
Genetic and Pathogenic Characterization of QX(GI-19)-Recombinant Infectious Bronchitis Viruses in South Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Viral RNA Extraction, RT-PCR, and Sequencing
2.3. Phylogenetic Analysis, Sequence Comparisons, and Recombination Analysis
2.4. Pathogenicity Testing
2.5. Statistical Analysis
3. Results
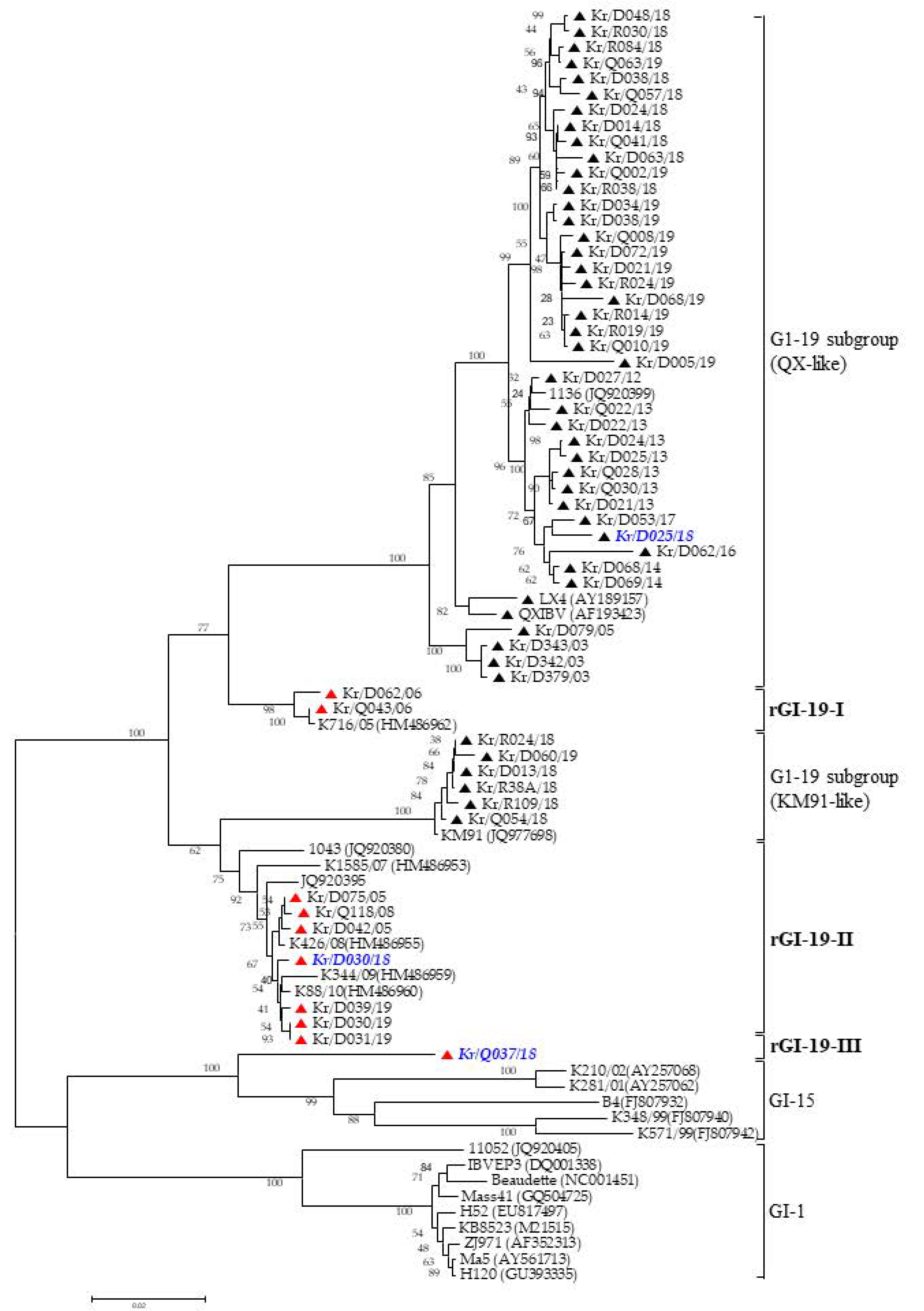
3.1. Phylogenetic Analysis
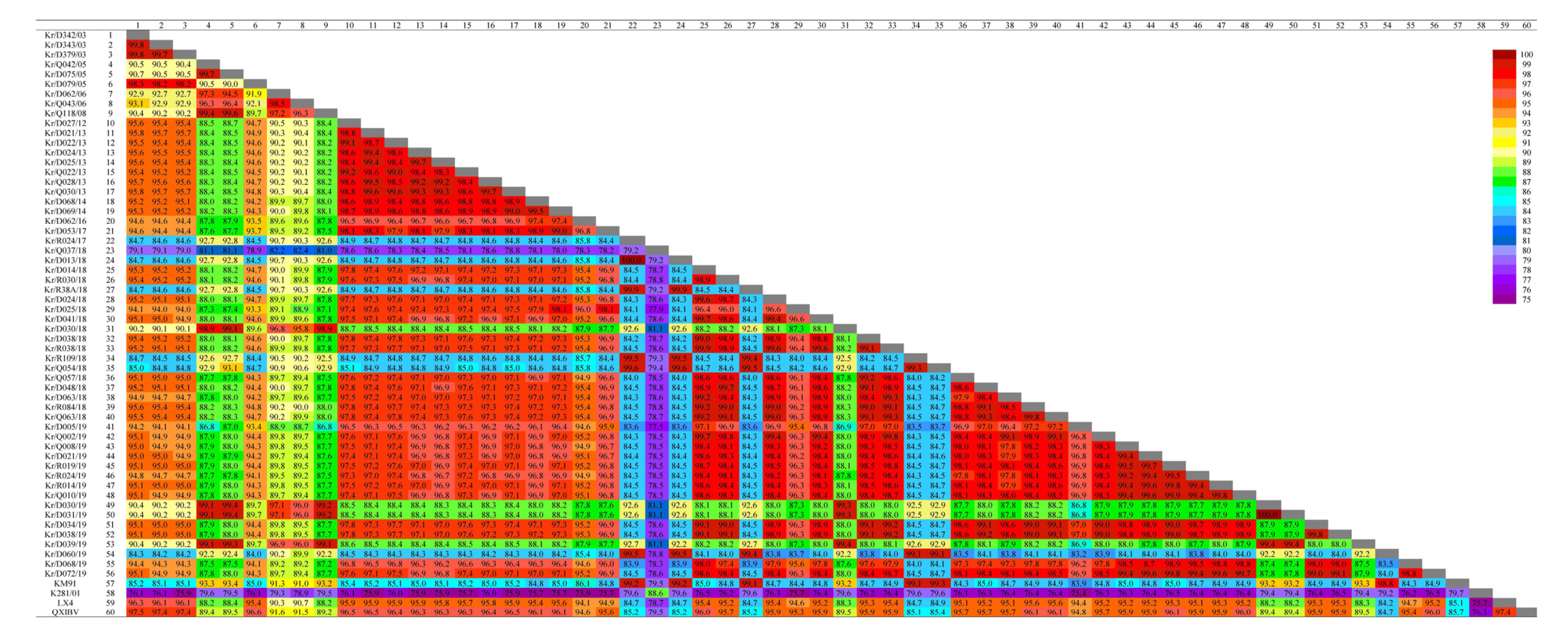
3.2. Sequence Comparisons
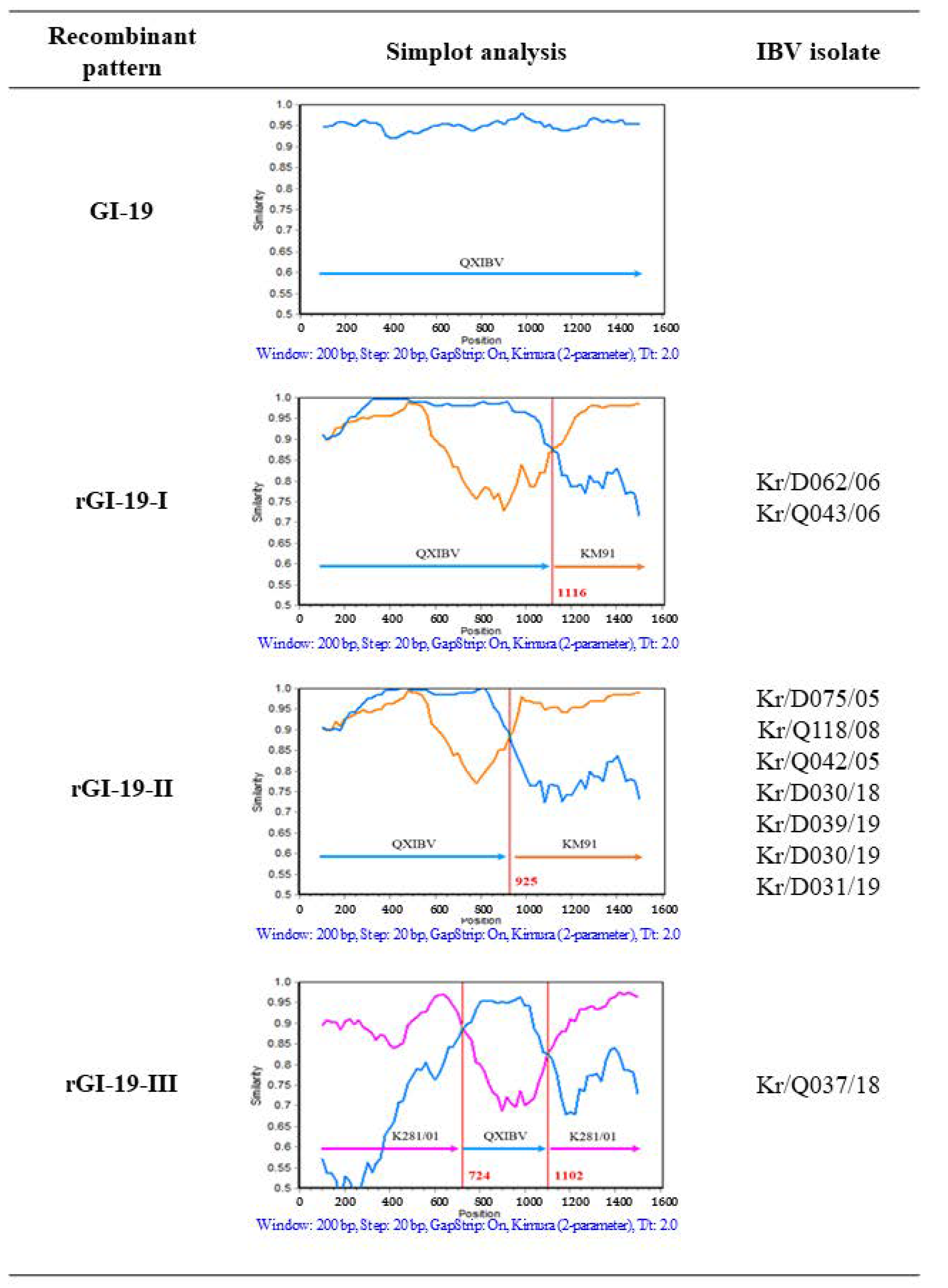
3.3. Recombination Analysis
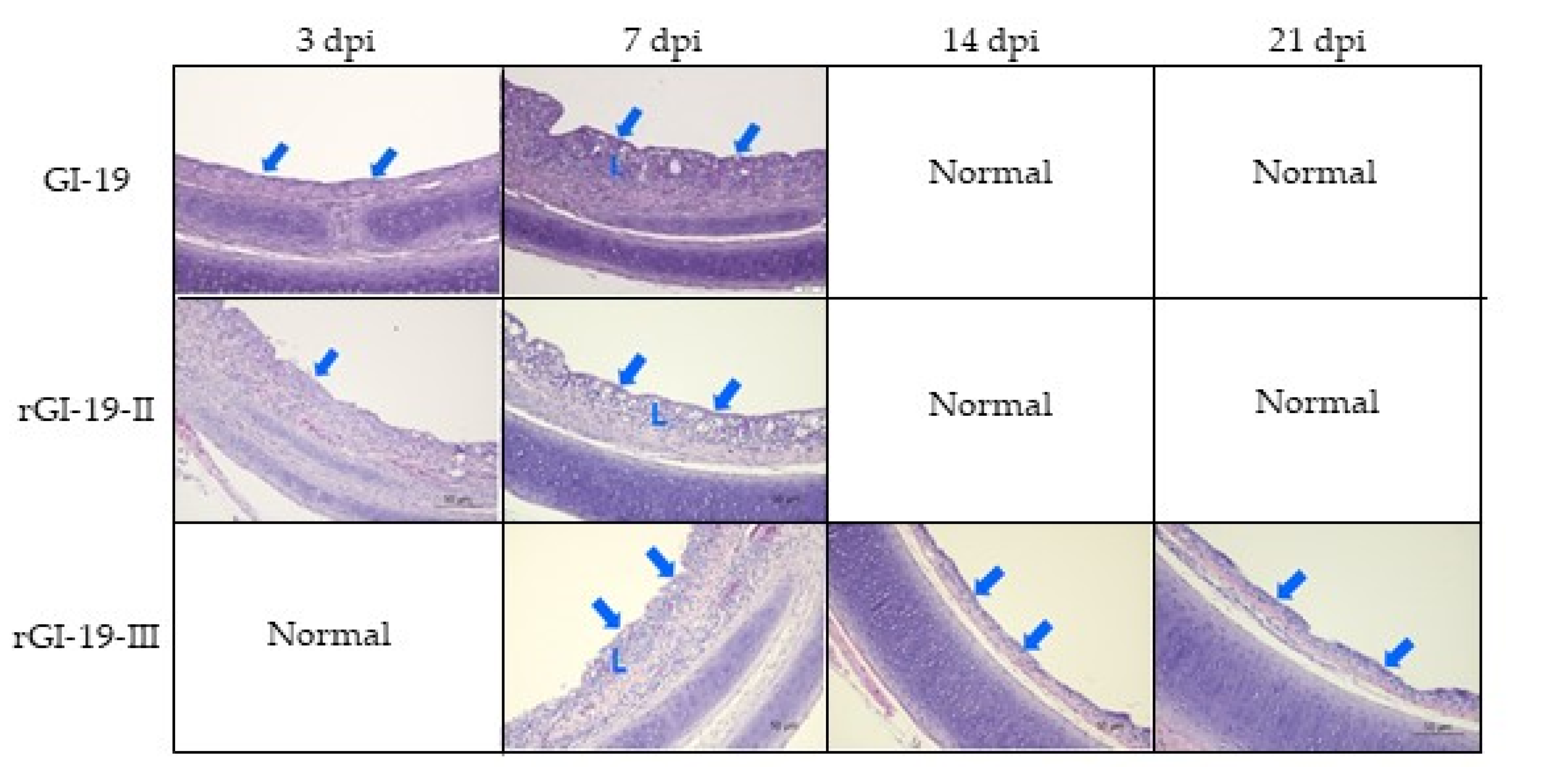
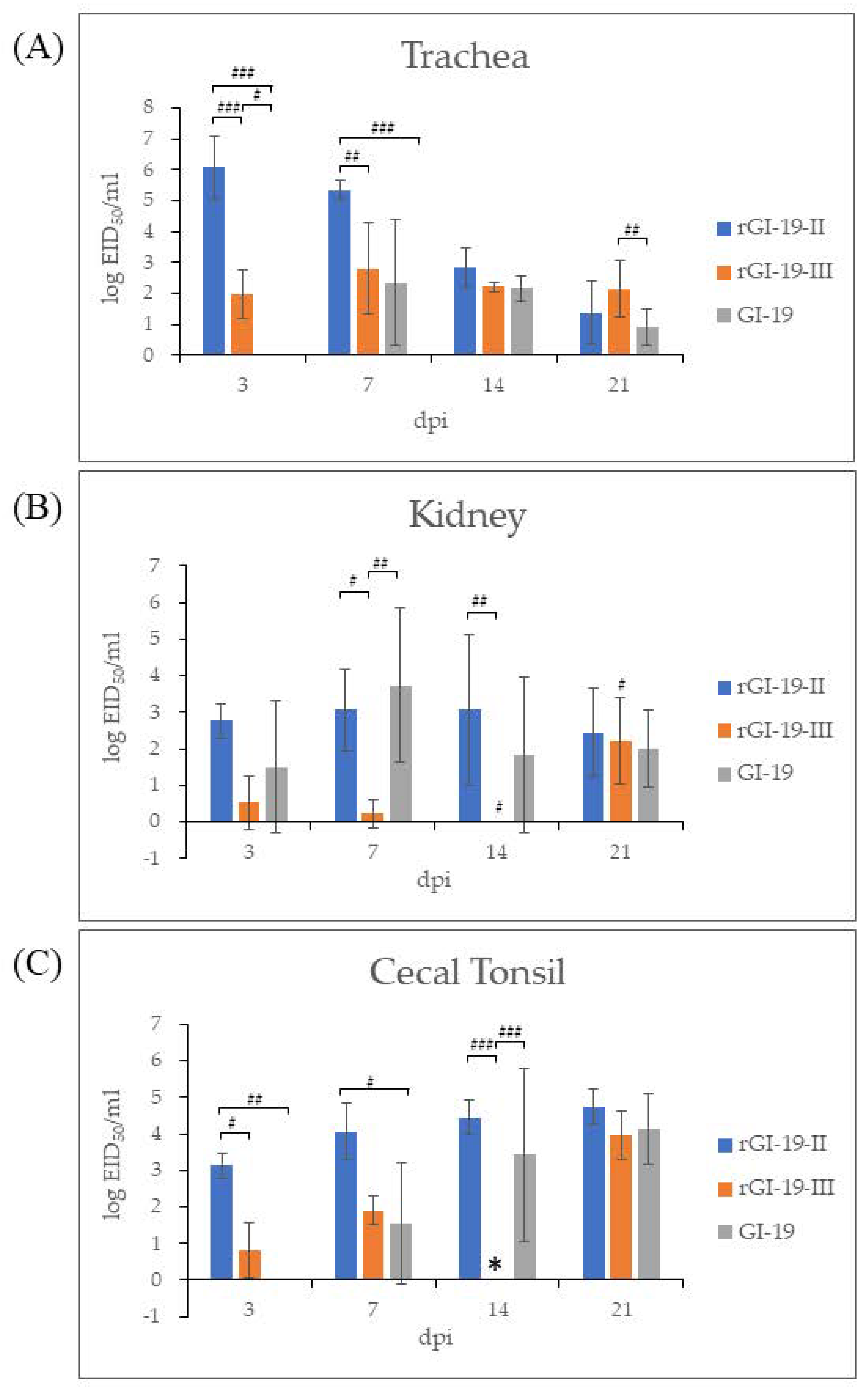
3.4. Animal Experiments and Histopathology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abu-Akkada, S.S.; Awad, A.M. Isolation, propagation, identification and comparative pathogenicity of five Egyptian field strains of Eimeria tenella from broiler chickens in five different provinces in Egypt. Res. Vet. Sci. 2012, 92, 92–95. [Google Scholar] [CrossRef]
- Colvero, L.; Villarreal, L.; Torres, C.; Brandao, P. Assessing the economic burden of avian infectious bronchitis on poultry farms in Brazil. Rev. Sci. Tech. 2015, 34, 993–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Jiang, Y.; Low, S.; Wang, Z.; Nam, S.J.; Liu, W.; Kwangac, J. Characterizatioin of three infectious bronchitis virus isolates from China associated with proventriculus in vaccinated chickens. Avian Dis. 2001, 45, 416–424. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, M.; Tian, X.; Shao, H.; Qian, K.; Ye, J.; Qin, A. Identification of a novel recombinant virulent avian infectious bronchitis virus. Veter. Microbiol. 2017, 199, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant Avian Infectious Bronchitis Virus Expressing a Heterologous Spike Gene Demonstrates that the Spike Protein Is a Determinant of Cell Tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, D.; Davia, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus IBV: Virus retaining spike glycopolypeptide S2 but not S1 is unable to induce virus-neutralizing or haemagglutination-inhibiting antibody, or induce chicken tracheal protection. J. Gen. Virol. 1986, 67, 1435–1442. [Google Scholar] [CrossRef]
- Pohuang, T.; Sasipreeyajan, J. The pathogenesis of a new variant genotype and GI-19-like infectious bronchitis virus isolated from chickens in Thailand. Thai J. Vet. Med. 2012, 42, 51–57. [Google Scholar]
- Parsons, L.M.; Bouwman, K.M.; Azurmendi, H.; de Vries, R.; Cipollo, J.F.; Verheije, M.H. Glycosylation of the viral attachment protein of avian coronavirus is essential for host cell and receptor binding. J. Biol. Chem. 2019, 294, 7797–7809. [Google Scholar] [CrossRef] [Green Version]
- Promkuntod, N.; van Eijndhoven, R.; de Vrieze, G.; Gröne, A.; Verheije, M. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Shan, D.; Fang, S.; Han, Z.; Ai, H.; Zhao, W.; Chen, Y.; Jiang, L.; Liu, S. Effects of hypervariable regions in spike protein on pathogenicity, tropism, and serotypes of infectious bronchitis virus. Virus Res. 2018, 250, 104–113. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.-Y.; Zhao, Y.; Chen, Y.; Zhao, J.; Zhang, G.-Z. Characterization and analysis of an infectious bronchitis virus strain isolated from southern China in 2013. Virol. J. 2016, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Chen, Y.; Zhao, J.; Xu, G.; Zhao, Y.; Zhang, G. Pathogenicity of a TW-like strain of infectious bronchitis virus and evaluation of the protection induced against it by a GI-19-Like strain. Front. Microbiol. 2016, 7, 1653. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.Y.; Chen, T.; Zhang, X.; Shao, G.M.; Cao, Y.; Chen, D.K.; Lin, W.C.; Chen, F.; Xie, Q.M. Molecular characteristic and pathogenicity analysis of a virulent recombinant avian infectious bronchitis virus isolated in China. Poult. Sci. 2018, 97, 3519–3531. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W. Reviw of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.-K.; Jeon, W.-J.; Lee, Y.-J.; Jeong, O.-M.; Choi, J.-G.; Kwon, J.-H.; Choi, K.-S. Genetic Diversity of Avian Infectious Bronchitis Virus Isolates in Korea Between 2003 and 2006. Avian Dis. 2008, 52, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.S.; Lee, E.K.; Jeon, W.J.; Park, M.J.; Kim, J.W.; Kwon, J.H. Pathogenicity and antigenicity of a new variant of Korean nephropathogenic infectious bronchitis virus. J. Vet. Sci. 2009, 10, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Na Youn, H.; Kwon, J.S.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine 2010, 28, 2887–2894. [Google Scholar] [CrossRef]
- Lim, T.-H.; Lee, H.-J.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; et al. An emerging recombinant cluster of nephropathogenic strains of avian infectious bronchitis virus in Korea. Infect. Genet. Evol. 2011, 11, 678–685. [Google Scholar] [CrossRef]
- Le, T.B.; Lee, H.-J.; Le, V.P.; Choi, K.-S. Multiple Genotypes of Avian Infectious Bronchitis Virus Circulating in Vietnam. Korean J. Poult. Sci. 2019, 46, 127–136. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Intersubtype Recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Dolz, R.; Vergara-Alert, J.; Pérez, M.; Pujols, J.; Majó, N. New insights on infectious bronchitis virus pathogenesis: Characterization of Italy 02 serotype in chicks and adult hens. Veter. Microbiol. 2012, 156, 256–264. [Google Scholar] [CrossRef]
- Callison, S.A.; Hilt, D.A.; Boynton, T.O.; Sample, B.F.; Robison, R.; Swayne, D.E.; Jackwood, M.W. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J. Virol. Methods 2006, 138, 60–65. [Google Scholar] [CrossRef]
- Sun, C.; Han, Z.; Ma, H.; Zhang, Q.; Yan, B.; Shao, Y.; Shao, Y.; Xu, J.; Kong, X.; Liu, S. Phylogenetic analysis of infectious bronchitis coronaviruses newly isolated in China, and pathogenicity and evaluation of protection induced by GI-1achusetts serotype H120 vaccine against GI-19-like strains. Avian Pathol. 2011, 40, 43–54. [Google Scholar] [CrossRef]
- Mo, M.-L.; Hong, S.-M.; Kwon, H.-J.; Kim, I.-H.; Song, C.-S.; Kim, J.-H. Genetic Diversity of Spike, 3a, 3b and E Genes of Infectious Bronchitis Viruses and Emergence of New Recombinants in Korea. Viruses 2013, 5, 550–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.S.; Lee, Y.J. Molecular and epidemiological characteristics of infectious bronchitis virus isolated in Korea. Korean J. Perinat. Med. 1986, 2, 91–98. [Google Scholar]
- Li, W.; Junker, D.; Hock, L.; Ebiary, E.; Collisson, E.W. Evolutionary implications of genetic variations in the S1 gene of infectious bronchitis virus. Virus Res. 1994, 34, 327–338. [Google Scholar] [CrossRef]
- Chen, H.W.; Huang, Y.P.; Wang, C.H. Identification of Taiwan and China-like recombinant avian infectioius bronchiis viruses in Taiwan. Virus Res. 2009, 140, 121–129. [Google Scholar] [CrossRef]
- Kuo, S.M.; Wang, C.H.; Hou, M.H.; Huang, Y.P.; Kao, H.W.; Su, H.L. Evolution of infectious bronchitis virus in Taiwan: Characterization of RNA recombination in the nucleocapsid gene. Vet. Microbiol. 2010, 144, 292–302. [Google Scholar] [CrossRef]
- Mase, M.; Inoue, T.; Yamaguchi, S.; Imada, T. Genetic Diversity of Avian Infectious Bronchitis Viruses in Japan Based on Analysis of S2 Glycoprotein Gene. J. Veter. Med. Sci. 2009, 71, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farsang, A.; Ros, C.; Renström, L.H.M.; Baule, C.; Soós, T.; Belák, S. Molecular epizootiology of infectious bronchitis virus in Sweden indicating the involvement of a vaccine strain. Avian Pathol. 2002, 31, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, L.; Godeke, G.-J.; Raamsman, M.J.B.; Masters, P.S.; Rottier, P.J.M. Retargeting of Coronavirus by Substitution of the Spike Glycoprotein Ectodomain: Crossing the Host Cell Species Barrier. J. Virol. 2000, 74, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- De Wit, J.J.S.; Cook, J.K.A.; Van Der Heijden, H.M.J.F. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Cheng, J.; Huo, C.; Zhao, J.; Liu, T.; Li, X.; Yan, S.; Wang, Z.; Hu, Y.; Zhang, G. Pathogenicity differences between GI-19-like and GI-1-type infectious bronchitis viruses. Vet. Microbiol. 2018, 213, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Denney, L.; Ho, L.-P. The role of respiratory epithelium in host defence against influenza virus infection. Biomed. J. 2018, 41, 218–233. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-Y.; Chen, H.-W. Infectious Bronchitis Virus Variants: Molecular Analysis and Pathogenicity Investigation. Int. J. Mol. Sci. 2017, 18, 2030. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IBV Strain | Year of Isolation | Type of Chicken | Location | Accession Number |
|---|---|---|---|---|
| Kr/D342/03 | 2003 | Broiler | Chungcheong | MW984619 |
| Kr/D343/03 | 2003 | Breeding | Chungcheong | MW984620 |
| Kr/D379/03 | 2003 | Broiler | Chungcheong | MW984621 |
| Kr/Q042/05 | 2005 | Broiler | Jeolla | MW984622 |
| Kr/D075/05 | 2005 | Broiler | Chungcheong | MW984623 |
| Kr/D079/05 | 2005 | Native | Geyonggi | MW984624 |
| Kr/D062/06 | 2006 | Broiler | Gangwon | MW984625 |
| Kr/Q043/06 | 2006 | Broiler | Geyonggi | MW984626 |
| Kr/Q118/08 | 2008 | Broiler | Chungcheong | MW984627 |
| Kr/D027/12 | 2012 | Broiler | Chungcheong | MW984628 |
| Kr/D021/13 | 2013 | Broiler | Jeolla | MW984629 |
| Kr/D022/13 | 2013 | Broiler | Jeolla | MW984630 |
| Kr/D024/13 | 2013 | Broiler | Jeolla | MW984631 |
| Kr/D025/13 | 2013 | Broiler | Jeolla | MW984632 |
| Kr/Q022/13 | 2013 | Broiler | Jeolla | MW984633 |
| Kr/Q028/13 | 2013 | Broiler | Jeolla | MW984634 |
| Kr/Q030/13 | 2013 | Broiler | Jeolla | MW984635 |
| Kr/D068/14 | 2014 | Native | Chungcheong | MW984636 |
| Kr/D069/14 | 2014 | Native | Chungcheong | MW984637 |
| Kr/D062/16 | 2016 | Native | Geyonggi | MW984638 |
| Kr/D053/17 | 2017 | Broiler | Gyeonsang | MW984639 |
| Kr/R024/17 | 2017 | Native | Chungcheong | MW984640 |
| Kr/Q037/18 | 2018 | Native | Gyeonsang | MW984641 |
| Kr/D013/18 | 2018 | Broiler | Jeolla | MW984642 |
| Kr/D014/18 | 2018 | Broiler | Geyonggi | MW984643 |
| Kr/R030/18 | 2018 | Broiler | Chungcheong | MW984644 |
| Kr/R38A/18 | 2018 | Native | Geyonggi | MW984645 |
| Kr/D024/18 | 2018 | Layer | Geyonggi | MW984646 |
| Kr/D025/18 | 2018 | Broiler | Jeolla | MW984647 |
| Kr/Q041/18 | 2018 | Broiler | Gangwon | MW984648 |
| Kr/D030/18 | 2018 | Broiler | Geyonggi | MW984649 |
| Kr/D038/18 | 2018 | Layer | Chungcheong | MW984650 |
| Kr/R038/18 | 2018 | Native | Geyonggi | MW984651 |
| Kr/R109/18 | 2018 | Native | Gyeonsang | MW984652 |
| Kr/Q054/18 | 2018 | Broiler | Jeolla | MW984653 |
| Kr/Q057/18 | 2018 | Broiler | Jeolla | MW984654 |
| Kr/D048/18 | 2018 | Broiler | Chungcheong | MW984655 |
| Kr/D063/18 | 2018 | Broiler | Geyonggi | MW984656 |
| Kr/R084/18 | 2018 | Broiler | Chungcheong | MW984657 |
| Kr/Q063/18 | 2019 | Broiler | Chungcheong | MW984658 |
| Kr/D005/19 | 2019 | Layer | Gyeonsang | MW984659 |
| Kr/Q002/19 | 2019 | Broiler | Chungcheong | MW984660 |
| Kr/Q008/19 | 2019 | Broiler | Gyeonsang | MW984661 |
| Kr/D021/19 | 2019 | Broiler | Jeolla | MW984662 |
| Kr/R019/19 | 2019 | Broiler | Gyeonsang | MW984663 |
| Kr/R024/19 | 2019 | Broiler | Gyeonsang | MW984664 |
| Kr/R014/19 | 2019 | Broiler | Gyeonsang | MW984665 |
| Kr/Q010/19 | 2019 | Broiler | Gyeonsang | MW984666 |
| Kr/D030/19 | 2019 | Broiler | Gyeonsang | MW984667 |
| Kr/D031/19 | 2019 | Layer | Geyonggi | MW984668 |
| Kr/D034/19 | 2019 | Broiler | Jeolla | MW984669 |
| Kr/D038/19 | 2019 | Broiler | Jeolla | MW984670 |
| Kr/D039/19 | 2019 | Broiler | Geyonggi | MW984671 |
| Kr/D060/19 | 2019 | Native | Chungcheong | MW984672 |
| Kr/D068/19 | 2019 | Layer | Geyonggi | MW984673 |
| Kr/D072/19 | 2019 | Broiler | Chungcheong | MW984674 |
| Isolate | Recombinant Pattern | Ciliostasis Score A | |||
|---|---|---|---|---|---|
| 3 dpi | 7 dpi | 14 dpi | 21 dpi | ||
| Kr/D030/18 | rGI-19-II | 0.0 ± 0.0 B | 4.0 ± 0.0 | 2.3 ± 0.6 | 0.9 ± 0.6 |
| Kr/Q037/18 | rGI-19-III | 0.0 ± 0.1 | 2.1 ± 0.9 | 3.9 ± 0.2 | 2.6 ± 0.9 |
| Kr/D025/18 | GI-19 | 0.0 ± 0.0 | 3.7 ± 0.5 | 4.0 ± 0.0 | 0.2 ± 0.5 |
| Control | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youn, S.-Y.; Lee, J.-Y.; Bae, Y.-C.; Kwon, Y.-K.; Kim, H.-R. Genetic and Pathogenic Characterization of QX(GI-19)-Recombinant Infectious Bronchitis Viruses in South Korea. Viruses 2021, 13, 1163. https://doi.org/10.3390/v13061163
Youn S-Y, Lee J-Y, Bae Y-C, Kwon Y-K, Kim H-R. Genetic and Pathogenic Characterization of QX(GI-19)-Recombinant Infectious Bronchitis Viruses in South Korea. Viruses. 2021; 13(6):1163. https://doi.org/10.3390/v13061163
Chicago/Turabian StyleYoun, So-Youn, Ji-Youn Lee, You-Chan Bae, Yong-Kuk Kwon, and Hye-Ryoung Kim. 2021. "Genetic and Pathogenic Characterization of QX(GI-19)-Recombinant Infectious Bronchitis Viruses in South Korea" Viruses 13, no. 6: 1163. https://doi.org/10.3390/v13061163
APA StyleYoun, S. -Y., Lee, J. -Y., Bae, Y. -C., Kwon, Y. -K., & Kim, H. -R. (2021). Genetic and Pathogenic Characterization of QX(GI-19)-Recombinant Infectious Bronchitis Viruses in South Korea. Viruses, 13(6), 1163. https://doi.org/10.3390/v13061163