
NMR Experiments Shed New Light on Glycan Recognition by Human and Murine Norovirus Capsid Proteins

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Biosynthesis and Purification
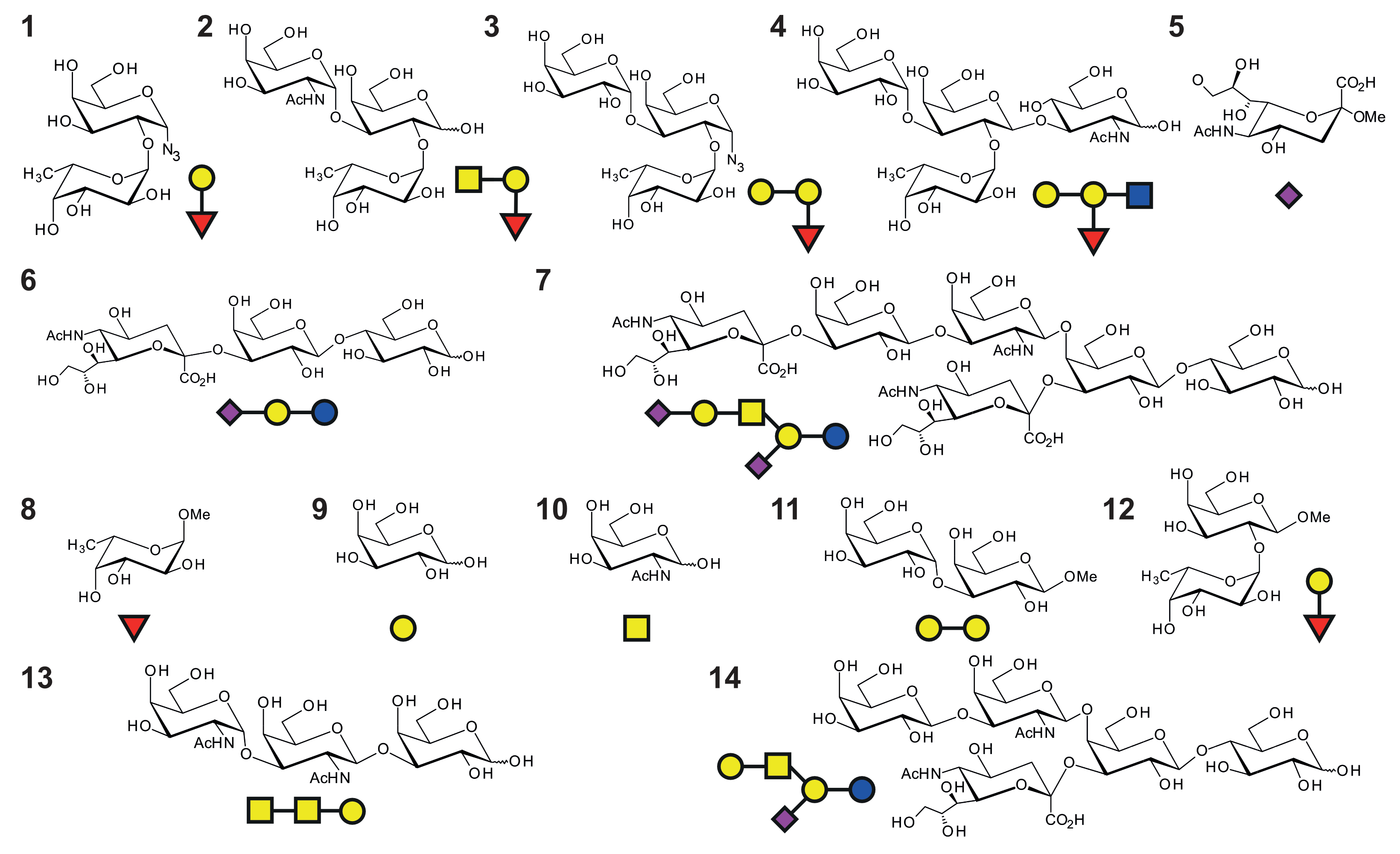
2.2. Enzymatic Synthesis of Blood Group B-Trisaccharide (3)
2.3. NMR Spectroscopy
2.4. Cell Culture
2.5. MNV Cultivation and Titration
2.6. Production of Lentiviral Vectors and Transduction of CHO Derived Cell Lines
3. Results
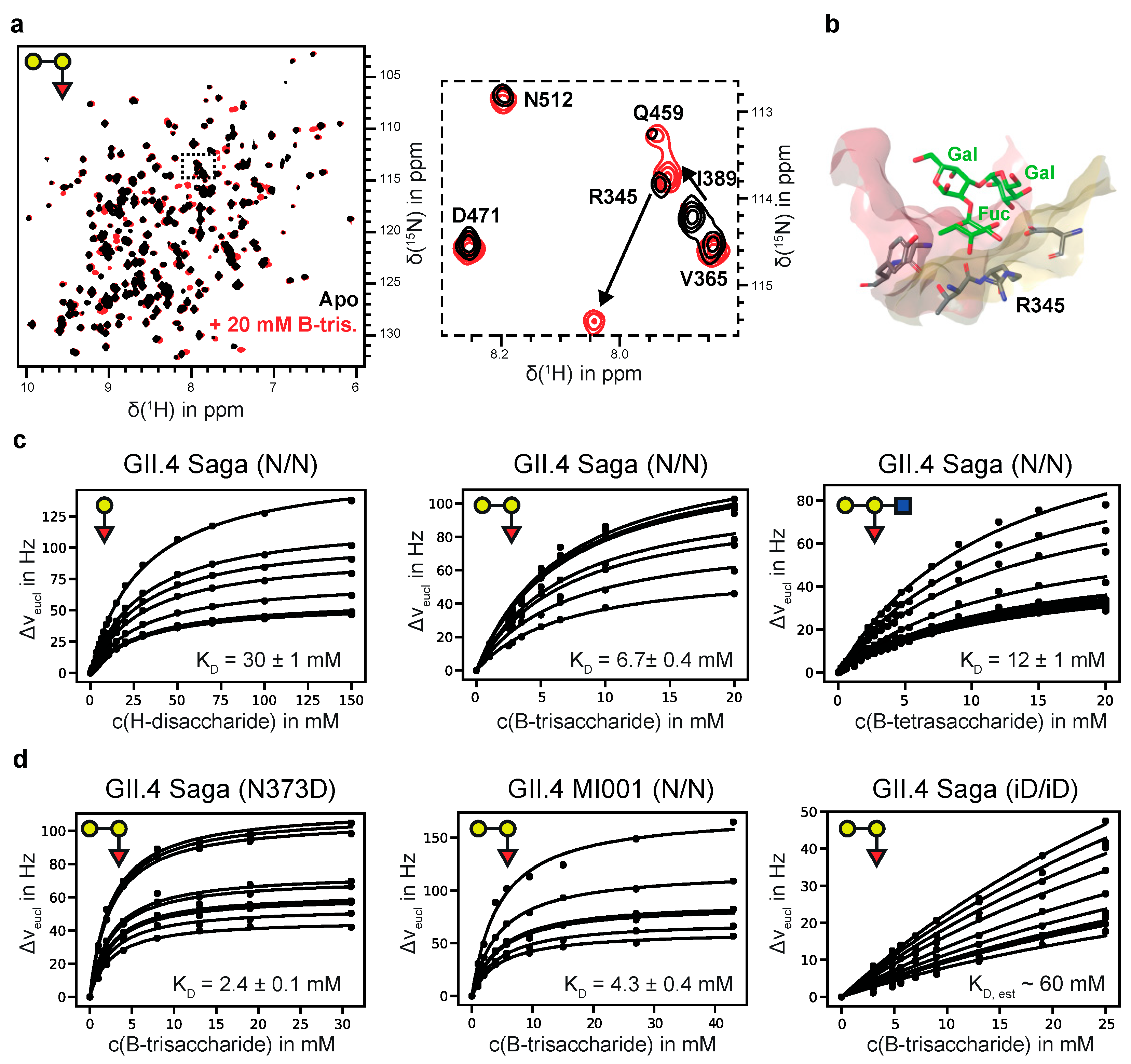
3.1. Chemical Shift Perturbation (CSP) NMR Experiments Provide Accurate Dissociation Constants for HBGAs Binding to Human GII.4 SAGA and MI001 P-Dimers
3.2. MNV P-Domains Do Not Bind to HBGAs or Other Neutral Glycans
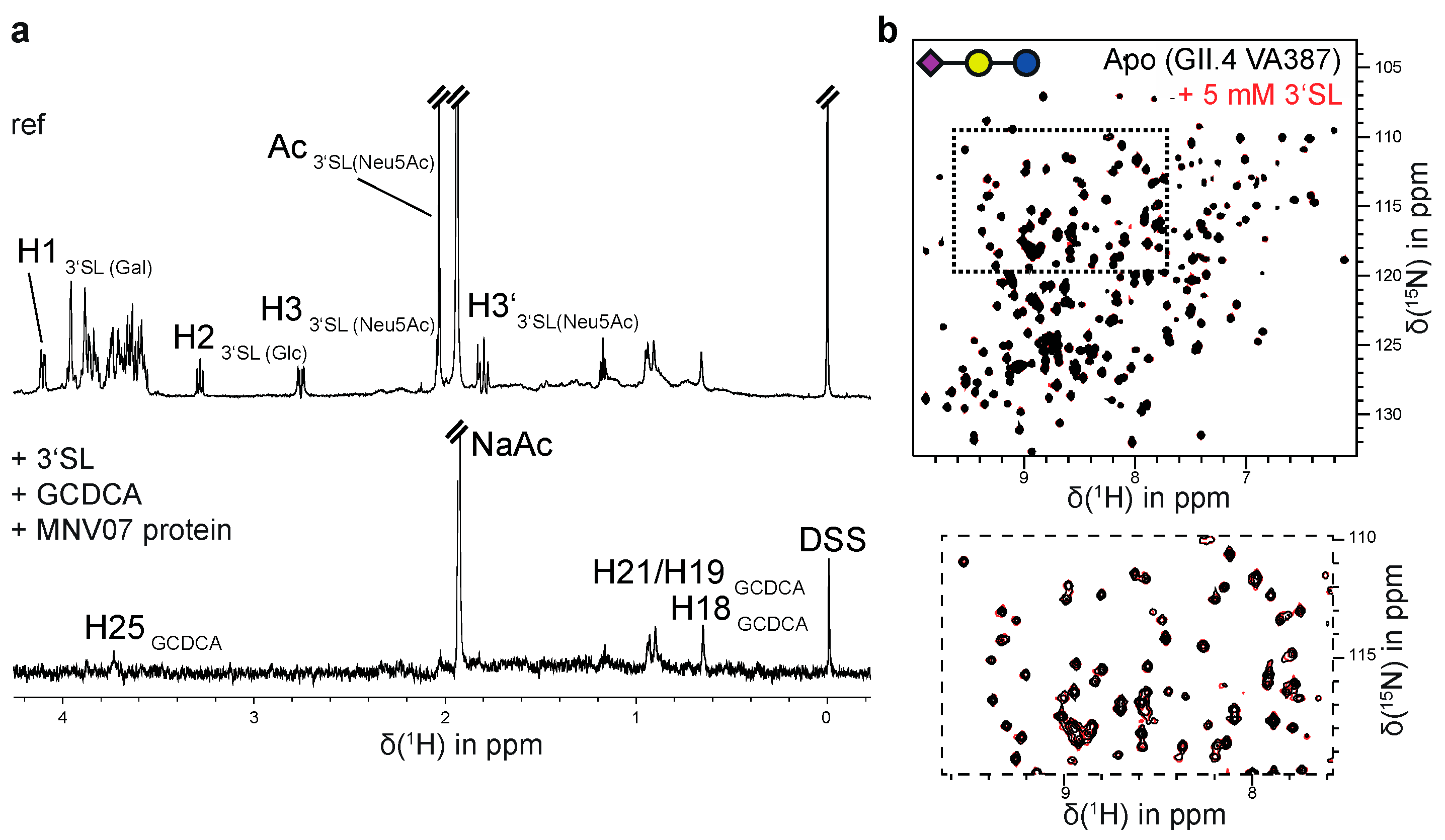
3.3. MNV and GII.4 P-Dimers Do Not Recognize Sialic Acid
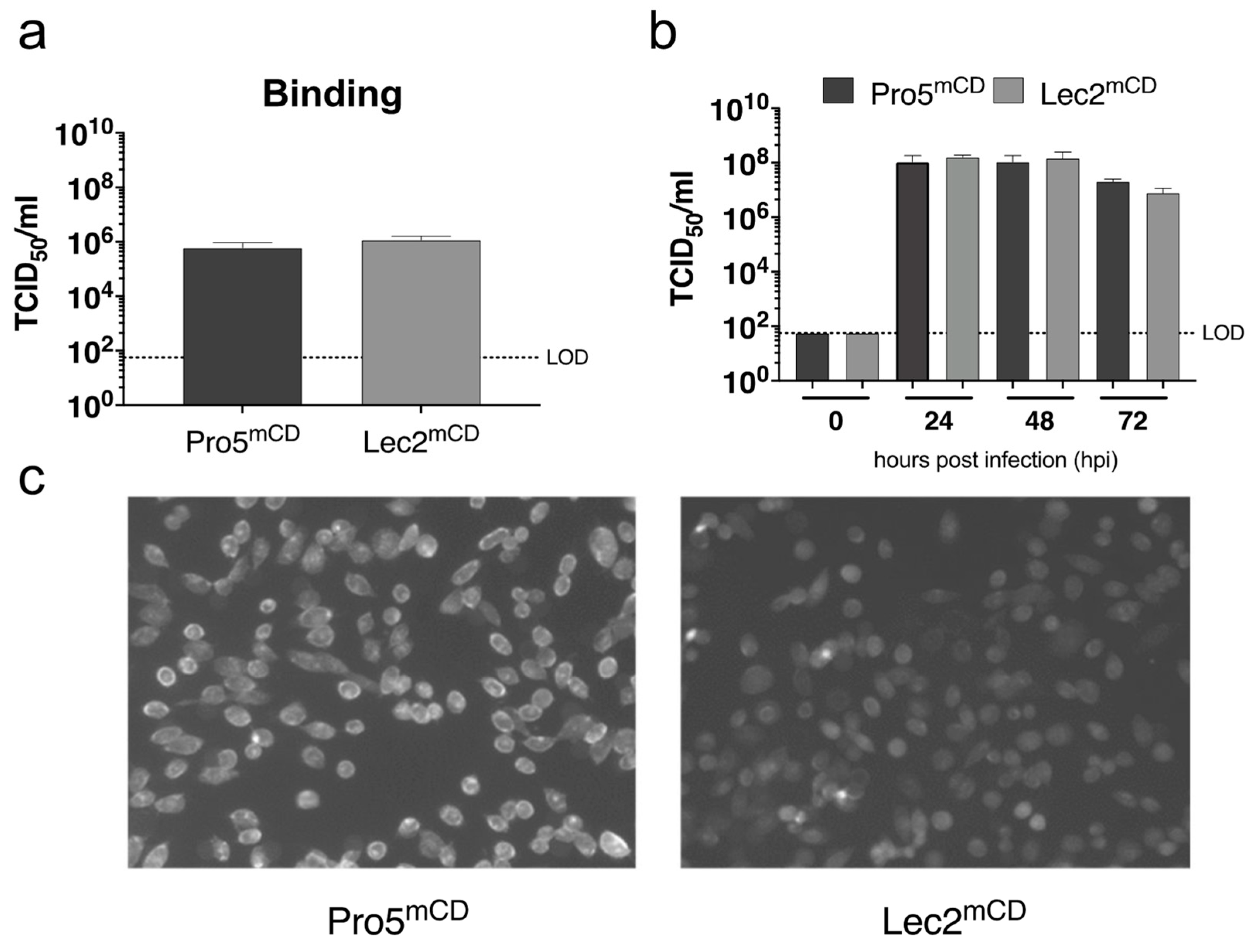
3.4. Sialic Acid on CHO Derived Cells Expressing the MNV-1 Entry Receptor Does Not Contribute to Infection
3.5. Glycan Binding to HuNoV P-Domains Has No Equivalent in MNV P-Dimers
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chhabra, P.; De Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- van Beek, J.; de Graaf, M.; Al-Hello, H.; Allen, D.J.; Ambert-Balay, K.; Botteldoorn, N.; Koopmans, M.P. Molecular surveillance of norovirus, 2005–16: An epidemiological analysis of data collected from the NoroNet network. Lancet Infect. Dis. 2018, 18, 545–553. [Google Scholar] [CrossRef]
- de Graaf, M.; van Beek, J.; Koopmans, M.P. Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 2016, 14, 421–433. [Google Scholar] [CrossRef]
- Cao, S.; Lou, Z.; Tan, M.; Chen, Y.; Liu, Y.; Zhang, Z.; Zhang, X.C.; Jiang, X.; Li, X.; Rao, Z. Structural basis for the recognition of blood group trisaccharides by norovirus. J. Virol. 2007, 81, 5949–5957. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Farkas, T.; Marionneau, S.; Zhong, W.; Ruvoën-Clouet, N.; Morrow, A.L.; Altaye, M.; Pickering, L.K.; Newburg, D.S.; LePendu, J.; et al. Noroviruses bind to human ABO, Lewis, and secretor histo-blood group antigens: Identification of 4 distinct strain-specific patterns. J. Infect. Dis. 2003, 188, 19–31. [Google Scholar] [CrossRef]
- Marionneau, S.; Ruvoën, N.; Le Moullac–Vaidye, B.; Clement, M.; Cailleau–Thomas, A.; Ruiz–Palacois, G.; Huang, P.; Jiang, X.; Le Pendu, J. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 2002, 122, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Bartnicki, E.; Cunha, J.B.; Kolawole, A.O.; Wobus, C.E. Recent advances in understanding noroviruses. F1000Research 2017, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M.; Wobus, C.E. A working model of how noroviruses infect the intestine. PLOS Pathog. 2015, 11, e1004626. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.K.; Leuthold, M.M.; Hansman, G.S. Structural Constraints on Human Norovirus Binding to Histo-Blood Group Antigens. mSphere 2016, 1, e00049-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona-Vicente, N.; Vila-Vicent, S.; Allen, D.; Gozalbo-Rovira, R.; Iturriza-Gómara, M.; Buesa, J.; Rodríguez-Díaz, J. Characterization of a Novel Conformational GII.4 Norovirus Epitope: Implications for Norovirus-Host Interactions. J. Virol. 2016, 90, 7703–7714. [Google Scholar] [CrossRef] [Green Version]
- Guix, S.; Asanaka, M.; Katayama, K.; Crawford, S.E.; Neill, F.H.; Atmar, R.L.; Estes, M.K. Norwalk virus RNA is infectious in mammalian cells. J. Virol. 2007, 81, 12238–12248. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Ettayebi, K.; Tenge, V.R.; Karandikar, U.C.; Lewis, M.A.; Lin, S.-C.; Neill, F.H.; Ayyar, B.V.; Zeng, X.-L.; Larson, G.; et al. Genetic Manipulation of Human Intestinal Enteroids Demonstrates the Necessity of a Functional Fucosyltransferase 2 Gene for Secretor-Dependent Human Norovirus Infection. mBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.A.; Wilen, C.B.; Dai, Y.-N.; Orchard, R.C.; Kim, A.S.; Stegeman, R.A.; Hsieh, L.L.; Smith, T.J.; Virgin, H.W.; Fremont, D.H. Structural basis for murine norovirus engagement of bile acids and the CD300lf receptor. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, T.; Koromyslova, A.; Malak, V.; Hansman, G.S. Atomic structure of the murine norovirus protruding domain and sCD300lf receptor complex. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Fujimoto, A.; Takai-Todaka, R.; Miki, M.; Doan, Y.H.; Murakami, K.; Yokoyama, M.; Murata, K.; Nakanishi, A.; Katayama, K. Functional receptor molecules CD300lf and CD300ld within the CD300 family enable murine noroviruses to infect cells. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef] [Green Version]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldridge, M.T.; McCune, B.T.; Lee, Y.-C.J.; Lee, S.; Pruett-Miller, S.M.; Nelson, C.A.; Fremont, D.H.; et al. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Stuart, A.D.; Brown, T.D.K. Alpha2,6-linked sialic acid acts as a receptor for Feline calicivirus. J. Gen. Virol. 2007, 88, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Wei, C.; Huang, P.; Fan, Q.; Quigley, C.; Xia, M.; Fang, H.; Zhang, X.; Zhong, W.; Klassen, J.S.; et al. Tulane virus recognizes sialic acids as cellular receptors. Sci. Rep. 2015, 5, 11784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, S.; Perry, J.W.; Yetming, K.; Patel, S.P.; Auble, H.; Shu, L.; Nawar, H.F.; Lee, C.H.; Connell, T.D.; Shayman, J.A.; et al. Ganglioside-linked terminal sialic acid moieties on murine macrophages function as attachment receptors for murine noroviruses. J. Virol. 2009, 83, 4092–4101. [Google Scholar] [CrossRef] [Green Version]
- Taube, S.; Perry, J.W.; McGreevy, E.; Yetming, K.; Perkins, C.; Henderson, K.; Wobus, C.E. Murine noroviruses bind glycolipid and glycoprotein attachment receptors in a strain-dependent manner. J. Virol. 2012, 86, 5584–5593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegener, H.; Mallagaray, Á.; Schöne, T.; Peters, T.; Lockhauserbäumer, J.; Yan, H.; Uetrecht, C.; Hansman, G.S.; Taube, S. Human norovirus GII.4(MI001) P dimer binds fucosylated and sialylated carbohydrates. Glycobiology 2017, 27, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Tan, M.; Xia, M.; Kitova, E.N.; Jiang, X.; Klassen, J.S. Gangliosides are ligands for human noroviruses. J. Am. Chem. Soc. 2014, 136, 12631–12637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, S.; Mallagaray, A.; Peters, T. Norovirus, glycans and attachment. Curr. Opin. Virol. 2018, 31, 33–42. [Google Scholar] [CrossRef]
- Shanker, S.; Hu, L.; Ramani, S.; Atmar, R.L.; Estes, M.K.; Prasad, B.V.V. Structural features of glycan recognition among viral pathogens. Curr. Opin. Struct. Biol. 2017, 44, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Fiege, B.; Rademacher, C.; Cartmell, J.; Kitov, P.I.; Parra, F.; Peters, T. Molecular Details of the Recognition of Blood Group Antigens by a Human Norovirus as Determined by STD NMR Spectroscopy. Angew. Chem. Int. Ed. 2012, 51, 928–932. [Google Scholar] [CrossRef] [PubMed]
- Mallagaray, A.; Rademacher, C.; Parra, F.; Hansman, G.; Peters, T. STD NMR Titrations Reveal Complex Multistep-Binding of L-Fucose to Norovirus Particles. Glycobiology 2017, 27, 80–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallagaray, A.; Lockhauserbäumer, J.; Hansman, G.; Uetrecht, C.; Peters, T. Attachment of Norovirus to Histo Blood Group Antigens: A Cooperative Multistep Process. Angew. Chem. Int. Ed. 2015, 54, 12014–12019. [Google Scholar] [CrossRef] [PubMed]
- Hansman, G.S.; Shahzad-Ul-Hussan, S.; McLellan, J.S.; Chuang, G.-Y.; Georgiev, I.; Shimoike, T.; Katayama, K.; Bewley, C.A.; Kwong, P.D. Structural basis for norovirus inhibition and fucose mimicry by citrate. J. Virol. 2012, 86, 284–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Zheng, R.; Richards, M.R.; Tan, M.; Kitova, E.N.; Jiang, X.; Klassen, J.S. Quantifying the Binding Stoichiometry and Affinity of Histo-Blood Group Antigen Oligosaccharides for Human Noroviruses. Glycobiology 2018, 28, 488–498. [Google Scholar] [CrossRef]
- Han, L.; Kitova, E.N.; Tan, M.; Jiang, X.; Pluvinage, B.; Boraston, A.B.; Klassen, J.S. Affinities of human histo-blood group antigens for norovirus capsid protein complexes. Glycobiology 2015, 25, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Kitov, P.I.; Kitova, E.N.; Tan, M.; Wang, L.; Xia, M.; Jiang, X.; Klassen, J.S. Affinities of recombinant norovirus P dimers for human blood group antigens. Glycobiology 2013, 23, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Nasir, W.; Bally, M.; Zhdanov, V.P.; Larson, G.; Höök, F. Interaction of Virus-Like Particles with Vesicles Containing Glycolipids: Kinetics of Detachment. J. Phys. Chem. B 2015, 119, 11466–11472. [Google Scholar] [CrossRef]
- Parveen, N.; Rydell, G.E.; Larson, G.; Hytönen, V.P.; Zhdanov, V.P.; Höök, F.; Block, S. Competition for Membrane Receptors: Norovirus Detachment via Lectin Attachment. J. Am. Chem. Soc. 2019, 141, 16303–16311. [Google Scholar] [CrossRef] [PubMed]
- Nasir, W.; Frank, M.; Kunze, A.; Bally, M.; Parra, F.; Nyholm, P.-G.; Höök, F.; Larson, G. Histo-Blood Group Antigen Presentation Is Critical for Binding of Norovirus VLP to Glycosphingolipids in Model Membranes. ACS Chem. Biol. 2017, 12, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Mallagaray, A.; Creutznacher, R.; Dülfer, J.; Mayer, P.H.O.; Grimm, L.L.; Orduña, J.M.; Trabjerg, E.; Stehle, T.; Rand, K.D.; Blaum, B.S.; et al. A post-translational modification of human Norovirus capsid protein attenuates glycan binding. Nat. Commun. 2019, 10, 1320. [Google Scholar] [CrossRef] [Green Version]
- Creutznacher, R.; Schulze, E.; Wallmann, G.; Peters, T.; Stein, M.; Mallagaray, A. Chemical-Shift Perturbations Reflect Bile Acid Binding to Norovirus Coat Protein: Recognition Comes in Different Flavors. ChemBioChem 2020, 21, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Hansman, G.S.; Doan, L.T.P.; Kguyen, T.A.; Okitsu, S.; Katayama, K.; Ogawa, S.; Natori, K.; Takeda, N.; Kato, Y.; Nishio, O.; et al. Detection of norovirus and sapovirus infection among children with gastroenteritis in Ho Chi Minh City, Vietnam. Arch. Virol. 2004, 149, 1673–1688. [Google Scholar] [CrossRef] [PubMed]
- Seto, N.O.; Compston, C.A.; Szpacenko, A.; Palcic, M.M. Enzymatic synthesis of blood group A and B trisaccharide analogues. Carbohydr. Res. 2000, 324, 161–169. [Google Scholar] [CrossRef]
- Angulo, J.; Langpap, B.; Blume, A.; Biet, T.; Meyer, B.; Krishna, N.R.; Peters, H.; Palcic, M.M.; Peters, T. Blood Group B Galactosyltransferase: Insights into Substrate Binding from NMR Experiments. J. Am. Chem. Soc. 2006, 128, 13529–13538. [Google Scholar] [CrossRef]
- Sindhuwinata, N.; Munoz, E.; Munoz, F.J.; Palcic, M.M.; Peters, H.; Peters, T. Binding of an acceptor substrate analog enhances the enzymatic activity of human blood group B galactosyltransferase. Glycobiology 2010, 20, 718–723. [Google Scholar] [CrossRef] [Green Version]
- Baryshnikova, O.K.; Williams, T.C.; Sykes, B.D. Internal pH indicators for biomolecular NMR. J. Biomol. NMR 2008, 41, 5–7. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins Struct. Funct. Bioinform. 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Williamson, M.P. Using chemical shift perturbation to characterise ligand binding. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Meyer, B. Characterization of Ligand Binding by Saturation Transfer Difference NMR Spectroscopy. Angew. Chem. Int. Ed. 1999, 38, 1784–1787. [Google Scholar] [CrossRef]
- Hwang, S.; Alhatlani, B.; Arias, A.; Caddy, S.L.; Christodoulou, C.; Cunha, J.B.; Emmott, E.; Gonzalez-Hernandez, M.; Kolawole, A.; Lu, J.; et al. Murine Norovirus: Propagation, quantification, and genetic manipulation. Curr. Protoc. Microbiol. 2014, 33, 15K.2.1–15K.2.61. [Google Scholar] [CrossRef] [PubMed]
- Zufferey, R.; Nagy, D.; Mandel, R.J.; Naldini, L.; Trono, D. Multiply attenuated lentiviral vector achieves efficient gene delivery in vivo. Nat. Biotechnol. 1997, 15, 871–875. [Google Scholar] [CrossRef]
- Naldini, L.; Blomer, U.; Gage, F.H.; Trono, D.; Verma, I.M. Efficient transfer, integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector. Proc. Natl. Acad. Sci. USA 1996, 93, 11382–11388. [Google Scholar] [CrossRef] [Green Version]
- Kaludov, N.; Brown, K.E.; Walters, R.W.; Zabner, J.; Chiorini, J.A. Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity. J. Virol. 2001, 75, 6884–6893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galili, U.; Mandrell, R.E.; Hamadeh, R.M.; Shohet, S.B.; Griffiss, J.M. Interaction between human natural anti-alpha-galactosyl immunoglobulin G and bacteria of the human flora. Infect. Immun. 1988, 56, 1730–1737. [Google Scholar] [CrossRef] [Green Version]
- Zakhour, M.; Ruvoen-Clouet, N.; Charpilienne, A.; Langpap, B.; Poncet, D.; Peters, T.; Bovin, N.; Le Pendu, J. The aGal Epitope of the Histo-Blood Group Antigen Family Is a Ligand for Bovine Norovirus Newbury2 Expected to Prevent Cross-Species Transmission. PLoS Pathog. 2009, 5, e1000504. [Google Scholar] [CrossRef] [Green Version]
- Hakomori, S.I. Tumor-Associated Carbohydrate Antigens. Annu. Rev. Immunol. 1984, 2, 103–126. [Google Scholar] [CrossRef] [PubMed]
- Müller-Hermes, C.; Creutznacher, R.; Mallagaray, A. Complete assignment of Ala, Ile, Leu(ProS), Met and Val(ProS) methyl groups of the protruding domain from human norovirus GII.4 Saga. Biomol. NMR Assign. 2020, 14, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, T.; Koromyslova, A.; Hansman, G.S. Structural Basis for Human Norovirus Capsid Binding to Bile Acids. J. Virol. 2019, 93, e01581-18. [Google Scholar] [CrossRef] [Green Version]
- Stott, K.; Stonehouse, J.; Keeler, J.; Hwang, T.-L.; Shaka, A.J. Excitation Sculpting in High-Resolution Nuclear Magnetic Resonance Spectroscopy: Application to Selective NOE Experiments. J. Am. Chem. Soc. 1995, 117, 4199–4200. [Google Scholar] [CrossRef]
- Rydell, G.; Svensson, L.; Larson, G.; Johannes, L.; Römer, W. Human GII.4 norovirus VLP induces membrane invaginations on giant unilamellar vesicles containing secretor gene dependent alpha1,2-fucosylated glycosphingolipids. Biochim. Biophys. Acta 2013, 1828, 1840–1845. [Google Scholar] [CrossRef] [Green Version]
- Rydell, G.E.; Dahlin, A.B.; Höök, F.; Larson, G. QCM-D studies of human norovirus VLPs binding to glycosphingolipids in supported lipid bilayers reveal strain-specific characteristics. Glycobiology 2009, 19, 1176–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, H.; Roemer, W.; Smith, A.E.; Bacia, K.; Dmitrieff, S.; Chai, W.; Mancini, R.; Kartenbeck, J.; Chambon, V.; Berland, L.; et al. GM1 structure determines SV40-induced membrane invagination and infection. Nat. Cell Biol. 2010, 12, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Neu, U.; Woellner, K.; Gauglitz, G.; Stehle, T. Structural basis of GM1 ganglioside recognition by simian virus. Proc. Natl. Acad. Sci. USA 2008, 105, 5219–5224. [Google Scholar] [CrossRef] [Green Version]
- Tsai, B.; Gilbert, J.M.; Stehle, T.; Lencer, W.; Benjamin, T.L.; Rapoport, T.A. Gangliosides are receptors for murine polyoma virus and SV40. EMBO J. 2003, 22, 4346–4355. [Google Scholar] [CrossRef] [Green Version]
- Alvaro, M.; University of Lubeck, Lubeck, Germany. Personal communication, 2021.
- Charlotte, U.; Heinrich Pette Institute, Hamburg, Germany. Personal communication, 2020.
- Chen, Y.; Tan, M.; Xia, M.; Hao, N.; Zhang, X.C.; Huang, P.; Jiang, X.; Li, X.; Rao, Z. Crystallography of a Lewis-binding norovirus, elucidation of strain-specificity to the polymorphic human histo-blood group antigens. PLOS Pathog. 2011, 7, e1002152. [Google Scholar] [CrossRef]
- Rydell, G.E.; Nilsson, J.; Rodriguez-Diaz, J.; Ruvoën-Clouet, N.; Svensson, L.; Le Pendu, J.; Larson, G. Human noroviruses recognize sialyl Lewis x neoglycoprotein. Glycobiology 2009, 19, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, S.; Jiang, M.; Wobus, C.E. Glycosphingolipids as Receptors for non-enveloped viruses. Viruses 2010, 2, 1011–1049. [Google Scholar] [CrossRef] [PubMed]
- Taube, S.; Rubin, J.R.; Katpally, U.; Smith, T.J.; Kendall, A.; Stuckey, J.A.; Wobus, C.E. High-resolution X-ray structure and functional analysis of the murine norovirus 1 capsid protein protruding domain. J. Virol. 2010, 84, 5695–5705. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NoV Strain | Ligand | KD/mM |
|---|---|---|
| GII.4 Saga (N/N) | H-Dis 1 | 30.0 ± 1 |
| GII.4 Saga (N/N) | A-Tri 2 | 4.3 ± 0.4 |
| GII.4 Saga (N/N) | B-Tri 3 | 6.7 ± 0.4 |
| GII.4 Saga (N/N) | B-Tetra 4 | 12.0 ± 1 |
| GII.4 Saga (iD/iD) | B-Tri 3 | ca. 60 |
| GII.4 Saga (N373D) | B-Tri 3 | 2.4 ± 0.1 |
| GII.4 MI001 (N/N) | B-Tri 3 | 4.3 ± 0.4 |
| GII.10 Vietnam 026 VLPs | B-Tri 3 | 7.9 ± 0.7 |
| GII.10 Vietnam 026 VLPs (with saturating amounts of GCDCA [a]) | B-Tri 3 | 9.0 ± 1 |
| MNV Strain | Glycan Ligand | Exp [a] | B [b] | GCDCA | EDTA | Ca2+ |
|---|---|---|---|---|---|---|
| CW1 | Fuc 8 | STD | B | - | - | - |
| CW1 | H-Dis 12 | CSP | B | + | - | - |
| CW1 | A-Tri 2 | STD | B* | + | - | - |
| MNV07 | B-Tri 3 | CSP | B | - | - | - |
| CW1 | B-Tri 3 | CSP | B | - | - | - |
| MNV07 | B-Tri 3 | STD | B | + | - | - |
| CW1 | B-Tri 3 | CSP | B | + | - | - |
| CW1 | B-Tri 3 | CSP | B | + | + | - |
| CW1 | GalNAc 10 | STD | B* | + | - | - |
| CW1 | Fuc 8, Gal 9 | CSP | B | + | - | - |
| CW1 | Galili Dis 11 | CSP | B | + | - | - |
| CW1 | Forssman antigen 13 | STD | B* | + | - | - |
| MNV07 | Neu5Ac 5 | STD | A* | - | - | - |
| CW1 | Neu5Ac 5 | CSP | B | + | - | - |
| CW1 | Neu5Ac 5 | CSP | B | + | + | - |
| CW1 | Neu5Ac 5 | CSP | A | + | - | + |
| CW1 | 3′SL 6 | CSP | B | - | - | - |
| MNV07 | 3′SL 6 | STD | B | + | - | - |
| CW1 | 3′SL 6 | CSP | B | + | - | - |
| CW1 | 3′SL 6 | CSP | B | + | - | + |
| CW1 | 3′SL 6 | CSP | B | + | + | + |
| CW1 | GM1a 14 | CSP | B | + | - | - |
| MNV07 | GD1a 7 | CSP | A* | - | - |
| HuNoV Strain | Glycan Ligand | Buffer |
|---|---|---|
| GII.4 Saga (N/N) | Neu5Ac 5 | A |
| 3′SL 6 | B | |
| 3′SL 6 | C | |
| 3′SL 6 | D | |
| GII.4 Saga (iD/iD) | 3′SL 6 | B |
| GII.4 MI001 (N/N) | Neu5Ac 5 | B |
| 3′SL 6 | E | |
| GII.4 VA387 (N/N) | 3′SL 6 | B |
| 3′SL 6 | F | |
| 3′SL 6 | G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Creutznacher, R.; Maass, T.; Ogrissek, P.; Wallmann, G.; Feldmann, C.; Peters, H.; Lingemann, M.; Taube, S.; Peters, T.; Mallagaray, A. NMR Experiments Shed New Light on Glycan Recognition by Human and Murine Norovirus Capsid Proteins. Viruses 2021, 13, 416. https://doi.org/10.3390/v13030416
Creutznacher R, Maass T, Ogrissek P, Wallmann G, Feldmann C, Peters H, Lingemann M, Taube S, Peters T, Mallagaray A. NMR Experiments Shed New Light on Glycan Recognition by Human and Murine Norovirus Capsid Proteins. Viruses. 2021; 13(3):416. https://doi.org/10.3390/v13030416
Chicago/Turabian StyleCreutznacher, Robert, Thorben Maass, Patrick Ogrissek, Georg Wallmann, Clara Feldmann, Hannelore Peters, Marit Lingemann, Stefan Taube, Thomas Peters, and Alvaro Mallagaray. 2021. "NMR Experiments Shed New Light on Glycan Recognition by Human and Murine Norovirus Capsid Proteins" Viruses 13, no. 3: 416. https://doi.org/10.3390/v13030416
APA StyleCreutznacher, R., Maass, T., Ogrissek, P., Wallmann, G., Feldmann, C., Peters, H., Lingemann, M., Taube, S., Peters, T., & Mallagaray, A. (2021). NMR Experiments Shed New Light on Glycan Recognition by Human and Murine Norovirus Capsid Proteins. Viruses, 13(3), 416. https://doi.org/10.3390/v13030416