1. Introduction
Foot-and-mouth disease (FMD) is a highly contagious vesicular disease caused by FMD virus (FMDV), an aphthovirus within the family
picornaviridae that infects both domesticated and wild cloven-hoofed animals [
1]. The classical FMD symptoms in ruminants are characterised by fever, inappetence, lameness, excess salivation and vesicles in and around the mouth, teats and feet. These clinical signs normally subside approximately 10–14 days post-infection [
2]. However, up to 50% of FMD-recovered cattle may harbour virus in their oro-pharyngeal and naso-pharyngeal cavity at 28 or more days post-infection and are known as FMDV-carriers or persistently infected animals [
3,
4]. This asymptomatic carrier state of FMD complicates the control and eradication of the disease. The duration of the FMDV-carrier state may be influenced by a combination of viral and host-factors, and can last from months to years [
5]. Although transmission of FMD virus from domestic animal carriers to susceptible naïve animals has not been demonstrated under experimental conditions [
6,
7], circumstantial evidence from field studies has linked FMDV-carrier cattle to subsequent outbreaks [
8,
9,
10]. Furthermore, a recent experimental study has demonstrated clinical infection in naïve cattle when inoculated with oro-pharyngeal fluids (OPF) obtained from carrier animals [
11]. Since FMDV-carriers may be considered a risk for transmitting infection, they must be identified by post-vaccination serosurveillance to substantiate freedom from infection to regain the “FMD-free status without vaccination” for the purpose of international trade [
12,
13]. Carrier animals persistently infected with FMDV can be identified by detection of the virus in OPF collected with a probang sampling cup. However, recovery of infectious virus or viral genome from such oro-pharyngeal scrapings of FMDV persistently infected cattle is intermittent [
12,
14].
Tests for the detection of antibodies to FMDV non-structural proteins (NSP) have been used for detection of infection in vaccinated animals (DIVA). However, the currently validated NSP antibody tests [
15] may not detect all infected animals within a vaccinated population [
16] and do not distinguish between carriers and those that have eliminated FMDV. Therefore, new NSP tests or alternative tests which can be used either as screening tests or confirmatory tests to the existing NSP tests are needed.
Several studies describe the presence of FMDV-specific mucosal IgA (IgA) in oro-pharyngeal fluid as an indicator of FMDV persistence [
14,
17,
18,
19]. An IgA-ELISA, using saliva samples, to detect FMD carrier cattle following vaccination and challenge exposure has been developed previously [
14]. Although this IgA-ELISA can detect carrier cattle in vaccinated and unvaccinated populations, the non-specific reactions with some saliva samples from uninfected animals have hampered the introduction of the test (unpublished results).
The above findings led us to develop and validate a new mucosal IgA assay, using saliva, nasal and OPF samples, as a confirmatory or screening test for detection of persistently FMDV infected cattle.
4. Discussion
Since FMDV carriers may be considered to pose a risk of passing on infection, to regain the FMD-free status without ongoing vaccination for the purpose of international trade, with or without the use of temporary emergency vaccination, post-outbreak surveillance is required to demonstrate the absence of infection [
29]. Virus isolation and RT-PCR are two established methods to detect virus or viral genome from oro-pharyngeal fluids collected using a metallic probang cup [
30]. Virus isolation is as sensitive as RT-PCR during the early phase of infection, whereas RT-PCR detects more carriers in the later phase of infection in both cattle and sheep [
12,
16]. Even after combining both virus isolation and RT-PCR test results, at all the time points of sampling 100% carriers are not detected from the OPF samples collected sequentially in time [
12], which is a major constraint in FMD surveillance. Besides this, the approach is cumbersome for mass screening after widespread use of emergency vaccines. Therefore, there is a need for alternative or supplementary methods to detect subclinical infection with FMDV in vaccinated herds.
The presence of anti-FMDV IgA antibody in saliva has been described as an indicator of oro-pharyngeal replication of FMDV [
14]. Though an indirect IgA-ELISA using saliva samples to detect FMDV carrier cattle following vaccination and challenge exposure has been developed earlier, the assay was unsatisfactory on its diagnostic parameters. Therefore, an attempt has been made to develop and validate a modified IgA-ELISA targeting an increased specificity. Non-specificity in the assay may be either due to the high content of detached cells, proteases and tissue particles in the saliva samples [
17], or due to the use of tissue culture derived inactivated crude antigen. In order to achieve a better diagnostic specificity, different negative antigen controls (heterologous SAT2 antigen, BHK-21 cell lysate and blocking buffer without any antigen) were evaluated in the newly developed salivary IgA-ELISA. Accordingly, the best format of the IgA-ELISA was selected with respect to both specificity and sensitivity using receiver-operating characteristic (ROC) curve analysis [
31].
Normally, naïve samples are used to estimate the specificity of the diagnostic assay. However, considering the fact that serosurveillence should normally be conducted after emergency vaccination, it was decided to include saliva samples from both naive and vaccinated uninfected animals as a negative population control in ROC analysis for the selection of suitable negative antigen control.
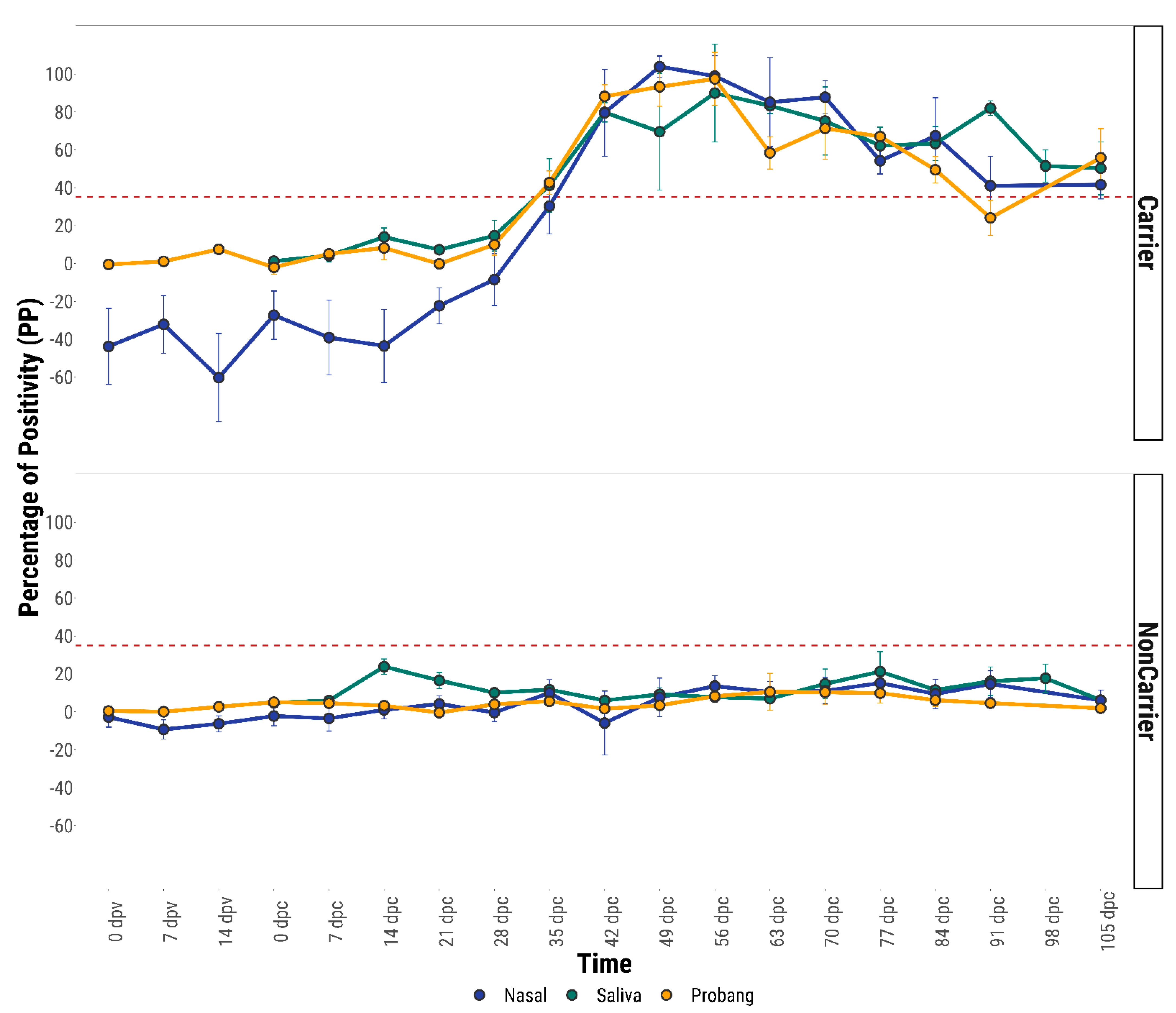
The anti-FMDV IgA response was found to be higher in nasal fluids of FMDV vaccinated and sub-clinically infected carrier animals than saliva and probang fluids. However, both in the acute (for unvaccinated control animals) and in the persistent infection phase (both vaccinated and unvaccinated), the IgA levels were seen to be increased in all three mucosal fluids (saliva, nasal and probang) compared to samples from the vaccinated uninfected, vaccinated recovered and unvaccinated recovered animals. After 28 days post-challenge, the anti-FMDV IgA responses in the nasal and saliva fluids of both vaccinated and unvaccinated carrier animals were found to be significantly higher than those from the non-carriers. The high level of anti-FMDV IgA in the mucosal fluids may be due to the constant stimulation of the local mucosal immune system by persistent FMDV on the dorsal soft palate or naso-pharynx. Furthermore, the presence of IgA antibody on the nasal mucosal surface is a genuine FMD specific antibody response sustained by resident antibody secreting B-cells, rather than plasma transudation [
17].
From the ROC estimations of diagnostic performance, a cut-off value of 35 percentage of positivity (35 PP) was determined for IgA-ELISA using the saliva, nasal and OPF. It was found that nasal IgA-ELISA performed better compared to saliva and OPF IgA-ELISA. With the cut-off value of 35 PP a specificity of 99.11% and sensitivity of 76.53% was obtained for nasal IgA-ELISA by ROC analysis, whilst these parameters were found to increase after Bayesian model fitting (Sp = 99.3% and Se = 85.4%). As this test has similar sensitivity and specificity to the PrioCHECK-NSP test, the IgA assay can be used as screening test for the detection of carrier cattle. The international workshop on serological testing for FMD previously organised in Brescia, Italy, recommended the use of more than one NSP test to increase the overall diagnostic specificity [
13]. In this case, the second confirmatory test should be selected with at least equal specificity and a good sensitivity. As the IgA test detects mucosal antibody and the PrioCHECK-NSP test detects anti-FMDV humoral antibody, using one of the tests for screening and the other as confirmatory test may help to increase the specificity and sensitivity to detect the FMDV carriers. The third option is to run both tests in parallel. In the present study, the concordance of sensitivity between the PrioCHECK-NSP and nasal IgA test was found to be 84.37%. By combining both the nasal-IgA test and PrioCHECK-NSP test in parallel the overall test sensitivity was increased to 96.87%.
Considering that there are no tests with perfect sensitivity and specificity, no sero-surveillance can provide an absolute guarantee of freedom from infection. This applies even to a situation when the whole population has to be tested, e.g., the vaccinated cattle population in an EU country after emergency vaccination. Therefore, serosurveillance should be seen as part of a package of risk mitigation measures that will include movement restrictions, epidemiological tracing and clinical surveillance [
13]. The problem of imperfect test sensitivity can be mitigated to some extent by attributing a status to a herd or population and then applying appropriate disease control measures. A herd could be classified as infected after one animal has reacted positively. However, as no test has a specificity of 100%, many large herds may have to be considered infected due to false positive test results. As this is unacceptable, simultaneous testing by using serological methods with a very high specificity would be needed [
13], albeit this would have the detrimental effect of decreasing the sensitivity. An alternative approach would be the culling of any animal reacting positive without classifying the herd as infected in the absence of further evidence of infection, e.g., clustering or increasing numbers of reactors or epidemiological information. In this case, the requirements for specificity could be lower and a higher sensitivity may be achievable [
13].
During this study, the PrioCHECK NSP assay also scored positive for 18 cattle that were confirmed as non-carrier by VI and RT-PCR testing done at weekly intervals. This result confirms that NSP tests are not specific for the detection of carriers, as NSP antibody responses occur in infected animals that go on to eliminate FMDV. In addition, the NSP test may not be useful for individual animal screening, as cattle that have been vaccinated with high potency vaccine may produce stronger and earlier neutralising antibodies which limit the viral replication and sometimes fail to develop antibodies to NSPs following infection, but these animal could potentially become carriers [
30,
32]. Further in endemic countries where vaccine is not purified from NSP and multiple vaccination is practiced, there is an increased chance of NSP seroconversion without infection.
Although inactivated FMD vaccine, when administered parenterally, stimulates very little or no FMDV specific mucosal immune response [
14,
33], an immediate question arises about the effect of repeated vaccination on the mucosal anti-FMDV IgA response, particularly in the endemic countries where bi-annual prophylactic vaccination is carried out. In order to address this question, the opportunity was taken to test nasal and saliva samples for the detection of anti-FMDV specific antibody in repeatedly vaccinated animals (three times emergency vaccination at 21-day intervals). Anti-FMDV IgA antibody responses in nasal and saliva samples of six cattle remained below the cut-off value after multiple vaccinations, although a sub-threshold peak in IgA response was found for one animal, VC15, after the second vaccination. The anti-FMDV IgA responses in these repeatedly vaccinated animals suggest that the IgA assay may not be subject to non-specificity from multiple vaccinations even in endemic countries, although more animals need to be tested to confirm this. Furthermore, the analysis of serum samples originating from repeatedly vaccinated animals showed an increased humoral immune response of the anti-FMDV NSP antibody following the third vaccination in animals VC14 and VC19. This result supported the earlier finding [
34] where the authors suggested that if animals received multiple doses of FMD vaccine, a small proportion of them might test positive without actual exposure to wild type FMD virus. The detection of NSP antibody in repeatedly vaccinated animals prior to challenge may be due to the presence of trace amounts of contaminating NSP proteins in the commercial FMD vaccine.
In conclusion, from the analysis of anti-FMDV IgA response in mucosal fluids, it is evident that levels of FMDV specific IgA become elevated transiently during the acute phase of infection and were stronger in FMDV carrier animals irrespective of vaccination status. Out of three mucosal fluids, nasal secretion contained the highest level of anti-FMDV IgA antibody. However, a statistically significant difference (p ≤ 0.05) in the anti-FMDV IgA antibody response was found between the three fluids only on 35 dpc (p = 0.03), 42 dpc (0.001) and 63 dpc (p = 0.04) by ANOVA analysis. Both nasal and saliva fluid collection from ruminants are considered as non-invasive methods, whereas collection of OPF is invasive and difficult to do. Moreover, difficulty in collecting uniform OPF samples may contribute to inconsistent virus detection during consecutive probang sampling from known carrier animals. Cattle do not like and usually resist the collection of nasal fluids from their nostrils, whereas collection of saliva is much easier. Therefore, a salivary IgA test might be taken forward in outbreak situation in the field, even though the nasal IgA test has slightly superior test characteristics.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}