
Modeling the Impact of Vaccination on COVID-19 and Its Delta and Omicron Variants

 , , , and
, , , and
Abstract
:1. Introduction
- whether herd immunity can be achieved and the pandemic eventually eradicated;
- the risk of COVID-19 epidemic resurgence [4];
- what are the driving socio-demographic and health factors behind the epidemic progression;
- how to optimize vaccination strategy [5], or whether long-term co-existence with the virus should be pursued instead;
- whether and when to reopen international or national (e.g., state or provincial) borders;
- whether additional surge capacity is required at hospitals; and
- who to prioritize during vaccination (e.g., providing second doses or booster shots versus increasing efforts to give more people their first dose).
1.1. Background and Related Work
1.2. Contributions of This Paper
- We propose a mathematical model to characterize the epidemiological process of COVID-19 with vaccination and derive formulas for its vaccine reproduction number (similar to but assuming a fully vaccinated population) and effective reproduction number. We show that, based on the current ratio of CoronaVac (Sinovac) and Comirnaty (Pfizer–BioNTech) vaccinations in Hong Kong, herd immunity could not have been obtained via vaccination alone. Furthermore, no ratio of CoronaVac versus Comirnaty vaccinations can achieve herd immunity against Omicron via vaccination alone.
- Using our new model, we observe the impact of vaccination in terms of various parameters (including the initial vaccination coverage at the start of a new outbreak, the rate of new vaccinations, the average vaccine efficacy, and NPI intensity) on the cumulative number of hospitalized cases, for both the original and Delta strains of the SARS-CoV-2 virus, as well as the Omicron variant.
- We compute, for different vaccination rates and vaccine efficacies, the minimum initial vaccination coverage required such that the cumulative number of hospitalized/isolated cases is less than a given percentage of the population. Furthermore, we demonstrate how a high vaccination rate decreases the required initial vaccination coverage.
- We find that the traditional formula (1) for herd immunity, assuming a static vaccination coverage (i.e., an initial vaccinated group with no additional vaccinations), corresponds to a cumulative hospitalization ratio of about one percent.
- We show how regions can achieve herd immunity at a lower vaccination coverage ratio than that predicted by (1), but at the cost of additional infections (natural immunity versus induced immunity).
2. Methods
2.1. Compartmental Modeling for COVID-19
2.2. Reproduction Numbers
- The basic reproduction number , referred to above, is the average number of individuals an infectious individual will infect in a wholly susceptible population, without vaccination.
- The vaccine reproduction number is the average number of individuals an infectious individual will infect in a wholly vaccinated population.
- The effective reproduction number is the average number of individuals an infectious individual will infect based on the current situation at time t.
2.3. Herd Immunity and Multiple Vaccines
- If : either vaccine can achieve herd immunity via vaccination alone.
- If : neither vaccine can achieve herd immunity via vaccination alone.
- Otherwise: whether herd immunity can be achieved via vaccination alone depends on the mix of vaccines administered.
2.4. Asymptotic Behavior
3. Results and Discussions
3.1. Ancestral Strain
3.2. The Second and Third Waves in Hong Kong
3.3. Sensitivity with Respect to Initial Vaccination Coverage and Vaccination Rate k
3.4. Sensitivity with Respect to Vaccine Efficacy and Initial Vaccination Coverage
3.5. Measuring the Effective Reproduction Number over Time
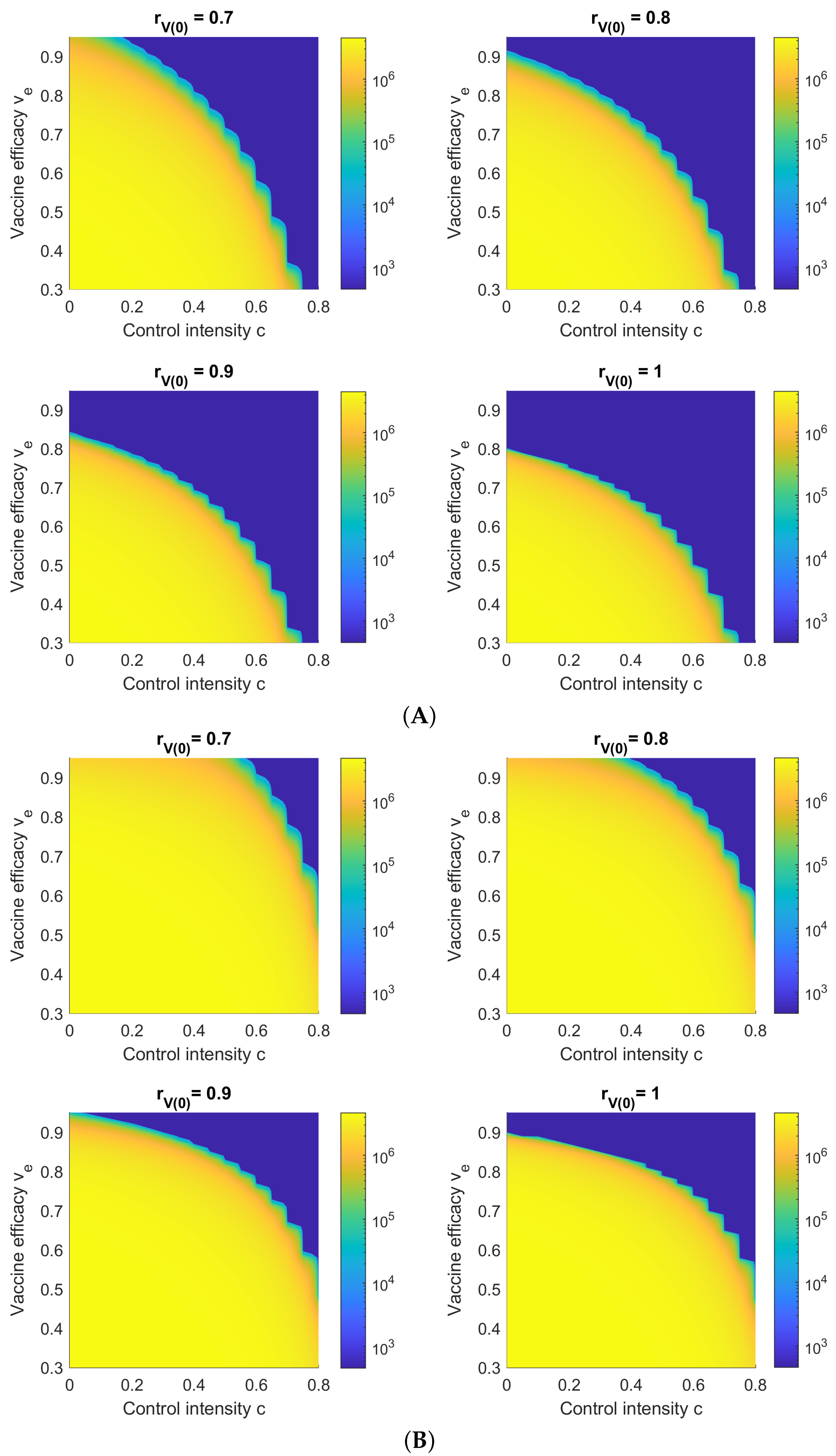
3.6. Sensitivity with Respect to Vaccine Efficacy and NPI Control Intensity c
3.7. Sensitivity with Respect to Vaccine Efficacy and Reproduction Number
3.8. Dynamic Versus Static Vaccination Thresholds
- denotes the minimum initial vaccination coverage for a given vaccination rate k such that .
- denotes the dynamic vaccination threshold, which we define as , where is the initial vaccination coverage and denotes the minimum time after which even if all new vaccinations were to cease at time .
4. Conclusions
- For the ancestral strain of SARS-CoV-2 () and a 6:4 ratio of Comirnaty and CoronaVac vaccination, as currently observed in Hong Kong, the theoretical vaccination threshold for herd immunity is , implying that herd immunity is possible via vaccination alone.
- For the Delta variant (), herd immunity is possible via vaccination alone using Comirnaty, but not using the current ratio of Comirnaty and CoronaVac in Hong Kong, even if the entire population is to be vaccinated—the proportion of Comirnaty vaccinations has to increase.
- For the Omicron variant (), herd immunity is impossible via vaccination alone, regardless of vaccine mix and even if the entire population is to be vaccinated. Similar results can be obtained for newer variants such as BA.4 or BA.5 by adjusting the model’s parameters.
- NPI control measures are required to limit an outbreak until vaccination has reached a sufficiently high level. Moreover, if the basic reproduction number is high and the vaccine efficacy relatively low, then control measures are required to prevent a mass outbreak even when vaccine coverage is high (in some cases, even if the population is fully vaccinated).
- Faster vaccination results in a lower initial vaccine coverage ratio required to maintain the size of an outbreak under a given limit. Thus governments can consider lifting controls even before the theoretical vaccination threshold for herd immunity is reached.
- Increasing the number of individuals who are allowed to become infected reduces the requirement on the number of individuals that need to be vaccinated before herd immunity is reached. However, over-reliance on infection-acquired natural immunity may lead to a large number of hospitalizations and a decline in the quality of medical care due to capacity exceedance, increasing the case fatality ratio.
Limitations of This Work
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| The following abbreviations are used in this manuscript: | |
| COVID-19 | Coronavirus disease 2019 |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| SVEIHR | susceptible-vaccinated-exposed-infected-hospitalized-removed |
| NPI | non-pharmaceutical intervention |
References
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans: Dynamics and Control; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Brauer, F.; Castillo-Chavez, C.; Feng, Z. Mathematical Models in Epidemiology; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Holmdahl, I.; Buckee, C. Wrong but useful—what COVID-19 epidemiologic models can and cannot tell us. New Engl. J. Med. 2020, 383, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Krueger, T.; Gogolewski, K.; Bodych, M.; Gambin, A.; Giordano, G.; Cuschieri, S.; Czypionka, T.; Perc, M.; Petelos, E.; Rosińska, M.; et al. Risk assessment of COVID-19 epidemic resurgence in relation to SARS-CoV-2 variants and vaccination passes. Commun. Med. 2022, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Markovič, R.; Šterk, M.; Marhl, M.; Perc, M.; Gosak, M. Socio-demographic and health factors drive the epidemic progression and should guide vaccination strategies for best COVID-19 containment. Results Phys. 2021, 26, 104433. [Google Scholar] [CrossRef] [PubMed]
- Matrajt, L.; Eaton, J.; Leung, T.; Dimitrov, D.; Schiffer, J.T.; Swan, D.A.; Janes, H. Optimizing vaccine allocation for COVID-19 vaccines shows the potential role of single-dose vaccination. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bubar, K.M.; Reinholt, K.; Kissler, S.M.; Lipsitch, M.; Cobey, S.; Grad, Y.H.; Larremore, D.B. Model-informed COVID-19 vaccine prioritization strategies by age and serostatus. Science 2021, 371, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Cai, J.; Yang, J.; Zhang, J.; Wu, Q.; Zheng, W.; Shi, H.; Ajelli, M.; Zhou, X.H.; Yu, H. Time-varying optimization of COVID-19 vaccine prioritization in the context of limited vaccination capacity. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Hill, E.M.; Dyson, L.; Tildesley, M.J.; Keeling, M.J. Modelling optimal vaccination strategy for SARS-CoV-2 in the UK. PLoS Comput. Biol. 2021, 17, e1008849. [Google Scholar] [CrossRef]
- Piraveenan, M.; Sawleshwarkar, S.; Walsh, M.; snd Samit Bhattacharyya, I.Z.; Farooqui, H.H.; Zomaya, A.; Perc, M. Optimal governance and implementation of vaccination programmes to contain the COVID-19 pandemic. R. Soc. Open Sci. 2002, 8, 210429. [Google Scholar] [CrossRef]
- Liu, Y.; Rocklöv, J. The reproductive number of the Delta variant of SARS-CoV-2 is far higher compared to the ancestral SARS-CoV-2 virus. J. Travel Med. 2021, 28, 124. [Google Scholar] [CrossRef]
- Liu, Y.; Rocklöv, J. The effective reproduction number for the omicron SARS-CoV-2 variant of concern is several times higher than Delta. J. Travel Med. 2022, 29, taac037. [Google Scholar] [CrossRef]
- Giordano, G.; Colaneri, M.; Di Filippo, A.; Blanchini, F.; Bolzern, P.; De Nicolao, G.; Sacchi, P.; Colaneri, P.; Bruno, R. Modeling vaccination rollouts, SARS-CoV-2 variants and the requirement for non-pharmaceutical interventions in Italy. Nat. Med. 2021, 27, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Grundel, S.M.; Heyder, S.; Hotz, T.; Ritschel, T.K.; Sauerteig, P.; Worthmann, K. How to coordinate vaccination and social distancing to mitigate SARS-CoV-2 outbreaks. SIAM J. Appl. Dyn. Syst. 2021, 20, 1135–1157. [Google Scholar] [CrossRef]
- Niu, R.; Chan, Y.C.; Wong, E.W.; van Wyk, M.A.; Chen, G. A stochastic SEIHR model for COVID-19 data fluctuations. Nonlinear Dyn. 2021, 106, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Niu, R.; Wong, E.W.; Chan, Y.C.; Van Wyk, M.A.; Chen, G. Modeling the COVID-19 pandemic using an SEIHR model with human migration. IEEE Access 2020, 8, 195503–195514. [Google Scholar] [CrossRef]
- Government of the Hong Kong Special Adminstrative Region. Hong Kong Vaccination Dashboard. Available online: https://www.covidvaccine.gov.hk/en/dashboard (accessed on 10 April 2022).
- Lopez Bernal, J.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of COVID-19 vaccines against the B.1.617.2 (Delta) variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef]
- Palacios, R.; Batista, A.P.; Albuquerque, C.S.N.; Patiño, E.G.; Santos, J.d.P.; Tilli Reis Pessoa Conde, M.; Piorelli, R.d.O.; Pereira Júnior, L.C.; Raboni, S.M.; Ramos, F.; et al. Efficacy and Safety of a COVID-19 Inactivated Vaccine in Healthcare Professionals in Brazil: The PROFISCOV Study. SSRN 2021. [Google Scholar] [CrossRef]
- Pormohammad, A.; Zarei, M.; Ghorbani, S.; Mohammadi, M.; Aghayari Sheikh Neshin, S.; Khatami, A.; Turner, D.L.; Djalalinia, S.; Mousavi, S.A.; Mardani-Fard, H.A.; et al. Effectiveness of COVID-19 Vaccines against Delta (B.1.617.2) Variant: A Systematic Review and Meta-Analysis of Clinical Studies. Vaccines 2021, 10, 23. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.M.; et al. COVID-19 vaccine effectiveness against the omicron (B.1.1.529) variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Pérez-Then, E.; Lucas, C.; Monteiro, V.S.; Miric, M.; Brache, V.; Cochon, L.; Vogels, C.B.F.; Malik, A.A.; de la Cruz, E.; Jorge, A.; et al. Neutralizing antibodies against the SARS-CoV-2 Delta and Omicron variants following heterologous CoronaVac plus BNT162b2 booster vaccination. R. Soc. Open Sci. 2021, 8, 210429. [Google Scholar] [CrossRef]
- Government of the Hong Kong Special Adminstrative Region. Hong Kong Population Data. Available online: https://www.news.gov.hk/eng/2021/08/20210812/20210812_163730_419.html (accessed on 10 April 2022).
- Howard, J.; Huang, A.; Li, Z.; Tufekci, Z.; Zdimal, V.; van der Westhuizen, H.M.; von Delft, A.; Price, A.; Fridman, L.; Tang, L.H.; et al. An evidence review of face masks against COVID-19. Proc. Natl. Acad. Sci. USA 2021, 118, e2014564118. [Google Scholar] [CrossRef]
- Shrotri, M.; Navaratnam, A.M.; Nguyen, V.; Byrne, T.; Geismar, C.; Fragaszy, E.; Beale, S.; Fong, W.L.E.; Patel, P.; Kovar, J.; et al. Spike-antibody waning after second dose of BNT162b2 or ChAdOx1. Lancet 2021, 398, 385–387. [Google Scholar] [CrossRef]
- Britton, T.; Ball, F.; Trapman, P. A mathematical model reveals the influence of population heterogeneity on herd immunity to SARS-CoV-2. Science 2020, 369, 846–849. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition |
|---|---|
| N | Population size |
| Transmission rate of exposed individuals, unvaccinated targets | |
| Transmission rate of infected individuals, unvaccinated targets | |
| (Mean) vaccine efficacy | |
| Transmission rate of exposed individuals, vaccinated targets (refer to Definition 1) | |
| Transmission rate of infected individuals, vaccinated targets (refer to Definition 1) | |
| Rate of exposed individuals becoming symptomatic | |
| Hospitalization/isolation rate of infected individuals. Our model does not differentiate between hospitalization and non-clinical isolation of confirmed cases. | |
| Recovery rate of exposed, infected, or hospitalized/isolated individuals | |
| k | Vaccination rate of susceptible individuals |
| Ratio of vaccinated individuals at time 0 (i.e., , where N is the total population) | |
| c | Control intensity of non-pharmaceutical interventions (NPIs), where denotes no control, and denotes complete cessation of disease transmission |
| Initial Value | Second Wave | Third Wave |
|---|---|---|
| 9 | 9 | |
| 1 | 1 | |
| 0 | 0 | |
| 72 | 1193 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Chan, Y.-C.; Niu, R.; Wong, E.W.M.; van Wyk, M.A. Modeling the Impact of Vaccination on COVID-19 and Its Delta and Omicron Variants. Viruses 2022, 14, 1482. https://doi.org/10.3390/v14071482
Wang J, Chan Y-C, Niu R, Wong EWM, van Wyk MA. Modeling the Impact of Vaccination on COVID-19 and Its Delta and Omicron Variants. Viruses. 2022; 14(7):1482. https://doi.org/10.3390/v14071482
Chicago/Turabian StyleWang, Jianbo, Yin-Chi Chan, Ruiwu Niu, Eric W. M. Wong, and Michaël Antonie van Wyk. 2022. "Modeling the Impact of Vaccination on COVID-19 and Its Delta and Omicron Variants" Viruses 14, no. 7: 1482. https://doi.org/10.3390/v14071482
APA StyleWang, J., Chan, Y. -C., Niu, R., Wong, E. W. M., & van Wyk, M. A. (2022). Modeling the Impact of Vaccination on COVID-19 and Its Delta and Omicron Variants. Viruses, 14(7), 1482. https://doi.org/10.3390/v14071482