

Heparin Inhibits SARS-CoV-2 Replication in Human Nasal Epithelial Cells

 , ,
, ,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Modeling
2.2. Virus and Cells
2.3. Human Nasal Airway Epithelial Cell Culture at Air-Liquid Interface (ALI)
2.4. SARS-CoV-2 Infection
2.5. Heparin-Treatment
2.6. Virus Titration
2.7. Viral RT-PCR
2.8. GFP Virus Detection
2.9. Immunofluorescence
2.10. Statistical Analysis
3. Results
3.1. Modeling of SARS-CoV-2 Spike with Heparin
3.2. Inhibition of SARS-CoV-2 Replication by Heparin in Vero Cells
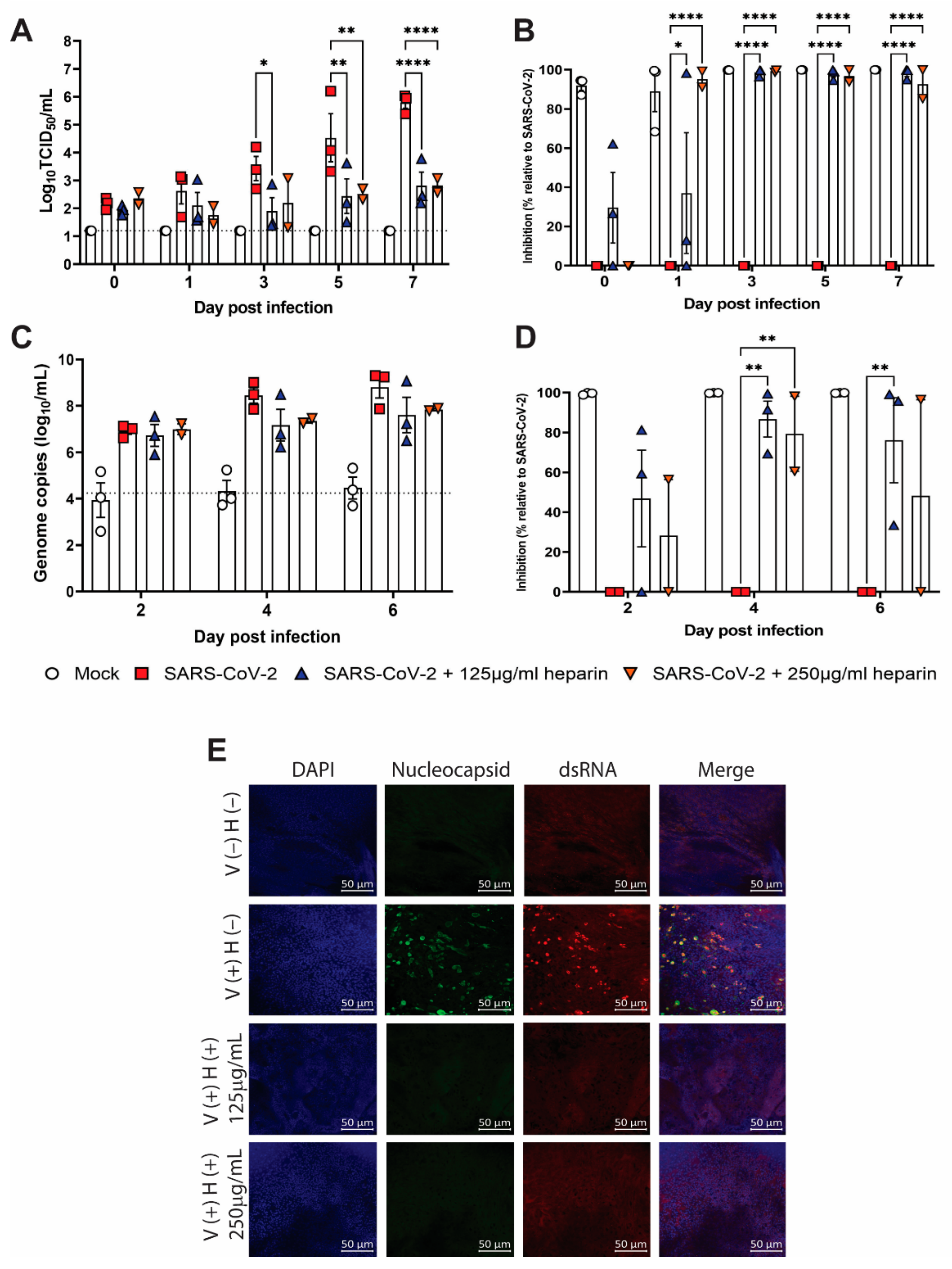
3.3. Inhibition of SARS-CoV-2 Replication by Heparin in Human Nasal Epithelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: A literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Joshi, A.D.; Lo, C.-H.; Drew, D.A.; Nguyen, L.H.; Guo, C.-G.; Ma, W.; Mehta, R.S.; Shebl, F.M.; Warner, E.T.; et al. Association of social distancing and face mask use with risk of COVID-19. Nat. Commun. 2021, 12, 3737. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 1, 2892. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Flight, K.E.; Higham, S.L.; Wang, Z.; Pierce, B.F. Progress of the COVID-19 vaccine effort: Viruses, vaccines and variants versus efficacy, effectiveness and escape. Nat. Rev. Immunol. 2021, 21, 626–636. [Google Scholar] [CrossRef]
- Sheikh-Mohamed, S.; Isho, B.; Chao, C.; Zuo, M.; Cohen, C.; Lustig, L.; Nahass, R.; Salomon, L.E.; Blacker, G.; Zarandi, M.; et al. Systemic and mucosal IgA responses are variably induced in response to SARS-CoV-2 mRNA vaccination and are associated with protection against subsequent infection. Mucosal. Immunol. 2022, 15, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of COVID-19—Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Jayk Bernal, A.; Gomes da Silva, M.M.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martín-Quirós, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for Oral Treatment of COVID-19 in Nonhospitalized Patients. N. Engl. J. Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Gunst, J.D.; Staerke, N.B.; Pahus, M.H.; Kristensen, L.H.; Bodilsen, J.; Lohse, N.; Dalgaard, L.S.; Brønnum, D.; Fröbert, O.; Hønge, B.; et al. Efficacy of the TMPRSS2 inhibitor camostat mesilate in patients hospitalized with COVID-19-a double-blind randomized controlled trial. e-Clin. Med. 2021, 35, 100849. [Google Scholar] [CrossRef] [PubMed]
- Boulware, D.R.; Pullen, M.F.; Bangdiwala, A.S.; Pastick, K.A.; Lofgren, S.M.; Okafor, E.C.; Skipper, C.P.; Nascene, A.A.; Nicol, M.R.; Abassi, M.; et al. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for COVID-19. N. Engl. J. Med. 2020, 383, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Ustianowski, A.; De Wit, S.; Launay, O.; Avila, M.; Templeton, A.; Yuan, Y.; Seegobin, S.; Ellery, A.; Levinson, D.J.; et al. Intramuscular AZD7442 (Tixagevimab-Cilgavimab) for Prevention of COVID-19. N. Engl. J. Med. 2022, 386, 2188–2200. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.M.; Sandoval, D.; Spliid, C.; Pihl, J.; Perett, H.; Painter, D.; Nayaranan, A.; Majowicz, S.; Kwong, S.; McVicar, R.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057.e15. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, C.; Swaroop, M.; Xu, M.; Wang, L.; Lee, J.; Wang, A.; Pradhan, M.; Hagen, N.; Chen, L.; et al. Heparan sulfate assists SARS-CoV-2 in cell entry and can be targeted by approved drugs in vitro. Cell Discov. 2020, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Jambrina, M.; Eder, J.; Kapten, T.; Hamme, J.; Helgers, L.; Vlaming, K.; Brower, P.; Nuenen, N.; Spaargaren, M.; de Bree, M.; et al. Infection and transmission of SARS-CoV-2 depend on heparan sulfate proteoglycans. EMBO J. 2021, 40, e106765. [Google Scholar] [CrossRef]
- Oduah, E.I.; Linhardt, R.J.; Sharfstein, S.T. Heparin: Past, Present, and Future. Pharmaceuticals 2016, 9, 2891. [Google Scholar] [CrossRef] [Green Version]
- Tandon, R.; Sharp, J.; Zhang, F.; Pomin, V.; Ashpole, N.; Mitra, D.; McCandless, M.; Jin, W.; Liu, H.; Sharma, P.; et al. Effective Inhibition of SARS-CoV-2 Entry by Heparin and Enoxaparin Derivatives. J. Virol. 2021, 95, 267. [Google Scholar] [CrossRef] [PubMed]
- Conzelmann, C.; Müller, J.A.; Perkhofer, L.; Sparrer, K.M.J.; Zelikin, A.N.; Münch, J.; Kleger, A. Inhaled and systemic heparin as a repurposed direct antiviral drug for prevention and treatment of COVID-19. Clin. Med. 2020, 20, e218–e221. [Google Scholar] [CrossRef]
- Paiardi, G.; Boyet, K.L.; Liu, H.; Dwivedi, R.; Ashpole, N.M.; Tandon, R.; Bidwell, G.L.; Cheng, Z.; Fassero, L.A.; Yu, C.S.; et al. The binding of heparin to spike glycoprotein inhibits SARS-CoV-2 infection by three mechanisms. J. Biol. Chem. 2022, 298, 101507. [Google Scholar] [CrossRef] [PubMed]
- van Haren, F.M.P.; Page, C.; Laffey, J.; Artigas, A.; Rimblas, M.; Nunes, Q.; Smith, R.; Shute, J.; Carroll, S.; Tree, J.; et al. Nebulised heparin as a treatment for COVID-19: Scientific rationale and a call for randomised evidence. Crit. Care 2020, 24, 454. [Google Scholar] [CrossRef] [PubMed]
- Harris, H.M.; Boyet, K.; Liu, H.; Dwivedi, R.; Ashpole, S.; Tandon, R.; Bidwell, G.; Cheng, Z.; Faserro, J.; Yu, R.; et al. Safety and Pharmacokinetics of Intranasally Administered Heparin. Pharm. Res. 2022, 39, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, A.G.; Benton, J.; Xu, P.; Roustan, C.; Martin, S.; Rosenthal, P.; Skehel, S.; Gamblin, S. SARS-CoV-2 and bat RaTG13 spike glycoprotein structures inform on virus evolution and furin-cleavage effects. Nat. Struct. Mol. Biol. 2020, 27, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- da Silva, F.L.B.; Giron, C.C.; Laaksonen, A. Electrostatic Features for the Receptor Binding Domain of SARS-CoV-2 Wildtype and Its Variants. Compass to the Severity of the Future Variants with the Charge-Rule. J. Phys. Chem. B 2022, 126, 6835–6852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; Wang, J. Surface charge changes in spike RBD mutations of SARS-CoV-2 and its variant strains alter the virus evasiveness via HSPGs: A review and mechanistic hypothesis. Front. Public Health 2022, 10, 952916. [Google Scholar] [CrossRef]
- Wang, J.T.; Wang, T.H.; Sheu, J.C.; Lin, S.M.; Lin, J.T.; Chen, D.S. Effects of anticoagulants and storage of blood samples on efficacy of the polymerase chain reaction assay for hepatitis C virus. J. Clin. Microbiol. 1992, 30, 750–753. [Google Scholar] [CrossRef] [Green Version]
- Yokota, M.; Tatsumi, N.; Nathalang, O.; Yamada, T.; Tsuda, I. Effects of heparin on polymerase chain reaction for blood white cells. J. Clin. Lab. Anal. 1999, 13, 133–140. [Google Scholar] [CrossRef]
- Subramaniyan, B.; Larabee, J.; Bodas, M.; Moore, A.; Burgett, A.; Myers, D.; Georgescu, C.; Wren, J.; Pafin, F.; Walters, M. Characterization of the SARS-CoV-2 Host Response in Primary Human Airway Epithelial Cells from Aged Individuals. Viruses 2021, 13, 1603. [Google Scholar] [CrossRef]
- Inde, Z.; Crocker, B.; Yapp, C.; Joshi, D.; Spetz, J.; Fraser, C.; Qin, F.; Xu, L.; Deskin, B.; Ghelfi, S.; et al. Age-dependent regulation of SARS-CoV-2 cell entry genes and cell death programs correlates with COVID-19 severity. Sci. Adv. 2021, 7, 2982. [Google Scholar] [CrossRef]
- Zhu, Y.; Chew, K.Y.; Wu, M.; Karawita, A.C.; McCallum, G.; Steele, L.E.; Yamamoto, A.; Labzin, L.I.; Yarlagadda, T.; Khromykh, A.A.; et al. Ancestral SARS-CoV-2, but not Omicron, replicates less efficiently in primary pediatric nasal epithelial cells. PLoS Biol. 2022, 20, e3001728. [Google Scholar] [CrossRef]
- Liu, L.; Chopra, P.; Li, X.; Bouwman, S.; Tompkins, S.; Wolfert, S.; de Vries, R.; Boons, G. Heparan Sulfate Proteoglycans as Attachment Factor for SARS-CoV-2. ACS Cent. Sci. 2021, 7, 1009–1018. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, L.Y.Y.; Suryadinata, R.; McCafferty, C.; Ignjatovic, V.; Purcell, D.F.J.; Robinson, P.; Morton, C.J.; Parker, M.W.; Anderson, G.P.; Monagle, P.; et al. Heparin Inhibits SARS-CoV-2 Replication in Human Nasal Epithelial Cells. Viruses 2022, 14, 2620. https://doi.org/10.3390/v14122620
Lee LYY, Suryadinata R, McCafferty C, Ignjatovic V, Purcell DFJ, Robinson P, Morton CJ, Parker MW, Anderson GP, Monagle P, et al. Heparin Inhibits SARS-CoV-2 Replication in Human Nasal Epithelial Cells. Viruses. 2022; 14(12):2620. https://doi.org/10.3390/v14122620
Chicago/Turabian StyleLee, Leo Yi Yang, Randy Suryadinata, Conor McCafferty, Vera Ignjatovic, Damian F. J. Purcell, Phil Robinson, Craig J. Morton, Michael W. Parker, Gary P. Anderson, Paul Monagle, and et al. 2022. "Heparin Inhibits SARS-CoV-2 Replication in Human Nasal Epithelial Cells" Viruses 14, no. 12: 2620. https://doi.org/10.3390/v14122620
APA StyleLee, L. Y. Y., Suryadinata, R., McCafferty, C., Ignjatovic, V., Purcell, D. F. J., Robinson, P., Morton, C. J., Parker, M. W., Anderson, G. P., Monagle, P., Subbarao, K., & Neil, J. A. (2022). Heparin Inhibits SARS-CoV-2 Replication in Human Nasal Epithelial Cells. Viruses, 14(12), 2620. https://doi.org/10.3390/v14122620