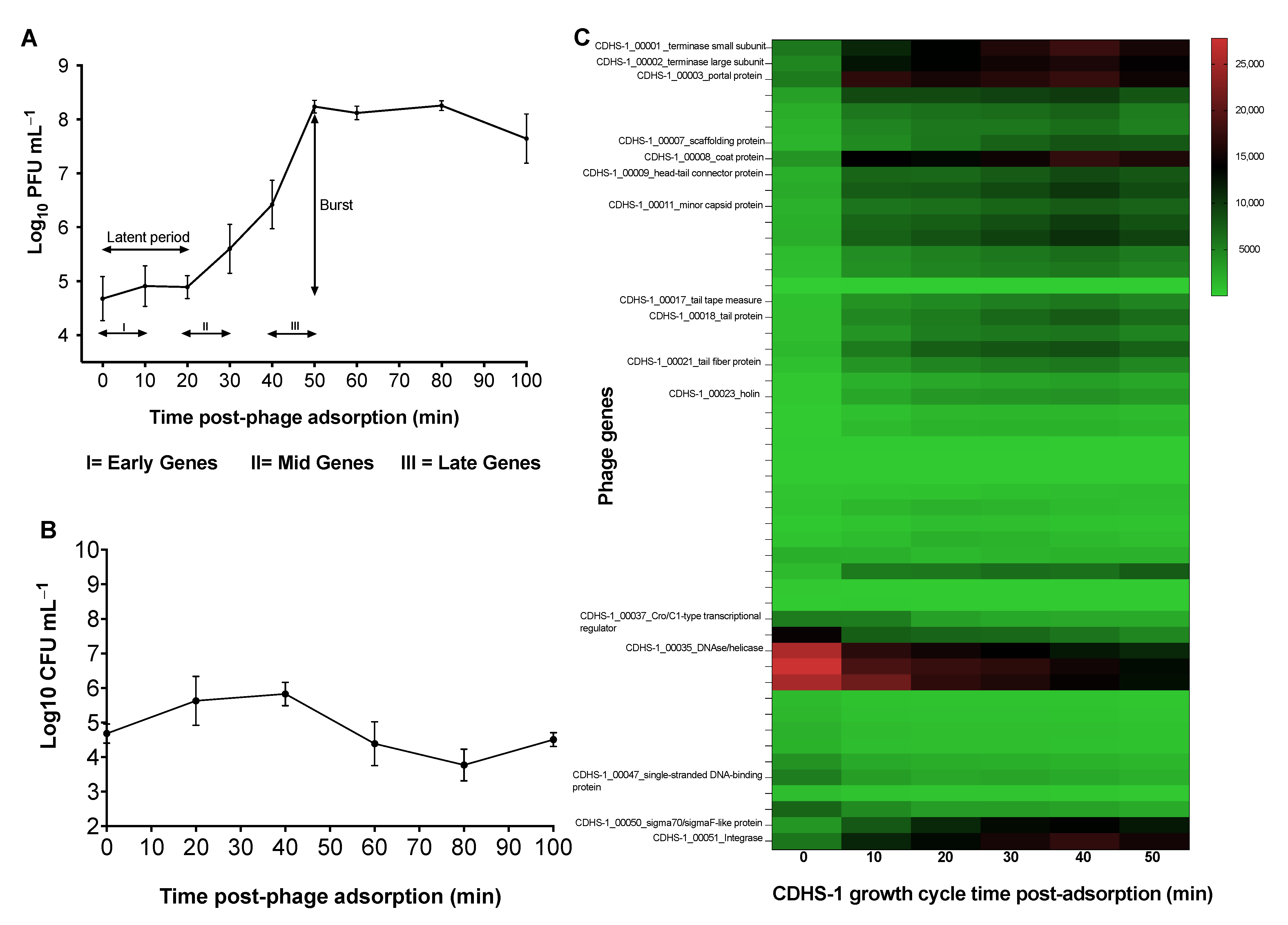
3.1. Growth Curve of Phage CDHS-1
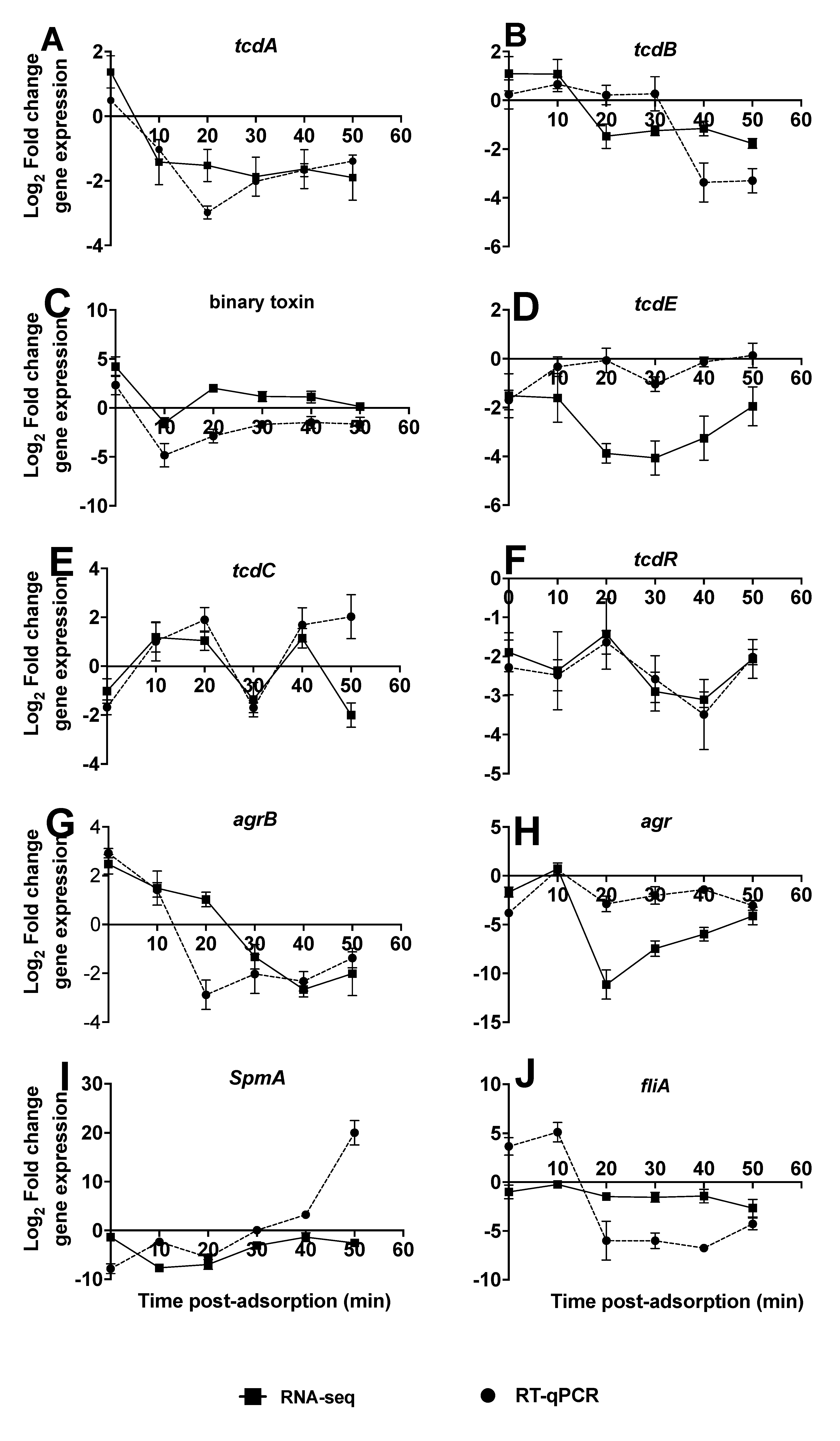
3.2. Transcriptional Changes in C. difficile R20291 during Infection with CDHS-1 Determined by RNA-seq
3.3. Regulations of Genes in CDHS-1 during R20291 Infection
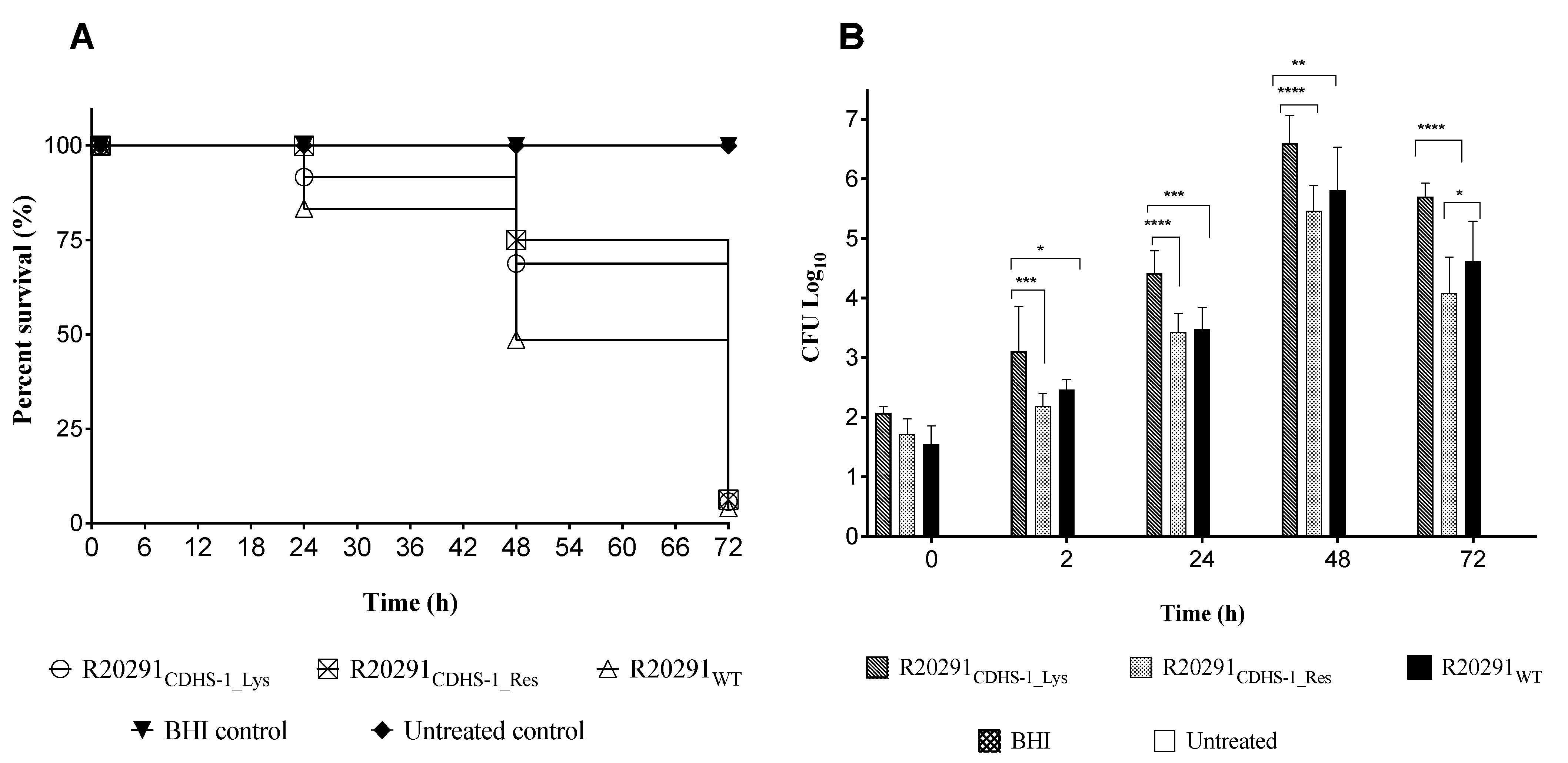
3.4. Impact of CDHS-1 Resistant and Lysogenic R20291 Strains on Survival Rates in G. mellonella
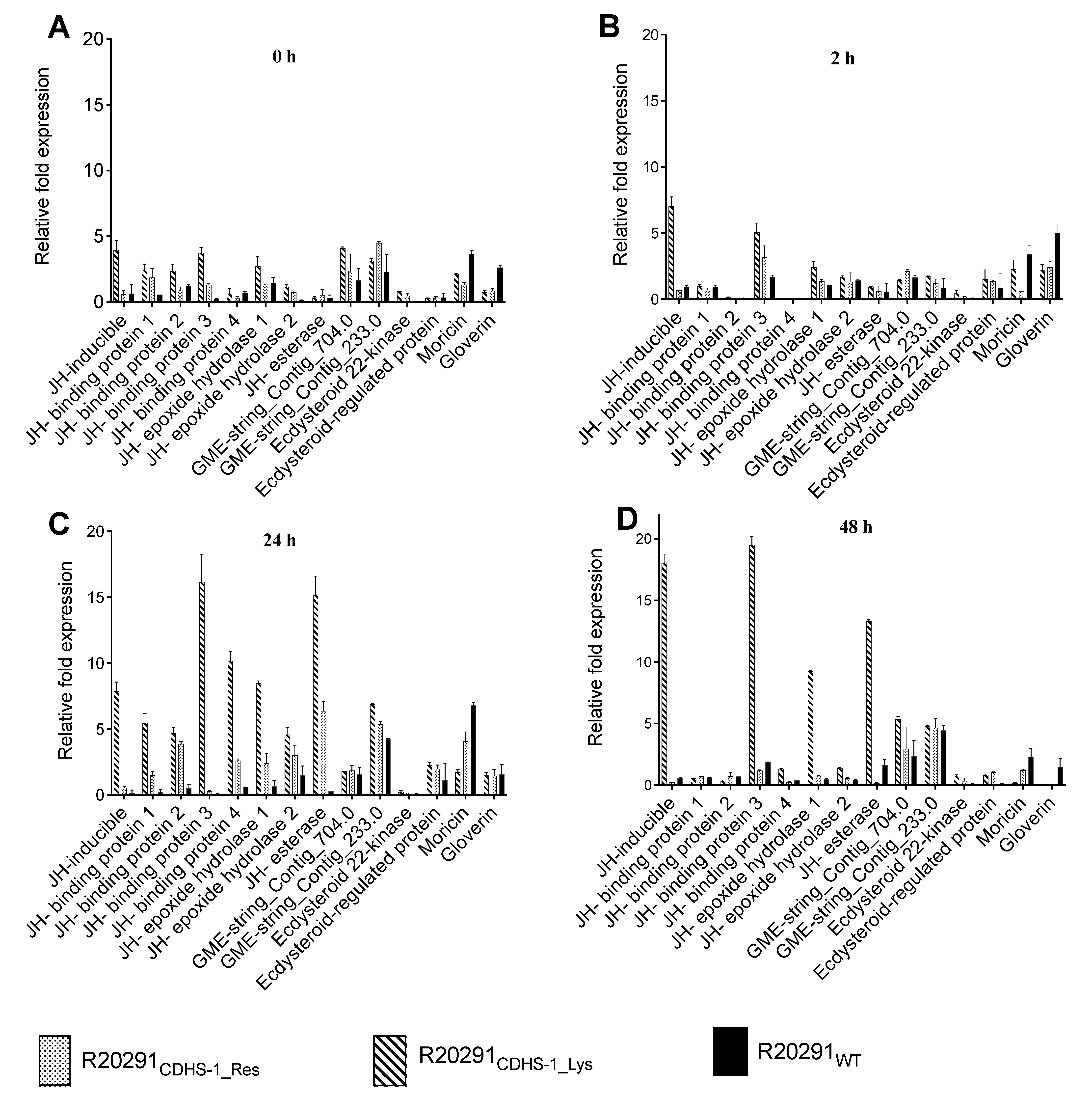
3.5. Expression Profiles of Growth, Reproduction and Infection Marker Genes in G. mellonella during Colonization with the CDHS-1 Resistant and Lysogenic R20291 Strains
4. Discussion
5. Conclusions and Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McFarland, L.V. Epidemiology, Risk Factors and Treatments for Antibiotic-Associated Diarrhea. Dig. Dis. 1998, 16, 292–307. [Google Scholar] [CrossRef]
- Kwon, J.H.; Olsen, M.A.; Dubberke, E.R. The Morbidity, Mortality, and Costs Associated with Clostridium difficile Infection. Infect. Dis. Clin. N. Am. 2015, 29, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium difficile colitis: Pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sun, C.; Wang, H.; Wang, J. The Role of Rho GTPases in Toxicity of Clostridium difficile Toxins. Toxins 2015, 7, 5254–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jank, T.; Giesemann, T.; Aktories, K. Rho-glucosylating Clostridium difficile toxins A and B: New insights into structure and function. Glycobiology 2007, 17, 15r–22r. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.H.; Tang, Y.J.; Silva, J.J. Analysis of the pathogenicity locus in Clostridium difficile strains. J. Infect. Dis. 2000, 181, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Haiko, J.; Westerlund-Wikström, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [Green Version]
- Borriello, S.P.; Davies, H.A.; Kamiya, S.; Reed, P.J.; Seddon, S. Virulence factors of Clostridium difficile. Rev. Infect. Dis. 1990, 12, 185–191. [Google Scholar] [CrossRef]
- He, M.; Sebaihia, M.; Lawley, T.D.; Stabler, R.A.; Dawson, L.F.; Martin, M.J.; Holt, K.E.; Seth-Smith, H.M.; Quail, M.A.; Rance, R.; et al. Evolutionary dynamics of Clostridium difficile over short and long time scales. Proc. Natl. Acad. Sci. USA 2010, 107, 7527–7532. [Google Scholar] [CrossRef] [Green Version]
- Goh, S.; Riley, T.V.; Chang, B.J. Isolation and Characterization of Temperate Bacteriophages of Clostridium difficile. Appl. Environ. Microbiol. 2005, 71, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- Nale, J.Y.; Spencer, J.; Hargreaves, K.R.; Buckley, A.M.; Trzepiński, P.; Douce, G.R.; Clokie, M.R.J. Bacteriophage Combinations Significantly Reduce Clostridium difficile Growth In Vitro and Proliferation In Vivo. Antimicrob. Agents Chemother. 2016, 60, 968–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, J.; Patel, P.V.; Hickenbotham, P.T.; Nale, J.Y.; Hargreaves, K.R.; Clokie, M.R.J. Prophage carriage and diversity within clinically relevant strains of Clostridium difficile. Appl. Environ. Microbiol. 2012, 78, 6027–6034. [Google Scholar] [CrossRef] [Green Version]
- Goh, S.; Hussain, H.; Chang, B.J.; Emmett, W.; Riley, T.V.; Mullany, P. Phage ϕC2 Mediates Transduction of Tn6215, Encoding Erythromycin Resistance, between Clostridium difficile Strains. mBio 2013, 4, e00840-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nale, J.Y.; Shan, J.; Hickenbotham, P.T.; Fawley, W.N.; Wilcox, M.H.; Clokie, M.R.J. Diverse Temperate Bacteriophage Carriage in Clostridium difficile 027 Strains. PLoS ONE 2012, 7, e37263. [Google Scholar] [CrossRef] [Green Version]
- Tromans, S.; Nale, J.; Clokie, M.; Jenkins, D. Understanding the contribution of the temperate bacteriophages of C. difficile to clinical outcome. J. Infect. 2010, 61, 520–521. [Google Scholar] [CrossRef]
- Nale, J.; Redgwell, T.A.; Millard, A.; Clokie, M.R.J. Efficacy of an Optimised Bacteriophage Cocktail to Clear Clostridium difficile in a Batch Fermentation Model. Antibiotics 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Nale, J.Y.; Chutia, M.; Carr, P.; Hickenbotham, P.; Clokie, M.R.J. ‘Get in early’; biofilm and wax moth (Galleria mellonella) models reveal new insights into the therapeutic potential of Clostridium difficile bacteriophages. Front. Microbiol. 2016, 7, 1383. [Google Scholar] [CrossRef] [Green Version]
- Nale, J.Y.; Chutia, M.; Cheng, J.K.J.; Clokie, M.R.J. Refining the Galleria mellonella Model by Using Stress Marker Genes to Assess Clostridioides difficile Infection and Recuperation during Phage Therapy. Microorganisms 2020, 8, 1306. [Google Scholar] [CrossRef]
- Meader, E.; Mayer, M.J.; Gasson, M.J.; Steverding, D.; Carding, S.R.; Narbad, A. Bacteriophage treatment significantly reduces viable Clostridium difficile and prevents toxin production in an in vitro model system. Anaerobe 2010, 16, 549–554. [Google Scholar] [CrossRef]
- Meader, E.; Mayer, M.J.; Steverding, D.; Carding, S.R.; Narbad, A. Evaluation of bacteriophage therapy to control Clostridium difficile and toxin production in an in vitro human colon model system. Anaerobe 2013, 22, 25–30. [Google Scholar] [CrossRef]
- Sekulovic, O.; Fortier, L.-C. Global Transcriptional Response of Clostridium difficile Carrying the ϕCD38–2 Prophage. Appl. Environ. Microbiol. 2015, 81, 1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Zhang, Y.; Dong, K.; Kuo, C.J.; Li, C.; Zhu, Y.Q.; Qin, J.; Li, Q.T.; Chang, Y.F.; Guo, X.; et al. Isolation and Characterization of the Novel Phage JD032 and Global Transcriptomic Response during JD032 Infection of Clostridioides difficile Ribotype 078. mSystems 2020, 5, e00017-20. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Kropinski, A.M.; Clokie, M.R.J. Bacteriophage behavioral ecology: How phages alter their bacterial host’s habits. Bacteriophage 2014, 4, e29866. [Google Scholar] [CrossRef] [Green Version]
- Govind, R.; Vediyappan, G.; Rolfe, R.D.; Dupuy, B.; Fralick, J.A. Bacteriophage-Mediated Toxin Gene Regulation in Clostridium difficile. J. Virol. 2009, 83, 12037–12045. [Google Scholar] [CrossRef] [Green Version]
- Dowah, A.S.A.; Xia, G.; Kareem Ali, A.A.; Thanki, A.M.; Shan, J.; Wallis, R.; Clokie, M.R.J. Identification of the receptor-binding protein of Clostridium difficile phage CDHS-1 reveals a new class of receptor-binding domains. bioRxiv 2021. [Google Scholar] [CrossRef]
- Shan, J.; Ramachandran, A.; Thanki, A.M.; Vukusic, F.B.I.; Barylski, J.; Clokie, M.R.J. Bacteriophages are more virulent to bacteria with human cells than they are in bacterial culture; insights from HT-29 cells. Sci. Rep. 2018, 8, 5091. [Google Scholar] [CrossRef]
- Nale, J.Y.; Clokie, M.R.J. Preclinical data and safety assessment of phage therapy in humans. Curr. Opin. Biotechnol. 2021, 68, 310–317. [Google Scholar] [CrossRef]
- Mukherjee, K.; Hain, T.; Fischer, R.; Chakraborty, T.; Vilcinskas, A. Brain infection and activation of neuronal repair mechanisms by the human pathogen Listeria monocytogenes in the lepidopteran model host Galleria mellonella. Virulence 2013, 4, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Rocha, D.J.P.; Santos, C.S.; Pacheco, L.G.C. Bacterial reference genes for gene expression studies by RT-qPCR: Survey and analysis. Antonie van Leeuwenhoek 2015, 108, 685–693. [Google Scholar] [CrossRef]
- Gomes, A.É.I.; Stuchi, L.P.; Siqueira, N.M.G.; Henrique, J.B.; Vicentini, R.; Ribeiro, M.L.; Darrieux, M.; Ferraz, L.F.C. Selection and validation of reference genes for gene expression studies in Klebsiella pneumoniae using Reverse Transcription Quantitative real-time PCR. Sci. Rep. 2018, 8, 9001. [Google Scholar] [CrossRef] [PubMed]
- Govind, R.; Fralick, J.A.; Rolfe, R.D. In vivo lysogenization of a Clostridium difficile bacteriophage ФCD119. Anaerobe 2011, 17, 125–129. [Google Scholar]
- Sørensen, M.C.H.; Alphen, L.B.; Harboe, A.; Li, J.; Christensen, B.B.; Szymanski, C.M.; Brøndsted, L. Bacteriophage F336 Recognizes the Capsular Phosphoramidate Modification of Campylobacter jejuni NCTC11168. J. Bacteriol. 2011, 193, 6742–6749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekulovic, O.; Meessen-Pinard, M.; Fortier, L.C. Prophage-stimulated toxin production in Clostridium difficile NAP1/027 lysogens. J. Bacteriol. 2011, 193, 2726–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, S.; Chang, B.J.; Riley, T.V. Effect of phage infection on toxin production by Clostridium difficile. J. Med. Microbiol. 2005, 54, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C.; Moineau, S. Morphological and genetic diversity of temperate phages in Clostridium difficile. Appl. Environ. Microbiol. 2007, 73, 7358–7366. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, V.; Fralick, J.A.; Rolfe, R.D. Prevention of Clostridium difficile -induced ileocecitis with Bacteriophage. Anaerobe 1999, 5, 69–78. [Google Scholar] [CrossRef]
- Duffy, C.; Feiss, M. The large subunit of bacteriophage λ’s terminase plays a role in DNA translocation and packaging termination1 1Edited by M. Gottesman. J. Mol. Biol. 2002, 316, 547–561. [Google Scholar] [CrossRef]
- Shen, X.; Li, M.; Zeng, Y.; Hu, X.; Tan, Y.; Rao, X.; Jin, X.; Li, S.; Zhu, J.; Zhang, K.; et al. Functional identification of the DNA packaging terminase from Pseudomonas aeruginosa phage PaP3. Arch. Virol. 2012, 157, 2133–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauss, P.; Park, K.; Spencer, T.E.; Hacker, K.J. DNA helicase requirements for DNA replication during bacteriophage T4 infection. J. Bacteriol. 1994, 176, 1667–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soengas, M.a.S.; Gutiérrez, C.; Salas, M. Helix-destabilizing Activity of φ29 Single-stranded DNA Binding Protein: Effect on the Elongation Rate During Strand Displacement DNA Replication. J. Mol. Biol. 1995, 253, 517–529. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, H.; Gu, Z.; Roberts, J.W. High-resolution view of bacteriophage lambda gene expression by ribosome profiling. Proc. Natl. Acad. Sci. USA 2013, 110, 11928–11933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundling, A.; Manson, M.D.; Young, R. Holins kill without warning. Proc. Natl. Acad. Sci. USA 2001, 98, 9348–9352. [Google Scholar] [CrossRef] [Green Version]
- Young, R. Phage lysis: Do we have the hole story yet? Curr. Opin. Microbiol. 2013, 16, 790–797. [Google Scholar] [CrossRef] [Green Version]
- Delisle, A.L.; Barcak, G.J.; Guo, M. Isolation and Expression of the Lysis Genes of Actinomyces naeslundii Phage Av-1. Appl. Environ. Microbiol. 2006, 72, 1110–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardarelli, L.; Lam, R.; Tuite, A.; Baker, L.A.; Sadowski, P.D.; Radford, D.R.; Rubinstein, J.L.; Battaile, K.P.; Chirgadze, N.; Maxwell, K.L.; et al. The Crystal Structure of Bacteriophage HK97 gp6: Defining a Large Family of Head–Tail Connector Proteins. J. Mol. Biol. 2010, 395, 754–768. [Google Scholar] [CrossRef]
- Ionel, A.; Velázquez-Muriel, J.A.; Luque, D.; Cuervo, A.; Castón, J.R.; Valpuesta, J.M.; Martín-Benito, J.; Carrascosa, J.L. Molecular rearrangements involved in the capsid shell maturation of bacteriophage T7. J. Biol. Chem. 2011, 286, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Leiman, P.G.; Arisaka, F.; van Raaij, M.J.; Kostyuchenko, V.A.; Aksyuk, A.A.; Kanamaru, S.; Rossmann, M.G. Morphogenesis of the T4 tail and tail fibers. Virol. J. 2010, 7, 355. [Google Scholar] [CrossRef] [Green Version]
- Nale, J.Y.; Vinner, G.K.; Lopez, V.C.; Thanki, A.M.; Phothaworn, P.; Thiennimitr, P.; Garcia, A.; AbuOun, M.; Anjum, M.F.; Korbsrisate, S.; et al. An Optimized Bacteriophage Cocktail Can Effectively Control Salmonella in vitro and in Galleria mellonella. Front. Microbiol. 2021, 11, 609955. [Google Scholar] [CrossRef] [PubMed]
- Wagley, S.; Borne, R.; Harrison, J.; Baker-Austin, C.; Ottaviani, D.; Leoni, F.; Vuddhakul, V.; Titball, R.W. Galleria mellonella as an infection model to investigate virulence of Vibrio parahaemolyticus. Virulence 2018, 9, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, K.; Altincicek, B.; Hain, T.; Domann, E.; Vilcinskas, A.; Chakraborty, T. Galleria mellonella as a Model System for Studying Listeria Pathogenesis. Appl. Environ. Microbiol. 2010, 76, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Conditions | Time Post Phage Infection (Min) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Infected R20291 vs. Control | Infected R20291 vs. 0 Min Baseline | |||||||||||
| 0 | 10 | 20 | 30 | 40 | 50 | 10 | 20 | 30 | 40 | 50 | ||
| Bacteria, R20291 genes | Significantly expressed | 442 | 359 | 433 | 442 | 456 | 450 | 309 | 324 | 478 | 556 | 561 |
| Upregulated genes | 14 | 20 | 20 | 20 | 42 | 46 | 35 | 67 | 131 | 162 | 170 | |
| Downregulated genes | 428 | 359 | 433 | 442 | 456 | 450 | 274 | 257 | 347 | 394 | 391 | |
| Phage, CDHS-1 genes | Significantly expressed | 13 | 25 | 28 | 30 | 26 | 24 | 27 | 35 | 34 | 33 | 37 |
| Upregulated genes | 13 | 25 | 28 | 30 | 26 | 24 | 23 | 28 | 29 | 28 | 27 | |
| Downregulated genes | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 7 | 5 | 5 | 10 | |
| Host | Time | Upregulated | Downregulated |
|---|---|---|---|
| R20291 | 0 | Nucleotide and ATP binding proteins | Metabolic process and membrane protein |
| 10 | DNA and ATP binding, and permease protein | Metabolic process, integral component of membrane | |
| 20 | DNA binding, transcription and folding proteins | Putative membrane protein | |
| 30 | DNA binding protein, transcription and folding proteins | Putative membrane protein and conserved hypothetical protein | |
| 40 | DNA binding protein, transcription | Permease protein, putative membrane protein | |
| 50 | Membrane and integral component of membrane | Permease protein, putative membrane protein | |
| CDHS-1 | 0 | Cro/C1-type transcriptional regulator, ssDNA-binding and helicase protein | None |
| 10 | Cro/C1-type transcriptional regulator, ssDNA-binding protein | None | |
| 20 | Holin, endolysin and tail fiber protein | None | |
| 30 | Holin, endolysin and tail fiber protein | None | |
| 40 | Endolysin and tail fiber protein | None | |
| 50 | Tail fiber protein, endolysin and minor capsid protein | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nale, J.Y.; Al-Tayawi, T.S.; Heaphy, S.; Clokie, M.R.J. Impact of Phage CDHS-1 on the Transcription, Physiology and Pathogenicity of a Clostridioides difficile Ribotype 027 Strain, R20291. Viruses 2021, 13, 2262. https://doi.org/10.3390/v13112262
Nale JY, Al-Tayawi TS, Heaphy S, Clokie MRJ. Impact of Phage CDHS-1 on the Transcription, Physiology and Pathogenicity of a Clostridioides difficile Ribotype 027 Strain, R20291. Viruses. 2021; 13(11):2262. https://doi.org/10.3390/v13112262
Chicago/Turabian StyleNale, Janet Y., Thekra Sideeq Al-Tayawi, Shaun Heaphy, and Martha R. J. Clokie. 2021. "Impact of Phage CDHS-1 on the Transcription, Physiology and Pathogenicity of a Clostridioides difficile Ribotype 027 Strain, R20291" Viruses 13, no. 11: 2262. https://doi.org/10.3390/v13112262
APA StyleNale, J. Y., Al-Tayawi, T. S., Heaphy, S., & Clokie, M. R. J. (2021). Impact of Phage CDHS-1 on the Transcription, Physiology and Pathogenicity of a Clostridioides difficile Ribotype 027 Strain, R20291. Viruses, 13(11), 2262. https://doi.org/10.3390/v13112262