
Molecular Investigation of Canine Parvovirus-2 (CPV-2) Outbreak in Nevis Island: Analysis of the Nearly Complete Genomes of CPV-2 Strains from the Caribbean Region


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sampling
2.3. Screening
2.4. Amplification of CPV-2 Genome
2.5. Nucleotide Sequencing
2.6. Sequence Analysis
2.7. GenBank Accession Numbers
3. Results and Discussion
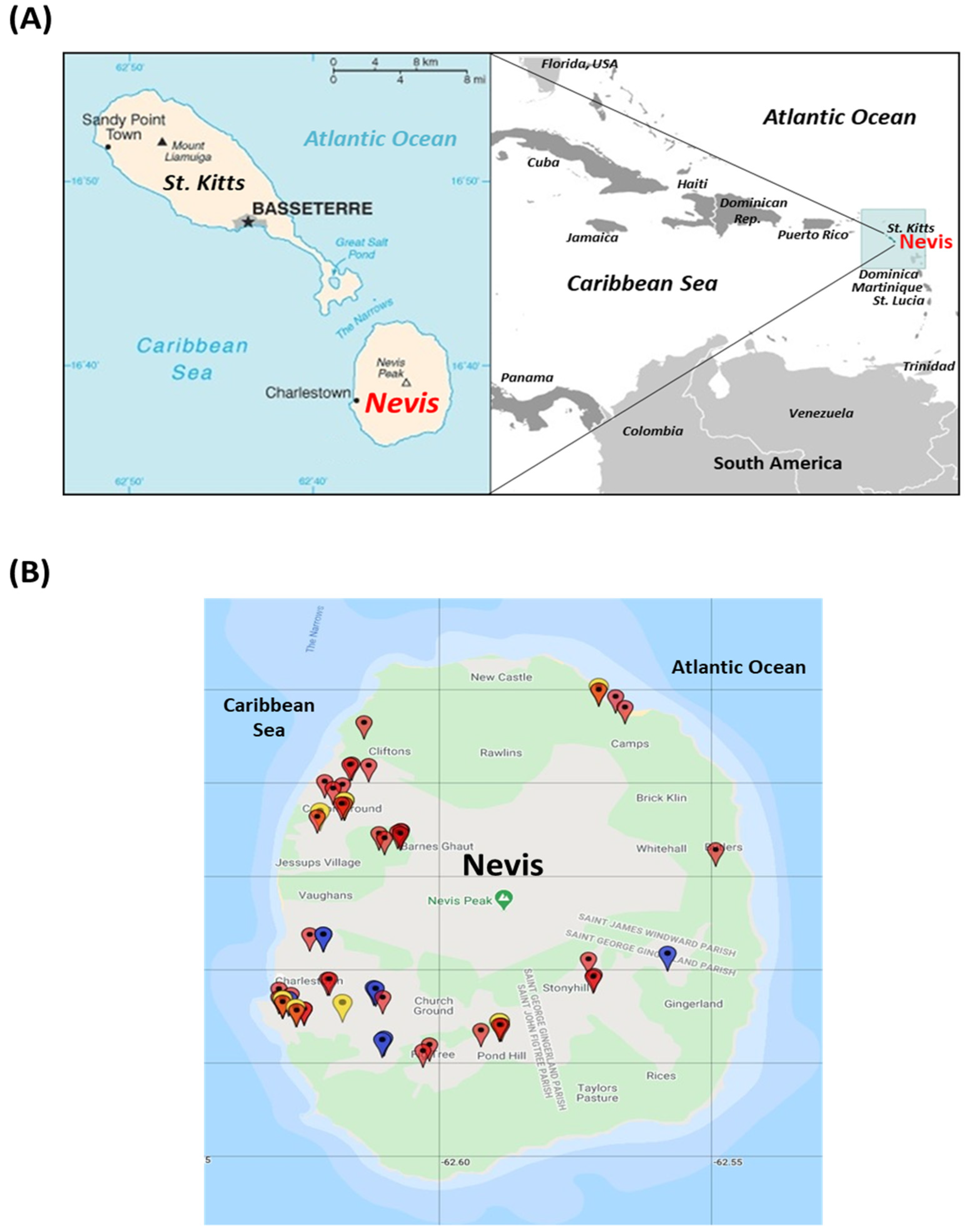
3.1. Molecular Investigation of CPV-2 Outbreak in Nevis Island
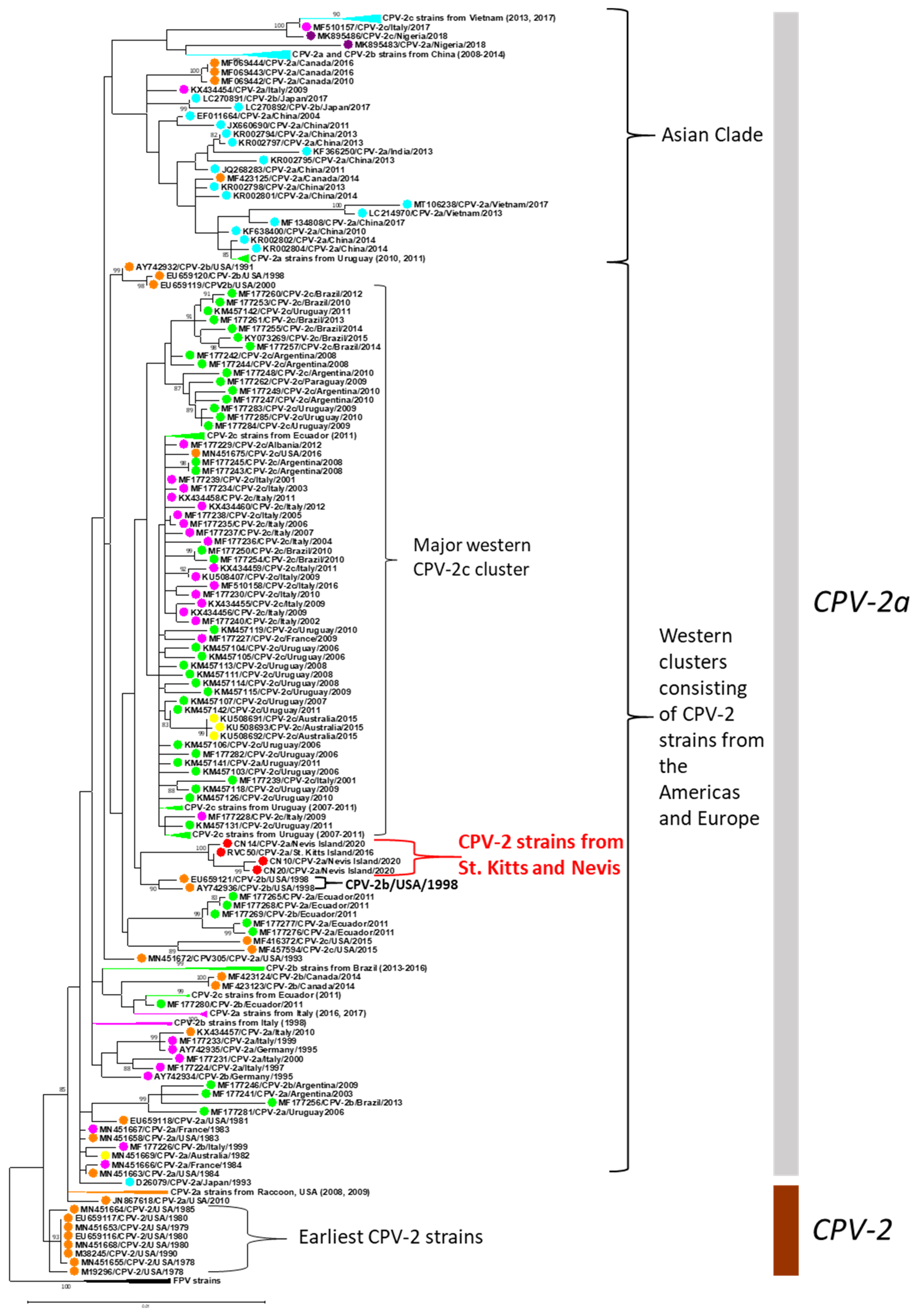
3.2. Analysis of the Nearly Complete Genomes of CPV-2 Strains from St. Kitts and Nevis Islands
3.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine parvovirus—A review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C.; Barrs, V. Canine parvovirus vaccination and immunisation failures: Are we far from disease eradication? Vet. Microbiol. 2020, 247, 108760. [Google Scholar] [CrossRef]
- Mazzaferro, E.M. Update on Canine Parvoviral Enteritis. Vet. Clin. N. Am. Small Anim. Pr. 2020, 50, 1307–1325. [Google Scholar] [CrossRef]
- Reed, A.P.; Jones, E.V.; Miller, T.J. Nucleotide sequence and genome organization of canine parvovirus. J. Virol. 1988, 62, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Niskanen, E.A.; Kalliolinna, O.; Ihalainen, T.O.; Häkkinen, M.; Vihinen-Ranta, M. Mutations in DNA Binding and Transactivation Domains Affect the Dynamics of Parvovirus NS1 Protein. J. Virol. 2013, 87, 11762–11774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niskanen, E.A.; Ihalainen, T.O.; Kalliolinna, O.; Häkkinen, M.M.; Vihinen-Ranta, M. Effect of ATP Binding and Hydrolysis on Dynamics of Canine Parvovirus NS1. J. Virol. 2010, 84, 5391–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Yuan, W.; Davis, I.; Parrish, C.R. Nonstructural Protein-2 and the Replication of Canine Parvovirus. Virology 1998, 240, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mietzsch, M.; Pénzes, J.J.; Mc Kenna, A. Twenty-Five Years of Structural Parvovirology. Viruses 2019, 11, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, J.; Chapman, M.S.; Agbandje, M.; Keller, W.; Smith, K.; Wu, H.; Luo, M.; Smith, T.J.; Rossmann, M.G.; Compans, R.W.; et al. The three-dimensional structure of canine parvovirus and its functional implications. Science 1991, 251, 1456–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hueffer, K.; Parker, J.; Weichert, W.S.; Geisel, R.E.; Sgro, J.-Y.; Parrish, C.R. The Natural Host Range Shift and Subsequent Evolution of Canine Parvovirus Resulted from Virus-Specific Binding to the Canine Transferrin Receptor. J. Virol. 2003, 77, 1718–1726. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.D.; Palermo, L.M.; Hafenstein, S.L.; Parrish, C.R. Different mechanisms of antibody-mediated neutralization of parvoviruses revealed using the Fab fragments of monoclonal antibodies. Virology 2007, 361, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, C.; Thompson, G. Canine parvovirus: The worldwide occurrence of antigenic variants. J. Gen. Virol. 2016, 97, 2043–2057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Zeng, W.; Zhang, X.; Li, S. The genetic evolution of canine parvovirus—A new perspective. PLoS ONE 2017, 12, e0175035. [Google Scholar] [CrossRef] [Green Version]
- Mira, F.; Canuti, M.; Purpari, G.; Cannella, V.; Di Bella, S.; Occhiogrosso, L.; Schirò, G.; Chiaramonte, G.; Barreca, S.; Pisano, P.; et al. Molecular Characterization and Evolutionary Analyses of Carnivore Protoparvovirus 1 NS1 Gene. Viruses 2019, 11, 308. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, I.E.H.; Lee, H.; Allison, A.B.; Lopez-Astacio, R.; Goodman, L.; Oyesola, O.O.; Omobowale, O.; Fagbohun, O.; Dubovi, E.J.; Hafenstein, S.L.; et al. Limited Intrahost Diversity and Background Evolution Accompany 40 Years of Canine Parvovirus Host Adaptation and Spread. J. Virol. 2019, 94, 01162-19. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R. Host range relationships and the evolution of canine parvovirus. Vet. Microbiol. 1999, 69, 29–40. [Google Scholar] [CrossRef]
- Truyen, U.; Evermann, J.F.; Vieler, E.; Parrish, C.R. Evolution of Canine Parvovirus Involved Loss and Gain of Feline Host Range. Virology 1996, 215, 186–189. [Google Scholar] [CrossRef] [Green Version]
- Parrish, C.R.; Aquadro, C.F.; Strassheim, M.L.; Evermann, J.F.; Sgro, J.Y.; Mohammed, H.O. Rapid antigenic-type replacement and DNA sequence evolution of canine parvovirus. J. Virol. 1991, 65, 6544–6552. [Google Scholar] [CrossRef] [Green Version]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for evolution of canine parvovirus type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar] [CrossRef]
- Grecco, S.; Iraola, G.; DeCaro, N.; Alfieri, A.; Alfieri, A.; Calderón, M.G.; Da Silva, A.P.; Name, D.; Aldaz, J.; Calleros, L.; et al. Inter- and intracontinental migrations and local differentiation have shaped the contemporary epidemiological landscape of canine parvovirus in South America. Virus Evol. 2018, 4, vey011. [Google Scholar] [CrossRef] [Green Version]
- Touihri, L.; Bouzid, I.; Daoud, R.; Desario, C.; EL Goulli, A.F.; DeCaro, N.; Ghorbel, A.; Buonavoglia, C.; Bahloul, C. Molecular characterization of canine parvovirus-2 variants circulating in Tunisia. Virus Genes 2008, 38, 249–258. [Google Scholar] [CrossRef]
- Martella, V.; DeCaro, N.; Elia, G.; Buonavoglia, C. Surveillance Activity for Canine Parvovirus in Italy. J. Vet. Med. Ser. B 2005, 52, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.; Nair, R.; Peda, A.; Aung, M.S.; Ashwinie, G.S.; Gallagher, C.A.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Molecular characterization of canine parvovirus and canine enteric coronavirus in diarrheic dogs on the island of St. Kitts: First report from the Caribbean region. Virus Res. 2017, 240, 154–160. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Yip, H.Y.E.; Peaston, A.; Woolford, L.; Khuu, S.J.; Wallace, G.; Kumar, R.S.; Patel, K.; Azari, A.A.; Akbarzadeh, M.; Sharifian, M.; et al. Diagnostic Challenges in Canine Parvovirus 2c in Vaccine Failure Cases. Viruses 2020, 12, 980. [Google Scholar] [CrossRef]
- De Oliveira, P.S.B.; Cargnelutti, J.F.; Masuda, E.K.; Weiblen, R.; Flores, E.F. New variants of canine parvovirus in dogs in southern Brazil. Arch. Virol. 2019, 164, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Castillo, C.; Neira, V.; Aniñir, P.; Grecco, S.; Pérez, R.; Panzera, Y.; Zegpi, N.-A.; Sandoval, A.; Sandoval, D.; Cofre, S.; et al. First Molecular Identification of Canine Parvovirus Type 2 (CPV2) in Chile Reveals High Occurrence of CPV2c Antigenic Variant. Front. Vet. Sci. 2020, 7, 194. [Google Scholar] [CrossRef]
- De La Torre, D.; Mafla, E.; Puga, B.; Erazo, L.; Astolfi-Ferreira, C.; Ferreira, A.P. Molecular characterization of canine parvovirus variants (CPV-2a, CPV-2b, and CPV-2c) based on the VP2 gene in affected domestic dogs in Ecuador. Vet. World 2018, 11, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Maya, L.; Calleros, L.; Francia, L.; Hernández, M.; Iraola, G.; Panzera, Y.; Sosa, K.; Pérez, R. Phylodynamics analysis of canine parvovirus in Uruguay: Evidence of two successive invasions by different variants. Arch. Virol. 2013, 158, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Ramirez, S.; Rendon-Marin, S.; Ruiz-Saenz, J. Phylogenetic, Evolutionary and Structural Analysis of Canine Parvovirus (CPV-2) Antigenic Variants Circulating in Colombia. Viruses 2020, 12, 500. [Google Scholar] [CrossRef]
- Quispe, R.Q.; Espinoza, L.L.; Beltrán, R.R.; Alcántara, R.H.R.; Hernández, L.M. Canine parvovirus types 2a and 2c detection from dogs with suspected parvoviral enteritis in Peru. Virus Dis. 2018, 29, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Cooper, E.; Lamm, C.; Murray, B.; Rezabek, G.; Johnston, L.; Campbell, G.; Johnson, B. Canine Parvovirus Types 2c and 2b Circulating in North American Dogs in 2006 and 2007. J. Clin. Microbiol. 2007, 45, 4044–4047. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.; Murphy, M.; Hueffer, K.; Chen, J. Investigation of a Canine Parvovirus Outbreak using Next Generation Sequencing. Sci. Rep. 2017, 7, 9633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, C.A.; Allard, V.; Cloutier, G. Canine parvovirus type 2b is the most prevalent genomic variant strain found in parvovirus antigen positive diarrheic dog feces samples across Canada. Can. Vet. J. 2016, 57, 29–31. [Google Scholar]
- Faz, M.; Martínez, J.S.; Gómez, L.B.; Quijano-Hernández, I.; Fajardo, R.; Del Ángel-Caraza, J. Origin and genetic diversity of canine parvovirus 2c circulating in Mexico. Arch. Virol. 2018, 164, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Kwan, E.; Carrai, M.; Lanave, G.; Hill, J.; Parry, K.; Kelman, M.; Meers, J.; Decaro, N.; Beatty, J.A.; Martella, V.; et al. Analysis of canine parvoviruses circulating in Australia reveals predominance of variant 2b and identifies feline parvovirus-like mutations in the capsid proteins. Transbound. Emerg. Dis. 2021, 68, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.A.; Chandrasekar, V.; Hébert, B.; Sullivan, G.M.; Rossmann, M.G.; Parrish, C.R. Host range and variability of calcium binding by surface loops in the capsids of canine and feline parvoviruses. J. Mol. Biol. 2000, 300, 597–610. [Google Scholar] [CrossRef] [Green Version]
- Organtini, L.J.; Allison, A.B.; Lukk, T.; Parrish, C.R.; Hafenstein, S. Global Displacement of Canine Parvovirus by a Host-Adapted Variant: Structural Comparison between Pandemic Viruses with Distinct Host Ranges. J. Virol. 2014, 89, 1909–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Buonavoglia, C. Canine parvovirus post-vaccination shedding: Interference with diagnostic assays and correlation with host immune status. Vet. J. 2017, 221, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Freisl, M.; Speck, S.; Truyen, U.; Reese, S.; Proksch, A.-L.; Hartmann, K. Faecal shedding of canine parvovirus after modified-live vaccination in healthy adult dogs. Vet. J. 2017, 219, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.B.; Kohler, D.J.; Ortega, A.; Hoover, E.A.; Grove, D.M.; Holmes, E.C.; Parrish, C.R. Host-Specific Parvovirus Evolution in Nature Is Recapitulated by In Vitro Adaptation to Different Carnivore Species. PLoS Pathog. 2014, 10, e1004475. [Google Scholar] [CrossRef] [PubMed]
- Manh, T.N.; Piewbang, C.; Rungsipipat, A.; Techangamsuwan, S. Molecular and phylogenetic analysis of Vietnamese canine parvovirus 2C originated from dogs reveals a new Asia-IV clade. Transbound. Emerg. Dis. 2021, 68, 1445–1453. [Google Scholar] [CrossRef]
- Kleymann, A.; Becker, A.A.M.J.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Detection and Molecular Characterization of Picobirnaviruses (PBVs) in the Mongoose: Identification of a Novel PBV Using an Alternative Genetic Code. Viruses 2020, 12, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, M.D.; Henriques, A.M.; Barros, S.C.; Fagulha, T.; Mendonça, P.; Carvalho, P.; Monteiro, M.; Fevereiro, M.; Basto, M.P.; Rosalino, L.M.; et al. Snapshot of Viral Infections in Wild Carnivores Reveals Ubiquity of Parvovirus and Susceptibility of Egyptian Mongoose to Feline Panleukopenia Virus. PLoS ONE 2013, 8, e59399. [Google Scholar] [CrossRef]
- Steinel, A.; Parrish, C.R.; Bloom, M.E.; Truyen, U. Parvovirus Infections in Wild Carnivores. J. Wildl. Dis. 2001, 37, 594–607. [Google Scholar] [CrossRef]
- Suspected Outbreak of Virus among Statia Dogs. Available online: https://www.thedailyherald.sx/islands/suspected-outbreak-of-virus-among-statia-dogs (accessed on 25 April 2021).
- St Lucia Seeing Alarming Increase in Deadly Canine Parvovirus. Available online: https://www.loopslu.com/content/st-lucia-seeing-alarming-increase-deadly-canine-parvovirus (accessed on 25 April 2021).



{kind=link}
{kind=link}
| Sample/Strain | Date of Sampling | Age | Sex | Breed | Vaccination against CPV-2 1 | SNAP® Parvo Test 2 | PCR 3 | GenBank Accession Number (VP2 Gene, Complete ORF) | CPV-2 Variant |
|---|---|---|---|---|---|---|---|---|---|
| CN1 | 1 August 2020 | 3 months | Female | Shih Tzu | None | Positive | Not performed | Not sequenced | Not determined |
| CN2 | 11 August 2020 | 4 months | Male | Bulldog | Incomplete 4 | Positive | Not performed | Not sequenced | Not determined |
| CN3 | 14 August 2020 | 2 months | Male | Pit bull | None | Positive | Not performed | Not sequenced | Not determined |
| CN4 | 15 August 2020 | 3 months 15 days | Male | Pit bull | None | Positive | Not performed | Not sequenced | Not determined |
| CN5 | 15 August 2020 | 4 months | Female | Pit bull | None | Positive | Not performed | Not sequenced | Not determined |
| CN6 | 21 August 2020 | 3 months | Female | Pit bull | Not available | Positive | Not performed | Not sequenced | Not determined |
| CN7 | 22 August 2020 | 2 months | Male | Island mix 5 | None | Positive | Positive | MW595661 | New CPV-2a 6 |
| CN8 | 24 August 2020 | 2 months 15 days | Male | Pit bull | None | Negative | Positive | MW595662 | New CPV-2a |
| CN9 | 25 August 2020 | 2 months | Female | Pit bull | None | Positive | Not performed | Not sequenced | Not determined |
| CN10 | 26 August 2020 | 4 months | Male | Island mix | None | Positive | Positive | MW595663 | New CPV-2a |
| CN11 | 28 August 2020 | 4 months 15 days | Male | Island mix | Incomplete 4 | Positive | Positive | MW595664 | New CPV-2a |
| CN12 | 28 August 2020 | 7 months | Male | Pit bull | None | Positive | Positive | MW595665 | New CPV-2a |
| CN13 | 28 August 2020 | 2 year | Male | Rottweiler mix | None | Negative | Positive | MW595666 | New CPV-2a |
| CN14 | 28 August 2020 | 7 months | Female | Island mix | Not available | Positive | Positive | MW595667 | New CPV-2a |
| CN15 | 29 August 2020 | 2 months 15 days | Female | Island mix | None | Negative | Positive | MW595668 | New CPV-2a |
| CN16 | 1 September 2020 | 10 months | Male | Island mix | Complete | Positive | Positive | MW595669 | New CPV-2a |
| CN17 | 1 September 2020 | 2 months | Male | Bulldog x Mastiff | None | Positive | Positive | MW595670 | New CPV-2a |
| CN18 | 1 September 2020 | 4 months | Male | Island mix | None | Negative | Positive | MW595671 | New CPV-2a |
| CN19 | 1 September 2020 | 4 months | Female | Pit bull | Not available | Negative | Positive | MW595672 | New CPV-2a |
| CN20 | 1 September 2020 | 4 months | Male | Pit bull | Not available | Positive | Positive | MW595673 | New CPV-2a |
| CN21 | 2 September 2020 | 1 year | Female | Island mix | None | Negative | Positive | MW595674 | New CPV-2a |
| CN22 | 3 September 2020 | 6 months | Male | Island mix | Complete | Positive | Positive | MW595675 | New CPV-2a |
| CN23 | 3 September 2020 | 2 months | Female | Pit bull | None | Negative | Positive | MW595676 | New CPV-2a |
| CN24 | 3 September 2020 | 4 months | Female | Not available | None | Negative | Positive | MW595677 | New CPV-2a |
| CN25 | 4 September 2020 | 2 months | Female | Island mix | None | Negative | Positive | MW595678 | New CPV-2a |
| CN26 | 4 September 2020 | 2 months | Female | Island mix | None | Negative | Positive | MW595679 | New CPV-2a |
| CN27 7 | 5 September 2020 | 1 month 15 days | Male | Great Dane | None | Negative | Positive | MW595680 | New CPV-2a |
| CN28 | 8 September 2020 | 6 months | Male | Pit bull | None | Positive | Positive | MW595681 | New CPV-2a |
| CN29 | 11 September 2020 | 2 months | Male | Pit bull | None | Positive | Positive | MW595682 | New CPV-2a |
| CN30 | 13 September 2020 | 2 months | Male | Pit bull | None | Positive | Positive | MW595683 | New CPV-2a |
| CN31 | 14 September 2020 | 7 months | Female | Pit bull mix | Complete | Positive | Positive | MW595684 | New CPV-2a |
| CN32 | 25 September 2020 | 2 months | Female | Island mix | None | Negative | Positive | MW595685 | New CPV-2a |
| CN33 | 29 September 2020 | 2 months 15 days | Female | Island mix | None | Positive | Positive | MW595686 | New CPV-2a |
| CN34 | 7 October 2020 | 7 months | Male | Island mix | None | Positive | Positive | MW595687 | New CPV-2a |
| CN35 | 8 October 2020 | 4 months | Female | Island mix | Incomplete 4 | Positive | Positive | MW595688 | New CPV-2a |
| CN36 | 8 October 2020 | 4 months | Male | Island mix | None | Negative | Positive | MW595689 | New CPV-2a |
| CN37 | 9 October 2020 | 4 months | Male | Island mix | None | Negative | Positive | MW595690 | New CPV-2a |
| CN38 | 9 October 2020 | 2 months | Male | Island mix | Incomplete 4 | Negative | Positive | MW595691 | New CPV-2a |
| CN39 | 17 October 2020 | 1 month 15 days | Male | Island mix | None | Positive | Positive | MW595692 | New CPV-2a |
| CN40 7 | 5 November 2020 | 3 months 15 days | Male | Great Dane | None | Negative | Positive | MW595693 | New CPV-2a |
| Amino Acid Position | 87 | 101 | 262 | 267 | 297 | 300 | 305 | 321 | 324 | 373 | 375 | 426 | 440 | 555 | 570 | Variant |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain/Place/Year | ||||||||||||||||
| CPV-b/USA/1978 | Met | Ile | Ala | Phe | Ser | Ala | Asp | Asn | Tyr | Asp | Asn | Asn | Thr | Val | Lys | CPV-2 |
| CPV-15/USA/1984 | Leu | Thr | Ala | Phe | Ser | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Ile | Lys | CPV-2a |
| CPV-39/USA/1984 | Leu | Thr | Ala | Phe | Ser | Gly | Tyr | Asn | Tyr | Asp | Asp | Asp | Thr | Val | Lys | CPV-2b |
| 219/08-13/ITA/2008 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Glu | Thr | Val | Lys | CPV-2c |
| CPV-435/USA/2003 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| RVC50/St. Kitts/2016 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| GX304/CHN/2017 | Leu | Thr | Thr | Tyr | Ala | Gly | Tyr | Asn | Ile | Asp | Asp | Asn | Ala | Val | Lys | New CPV-2a |
| Beaumaris/AUS/2017 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Ile | Asn | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CPV-436/USA/2003 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asp | Thr | Val | Lys | New CPV-2b |
| CN7/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN8/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN10/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN11/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN12/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN13/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN14/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asn | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN15/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN16/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN17/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN18/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN19/Nevis/2020 | Leu | Thr | Thr | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN20/Nevis/2020 | Leu | Thr | Thr | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN21/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN22/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN23/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asn | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN24/Nevis/2020 | Leu | Thr | Thr | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN25/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asn | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN26/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN27/Nevis/20201 | Leu | Thr | Thr | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN28/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN29/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN30/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN31/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN32/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN33/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN34/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN35/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN36/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN37/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN38/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN39/Nevis/2020 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| CN40/Nevis/20201 | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Asn | Tyr | Asp | Asp | Asn | Thr | Val | Lys | New CPV-2a |
| VANGUARD/vaccine | Arg | Ile | Ala | Phe | Ser | Ala | Asp | Asn | Tyr | Asp | Glu | Asn | Thr | Val | Lys | CPV-2 |
| Duramune/vaccine | Leu | Thr | Ala | Phe | Ala | Gly | Tyr | Lys | Tyr | Asp | Asp | Asp | Thr | Val | Glu | New CPV-2b |
| Nt Position 1 | CPV-2 Strain (Year/Place of Detection) | Nt Change → Translational Effect | Coding Region | ||||
|---|---|---|---|---|---|---|---|
| CPV-12 (1978/USA) | RVC50 (2016/St. Kitts) | CN10 (2020/Nevis) | CN14 (2020/Nevis) | CN20 (2020/Nevis) | |||
| 516 | G | A | A | A | A | G516A → Synonymous | Within the NS coding region |
| 562 | T | C | C | C | C | T562C → Synonymous | |
| 726 | G | G | A | G | A | G726A → Synonymous | |
| 753 | A | A | G | A | G | A753G 2 → Synonymous | |
| 1104 | T | T | C | T | C | T1104C 2 → Synonymous | |
| 1164 | A | A | G | A | G | A1164G → Synonymous | |
| 1209 | T | C | C | C | C | T1209C → Synonymous | |
| 1305 | T | C | C | C | C | T1305C 2 → Synonymous | |
| 1623 | A | G | G | A | G | A1623G → Synonymous | |
| 1752 | A | G | G | G | G | A1752G → NS2 Thr94Ala | |
| 1923 | G | A | A | A | A | G1923A → NS2 Asp151Asn | |
| 1926 | A | G | G | G | G | A1926G → NS2 Met152Val | |
| 1975 | T | C | C | C | C | T1975C → Synonymous | |
| 2086 | A | G | G | G | G | A2086G → VP1 Intron | |
| 2154 | G | A | A | A | A | G2154A → Synonymous | Within the VP coding region |
| 2436 | A | A | G | A | G | A2436G 2 → Synonymous | |
| 2574 | T | A | A | A | A | T2754A → Synonymous | |
| 2773 | A | T | T | T | T | A2773T → VP2 Met87Leu 3 | |
| 2816 | T | C | C | C | C | T2816C → VP2 Ile101Thr 3 | |
| 2923 | G | A | A | A | A | G2923A → Synonymous | |
| 2940 | A | G | G | G | G | A2940G → Synonymous | |
| 3006 | C | T | T | T | T | C3006T → Synonymous | |
| 3039 | T | T | G | T | T | T3039G 2 → Synonymous | |
| 3117 | A | A | G | A | A | A3117G → Synonymous | |
| 3297 | G | G | G | G | A | G3297A → VP2 Ala262Thr | |
| 3403 | T | G | G | G | G | T3403G → VP2 Ser297Ala | |
| 3413 | C | G | G | G | G | C3413G → VP2 Ala300Gly 3 | |
| 3427 | G | T | T | T | T | G3427T → VP2 Asp305Tyr 3 | |
| 3582 | T | C | C | C | C | T3582C → Synonymous | |
| 3631 | G | G | G | A | G | G3631A 2 → VP2 Asp373Asn | |
| 3637 | A | G | G | G | G | A3637G → VP2 Asn375Asp 3 | |
| 3702 | G | G | G | A | G | G3702A → Synonymous | |
| 3894 | A | G | G | G | G | A3894G → Synonymous | |
| 4017 | A | G | G | G | G | A4017G → Synonymous | |
| 4030 | T | C | C | C | C | T4030C → Synonymous | |
| Amino Acid Residue | NS1 | NS2 | VP2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 60 | 544 | 545 | 572 | 630 | 152 | 262 | 267 | 324 | 373 | 426 | |||
| Clade 1 | |||||||||||||
| CPV-2 origin | Ile | Tyr | Glu | Glu | Leu | Met | Ala | Phe | Tyr | Asp | Asn | ||
| Asian | Ile/Val | Phe/Tyr | Glu/Val | Lys | Leu/Pro | Val | Ala | Phe/Tyr | Ile/Tyr | Asp | Asn/Asp/Glu | ||
| Western | Ile | Phe/Tyr | Glu | Glu | Leu | Met/Val | Ala | Phe | Leu/Tyr | Asp | Asn/Asp/Glu | ||
| Strain/Place/Year | |||||||||||||
| CN10/Nevis/2020 | Ile | Tyr | Glu | Glu | Leu | Val | Ala | Phe | Tyr | Asp | Asn | ||
| CN14/Nevis/2020 | Ile | Tyr | Glu | Glu | Leu | Val | Ala | Phe | Tyr | Asn 3 | Asn | ||
| CN20/Nevis/2020 | Ile | Tyr | Glu | Glu | Leu | Val | Thr 2 | Phe | Tyr | Asp | Asn | ||
| RVC50/St. Kitts/2016 | Ile | Tyr | Glu | Glu | Leu | Val | Ala | Phe | Tyr | Asp | Asn | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gainor, K.; Bowen, A.; Bolfa, P.; Peda, A.; Malik, Y.S.; Ghosh, S. Molecular Investigation of Canine Parvovirus-2 (CPV-2) Outbreak in Nevis Island: Analysis of the Nearly Complete Genomes of CPV-2 Strains from the Caribbean Region. Viruses 2021, 13, 1083. https://doi.org/10.3390/v13061083
Gainor K, Bowen A, Bolfa P, Peda A, Malik YS, Ghosh S. Molecular Investigation of Canine Parvovirus-2 (CPV-2) Outbreak in Nevis Island: Analysis of the Nearly Complete Genomes of CPV-2 Strains from the Caribbean Region. Viruses. 2021; 13(6):1083. https://doi.org/10.3390/v13061083
Chicago/Turabian StyleGainor, Kerry, April Bowen, Pompei Bolfa, Andrea Peda, Yashpal S. Malik, and Souvik Ghosh. 2021. "Molecular Investigation of Canine Parvovirus-2 (CPV-2) Outbreak in Nevis Island: Analysis of the Nearly Complete Genomes of CPV-2 Strains from the Caribbean Region" Viruses 13, no. 6: 1083. https://doi.org/10.3390/v13061083
APA StyleGainor, K., Bowen, A., Bolfa, P., Peda, A., Malik, Y. S., & Ghosh, S. (2021). Molecular Investigation of Canine Parvovirus-2 (CPV-2) Outbreak in Nevis Island: Analysis of the Nearly Complete Genomes of CPV-2 Strains from the Caribbean Region. Viruses, 13(6), 1083. https://doi.org/10.3390/v13061083