
Three Distinct Reporter Systems of Hepatitis E Virus and Their Utility as Drug Screening Platforms


Abstract
:1. Introduction
2. The HEV-GLuc Replicon: An Advanced Tool for Monitoring HEV Replication and Drug Screening

{kind=link}
{kind=link}
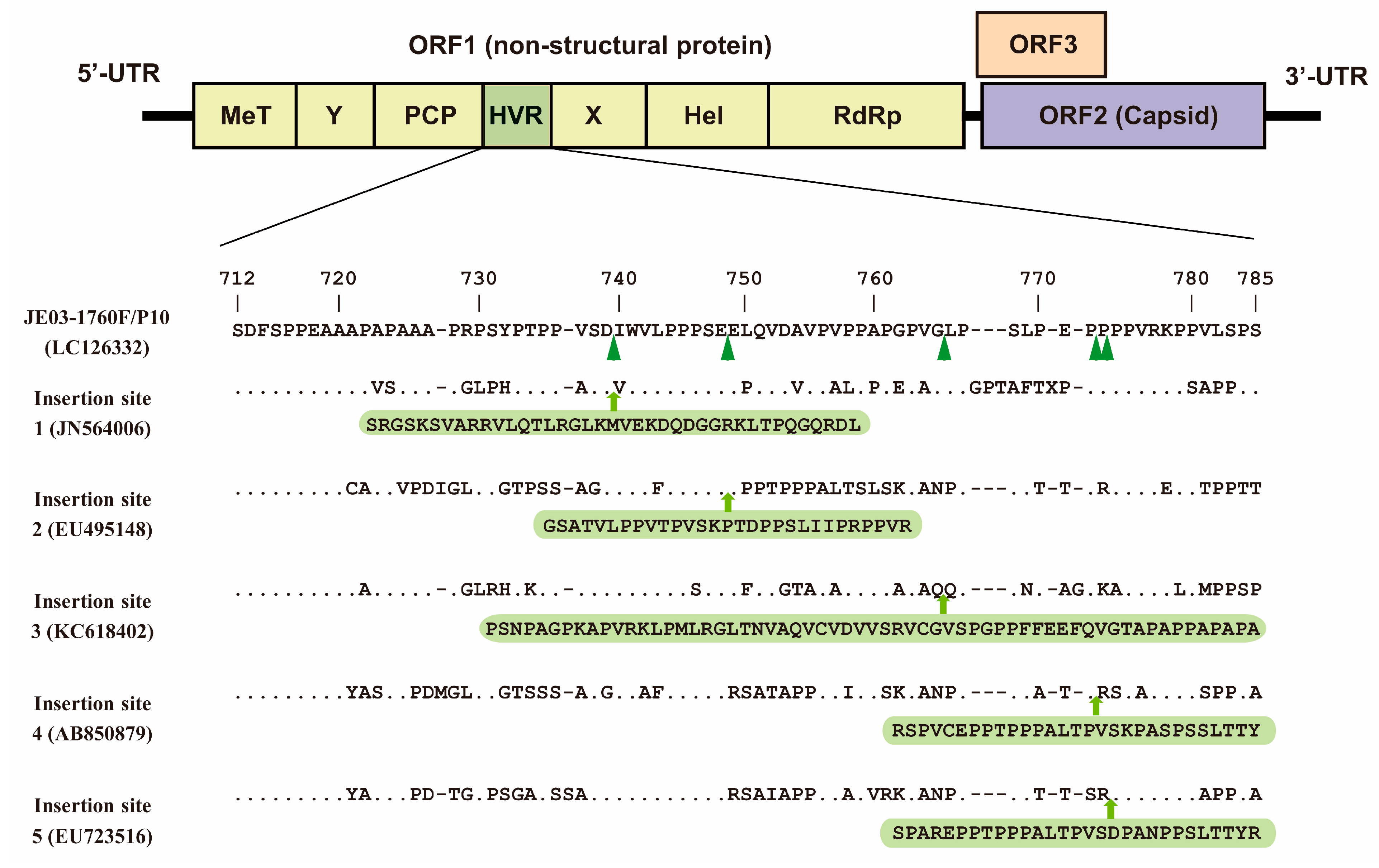{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype (Origin) | Application for: | Reference | |
|---|---|---|---|---|
| Drug Screening | Analysis of Life Cycle | |||
| Kernow-C1 p6 | HEV-3 (human) | NA | Role of insertion of human S17 ribosomal protein sequence in growth advantage | [71] |
| Deptropine | NA | [77] | ||
| Gemcitabine | NA | [78] | ||
| Isocotoin | NA | [79] | ||
| HEV83-2-27 | HEV-3 (swine) | NA | Importance of C-terminal 52 amino acids for HEV life cycle | [74] |
| Sar55/S17 | HEV-1 (human) | NA | Identification of possible determinants of host range | [67] |
| LA-B350 | HEV-C1 (rat) | NA | Establishment of subgenomic replicon for various HEV studies | [73] |
| Sar55 | HEV-1 (human) | NA | Identification of viral regulatory elements and intracellular genome dynamics | [72] |
| SHEV3 | HEV-3 (swine) | NA | Identification of viral regulatory elements and intracellular genome dynamics | [72] |
| TW6196-E | HEV-4 (human) | NA | Identification of viral regulatory elements and intracellular genome dynamics | [72] |
| JE03-1760F/P10 | HEV-3 (human) | Ciprofloxacine | NA | [60] |
| Kernow-C1 p6 (with HA- or V5-tagged ORF1) | HEV-3 (human) | NA | Identification of candidate HEV factories | [76] |
| HEV-3ra LR | HEV-3ra (rabbit) | NA | Investigation of the impact of ribavirin-treatment-failure-associated RdRp mutations of human HEV-3 on in vitro replication of HEV-3ra | [75] |
3. The Recombinant Infectious HEV-nanoKAZ
4. The Recombinant Infectious HEV-HiBiT
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purdy, M.A.; Drexler, J.F.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV virus taxonomy profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef]
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): Molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120–131. [Google Scholar] [CrossRef]
- Kabrane-Lazizi, Y.; Meng, X.J.; Purcell, R.H.; Emerson, S.U. Evidence that the genomic RNA of hepatitis E virus is capped. J. Virol. 1999, 73, 8848–8850. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Gorbalenya, A.E.; Purdy, M.A.; Rozanov, M.N.; Reyes, G.R.; Bradley, D.W. Computer-assisted assignment of functional domains in the nonstructural polyprotein of hepatitis E virus: Delineation of an additional group of positive-strand RNA plant and animal viruses. Proc. Natl. Acad. Sci. USA 1992, 89, 8259–8263. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Varma, S.P. Hepatitis E: Molecular virology and pathogenesis. J. Clin. Exp. Hepatol. 2013, 3, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.C.; Saliou, J.M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F.; et al. Hepatitis E virus lifecycle and identification of 3 forms of the ORF2 capsid protein. Gastroenterology 2018, 154, 211–223. [Google Scholar] [CrossRef]
- Yin, X.; Ying, D.; Lhomme, S.; Tang, Z.; Walker, C.M.; Xia, N.; Zheng, Z.; Feng, Z. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc. Natl. Acad. Sci. USA 2018, 115, 4773–4778. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Takahashi, M.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Nagashima, S.; Tanaka, T.; Okamoto, H. ORF3 protein of hepatitis E virus is essential for virion release from infected cells. J. Gen. Virol. 2009, 90, 1880–1891. [Google Scholar] [CrossRef]
- Emerson, S.U.; Nguyen, H.T.; Torian, U.; Burke, D.; Engle, R.; Purcell, R.H. Release of genotype 1 hepatitis E virus from cultured hepatoma and polarized intestinal cells depends on open reading frame 3 protein and requires an intact PXXP motif. J. Virol. 2010, 84, 9059–9069. [Google Scholar] [CrossRef]
- Nagashima, S.; Takahashi, M.; Jirintai; Tanaka, T.; Yamada, K.; Nishizawa, T.; Okamoto, H. A PSAP motif in the ORF3 protein of hepatitis E virus is necessary for virion release from infected cells. J. Gen. Virol. 2011, 92, 269–278. [Google Scholar] [CrossRef]
- Ding, Q.; Heller, B.; Capuccino, J.M.; Song, B.; Nimgaonkar, I.; Hrebikova, G.; Contreras, J.E.; Ploss, A. Hepatitis E virus ORF3 is a functional ion channel required for release of infectious particles. Proc. Natl. Acad. Sci. USA 2017, 114, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the ORF3 protein of hepatitis E virus (HEV) can capture HEV particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Jirintai; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; et al. Hepatitis E virus (HEV) strains in serum samples can replicate efficiently in cultured cells despite the coexistence of HEV antibodies: Characterization of HEV virions in blood circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct entry mechanisms for nonenveloped and quasi-enveloped hepatitis E viruses. J. Virol. 2016, 90, 4232–4242. [Google Scholar] [CrossRef]
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Characterization of the quasi-enveloped hepatitis E virus particles released by the cellular exosomal pathway. J. Virol. 2017, 91, e00822-17. [Google Scholar] [CrossRef] [PubMed]
- Primadharsini, P.P.; Nagashima, S.; Okamoto, H. Mechanism of cross-species transmission, adaptive evolution and pathogenesis of hepatitis E virus. Viruses 2021, 13, 909. [Google Scholar] [CrossRef]
- Rasche, A.; Sander, A.L.; Corman, V.M.; Drexler, J.F. Evolutionary biology of human hepatitis viruses. J. Hepatol. 2019, 70, 501–520. [Google Scholar] [CrossRef]
- Primadharsini, P.P.; Nagashima, S.; Okamoto, H. Genetic variability and evolution of hepatitis E virus. Viruses 2019, 11, 456. [Google Scholar] [CrossRef]
- Izopet, J.; Dubois, M.; Bertagnoli, S.; Lhomme, S.; Marchandeau, S.; Boucher, S.; Kamar, N.; Abravanel, F.; Guerin, J.L. Hepatitis E virus strains in rabbits and evidence of a closely related strain in humans, France. Emerg. Infect. Dis. 2012, 18, 1274–1281. [Google Scholar] [CrossRef]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Takahashi, K.; Kitajima, N.; Abe, N.; Mishiro, S. Complete or near-complete nucleotide sequences of hepatitis E virus genome recovered from a wild boar, a deer, and four patients who ate the deer. Virology 2004, 330, 501–505. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, Y.; Miyazaki, S.; Aikawa, T.; Ashida, K.; Tamaru, T.; Oguro, K.; Hayakawa, F.; Matsuoka, H.; et al. Prevalence and genotype/subtype distribution of hepatitis E virus (HEV) among wild boars in Japan: Identification of a genotype 5 HEV strain. Virus Res. 2020, 287, 198106. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a hepatitis E virus isolate obtained from a wild boar in Japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar] [CrossRef]
- Rasche, A.; Saqib, M.; Liljander, A.M.; Bornstein, S.; Zohaib, A.; Renneker, S.; Steinhagen, K.; Wernery, R.; Younan, M.; Gluecks, I.; et al. Hepatitis E virus infection in dromedaries, North and East Africa, United Arab Emirates, and Pakistan, 1983–2015. Emerg. Infect. Dis. 2016, 22, 1249–1252. [Google Scholar] [CrossRef]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic infection with camelid hepatitis E virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology 2016, 150, 355–357e353. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New hepatitis E virus genotype in Bactrian camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Takahashi, M.; Tsatsralt-Od, B.; Nyamdavaa, K.; Dulmaa, N.; Osorjin, B.; Tseren-Ochir, E.O.; Sharav, T.; Bayasgalan, C.; Sukhbaatar, B.; et al. Identification and a full genome analysis of novel camel hepatitis E virus strains obtained from Bactrian camels in Mongolia. Virus Res. 2021, 299, 198355. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.Y.; Wu, S.; Cai, J.; Zhang, A.J.; Leung, K.H.; Chung, T.W.H.; Chan, J.F.W.; Chan, W.M.; Teng, J.L.L.; et al. Rat hepatitis E virus as cause of persistent hepatitis after liver transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W.; et al. Transmission of rat hepatitis E virus infection to humans in Hong Kong: A clinical and epidemiological analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat hepatitis E virus linked to severe acute hepatitis in an immunocompetent patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Lhomme, S.; Abravanel, F.; Cintas, P.; Izopet, J. Hepatitis E virus infection: Neurological manifestations and pathophysiology. Pathogens 2021, 10, 1582. [Google Scholar] [CrossRef]
- Guinault, D.; Ribes, D.; Delas, A.; Milongo, D.; Abravanel, F.; Puissant-Lubrano, B.; Izopet, J.; Kamar, N. Hepatitis E virus-induced cryoglobulinemic glomerulonephritis in a nonimmunocompromised person. Am. J. Kidney Dis. 2016, 67, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Del Bello, A.; Guilbeau-Frugier, C.; Josse, A.G.; Rostaing, L.; Izopet, J.; Kamar, N. Successful treatment of hepatitis E virus-associated cryoglobulinemic membranoproliferative glomerulonephritis with ribavirin. Transpl. Infect. Dis. 2015, 17, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Cheema, S.S.; Cheema, M.F.; Gilani, S.; Cheema, S.R. Immunoglobulin A nephropathy associated with acute hepatitis E infection: First case report. Clin. Nephrol. Case Stud. 2023, 11, 95–98. [Google Scholar] [CrossRef]
- Riveiro-Barciela, M.; Bes, M.; Quer, J.; Valcarcel, D.; Piriz, S.; Gregori, J.; Llorens, M.; Salcedo, M.T.; Piron, M.; Esteban, R.; et al. Thrombotic thrombocytopenic purpura relapse induced by acute hepatitis E transmitted by cryosupernatant plasma and successfully controlled with ribavirin. Transfusion 2018, 58, 2501–2505. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Xu, Y.; Tan, Z.; Pang, L.; Li, S.; Li, Q.; Cai, W.; Nan, Y.; Tu, J. A case of acquired thrombotic thrombocytopenic purpura induced by acute severe hepatitis E: Successfully treated by plasma exchange and rituximab. Thromb. J. 2023, 21, 74. [Google Scholar] [CrossRef]
- Lv, F.; Zhao, Y.; Yang, X.D.; Chen, H.Z.; Ren, W.Y.; Chen, L.X.; Yi, Q.Q.; Zheng, W.; Pan, H.Y. Acute hepatitis E induced the first episode of immune-mediated thrombotic thrombocytopenic turpura: The first case report. Infect. Drug Resist. 2023, 16, 5149–5154. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Lal, A.; Idrees, M.; Hussain, A.; Jeet, C.; Malik, F.A.; Iqbal, Z.; Rehman, H. Hepatitis E virus-associated aplastic anaemia: The first case of its kind. J. Clin. Virol. 2012, 54, 96–97. [Google Scholar] [CrossRef]
- Thapa, R.; Pramanik, S.; Biswas, B.; Mallick, D. Hepatitis E virus infection in a 7-year-old boy with glucose 6-phosphate dehydrogenase deficiency. J. Pediatr. Hematol. Oncol. 2009, 31, 223–224. [Google Scholar] [CrossRef]
- Raj, M.; Kumar, K.; Ghoshal, U.C.; Saraswat, V.A.; Aggarwal, R.; Mohindra, S. Acute hepatitis E-associated acute pancreatitis: A single center experience and literature review. Pancreas. 2015, 44, 1320–1322. [Google Scholar] [CrossRef]
- Wu, J.; Xiang, Z.; Gao, C.; Huang, L.; Hua, J.; Tong, L.; Ling, B.; Yao, Y.; Jiang, B.; Wang, D.; et al. Chinese Consortium for the Study of Hepatitis E. Genotype 4 HEV infection triggers the initiation and development of acute pancreatitis. Microbes Infect. 2023, online ahead of print, 105190. [Google Scholar] [CrossRef]
- Sengupta, P.; Biswas, S.; Roy, T. Hepatitis E-induced acute myocarditis in an elderly woman. Case Rep. Gastroenterol. 2019, 13, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Emeriaud, H.; Huet, F.; Roubille, F.; Pasquie, J.L. Acute myocarditis induced by hepatitis E: An uncommon association. CJC Open 2022, 4, 729–731. [Google Scholar] [CrossRef] [PubMed]
- Dumoulin, F.L.; Liese, H. Acute hepatitis E virus infection and autoimmune thyroiditis: Yet another trigger? BMJ Case Rep. 2012, 2012, bcr1220115441. [Google Scholar] [CrossRef]
- Martinez-Artola, Y.; Poncino, D.; Garcia, M.L.; Munne, M.S.; Gonzalez, J.; Garcia, D.S. Acute hepatitis E virus infection and association with a subacute thyroiditis. Ann. Hepatol. 2015, 14, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL clinical practice guidelines on hepatitis E virus infection. J. Hepatol. 2018, 68, 1256–1271. [Google Scholar] [CrossRef]
- Lampejo, T. Sofosbuvir in the treatment of hepatitis E virus infection: A review of in vitro and in vivo evidence. J. Clin. Exp. Hepatol. 2022, 12, 1225–1237. [Google Scholar] [CrossRef]
- Todt, D.; François, C.; Anggakusuma; Behrendt, P.; Engelmann, M.; Knegendorf, L.; Vieyres, G.; Wedemeyer, H.; Hartmann, R.; Pietschmann, T.; et al. Antiviral activities of different interferon types and subtypes against hepatitis E virus replication. Antimicrob. Agents. Chemother. 2016, 60, 2132–2139. [Google Scholar] [CrossRef]
- Fang, Y.; Chen, C.Y.; Yu, H.C.; Lin, P.C. Neuropsychiatric disorders in chronic hepatitis C patients after receiving interferon or direct-acting antivirals: A nationwide cohort study. Front. Pharmacol. 2023, 14, 1191843. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Rostaing, L.; Abravanel, F.; Garrouste, C.; Esposito, L.; Cardeau-Desangles, I.; Mansuy, J.M.; Selves, J.; Peron, J.M.; Otal, P.; et al. Pegylated interferon-alpha for treating chronic hepatitis E virus infection after liver transplantation. Clin. Infect. Dis. 2010, 50, e30–e33. [Google Scholar] [CrossRef] [PubMed]
- Haagsma, E.B.; Riezebos-Brilman, A.; van den Berg, A.P.; Porte, R.J.; Niesters, H.G. Treatment of chronic hepatitis E in liver transplant recipients with pegylated interferon alpha-2b. Liver Transpl. 2010, 16, 474–477. [Google Scholar] [CrossRef]
- Saab, S.; Kalmaz, D.; Gajjar, N.A.; Hiatt, J.; Durazo, F.; Han, S.; Farmer, D.G.; Ghobrial, R.M.; Yersiz, H.; Goldstein, L.I.; et al. Outcomes of acute rejection after interferon therapy in liver transplant recipients. Liver Transpl. 2004, 10, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Peters van Ton, A.M.; Gevers, T.J.; Drenth, J.P. Antiviral therapy in chronic hepatitis E: A systematic review. J. Viral Hepat. 2015, 22, 965–973. [Google Scholar] [CrossRef]
- Debing, Y.; Emerson, S.U.; Wang, Y.; Pan, Q.; Balzarini, J.; Dallmeier, K.; Neyts, J. Ribavirin inhibits in vitro hepatitis E virus replication through depletion of cellular GTP pools and is moderately synergistic with alpha interferon. Antimicrob. Agents Chemother. 2014, 58, 267–273. [Google Scholar] [CrossRef]
- de Clercq, E. Strategies in the design of antiviral drugs. Nat. Rev. Drug Discov. 2002, 1, 13–25. [Google Scholar] [CrossRef]
- Roda, A.; Guardigli, M.; Pasini, P.; Mirasoli, M. Bioluminescence and chemiluminescence in drug screening. Anal. Bioanal. Chem. 2003, 377, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Eckert, N.; Wrensch, F.; Gartner, S.; Palanisamy, N.; Goedecke, U.; Jager, N.; Pohlmann, S.; Winkler, M. Influenza A virus encoding secreted Gaussia luciferase as useful tool to analyze viral replication and its inhibition by antiviral compounds and cellular proteins. PLoS ONE 2014, 9, e97695. [Google Scholar] [CrossRef]
- Nishiyama, T.; Kobayashi, T.; Jirintai, S.; Nagashima, S.; Primadharsini, P.P.; Nishizawa, T.; Okamoto, H. Antiviral candidates against the hepatitis E virus (HEV) and their combinations inhibit HEV growth in in vitro. Antiviral. Res. 2019, 170, 104570. [Google Scholar] [CrossRef]
- Nishiyama, T.; Kobayashi, T.; Jirintai, S.; Kii, I.; Nagashima, S.; Primadharsini, P.P.; Nishizawa, T.; Okamoto, H. Screening of novel drugs for inhibiting hepatitis E virus replication. J. Virol. Methods 2019, 270, 1–11. [Google Scholar] [CrossRef]
- Primadharsini, P.P.; Nagashima, S.; Nishiyama, T.; Takahashi, M.; Murata, K.; Okamoto, H. Development of recombinant infectious hepatitis E virus harboring the nanoKAZ gene and its application in drug screening. J. Virol. 2022, 96, e0190621. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Primadharsini, P.P.; Nishiyama, T.; Takahashi, M.; Murata, K.; Okamoto, H. Development of a HiBiT-tagged reporter hepatitis E virus and its utility as an antiviral drug screening platform. J. Virol. 2023, online ahead of print, e0050823. [Google Scholar] [CrossRef]
- Nagashima, S.; Kobayashi, T.; Tanaka, T.; Tanggis; Jirintai, S.; Takahashi, M.; Nishizawa, T.; Okamoto, H. Analysis of adaptive mutations selected during the consecutive passages of hepatitis E virus produced from an infectious cDNA clone. Virus Res. 2016, 223, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for hepatitis E virus. J. Gen. Virol. 2007, 88, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Primadharsini, P.P.; Nagashima, S.; Takahashi, M.; Murata, K.; Okamoto, H. Ritonavir blocks hepatitis E virus internalization and clears hepatitis E virus in vitro with ribavirin. Viruses 2022, 14, 2440. [Google Scholar] [CrossRef]
- England, C.G.; Ehlerding, E.B.; Cai, W. NanoLuc: A small luciferase is brightening up the field of bioluminescence. Bioconjug. Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Shukla, P.; Torian, U.; Faulk, K.; Emerson, S.U. Hepatitis E virus genotype 1 infection of swine kidney cells in vitro is inhibited at multiple levels. J. Virol. 2014, 88, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Debing, Y.; Ramiere, C.; Dallmeier, K.; Piorkowski, G.; Trabaud, M.A.; Lebosse, F.; Scholtes, C.; Roche, M.; Legras-Lachuer, C.; de Lamballerie, X.; et al. Hepatitis E virus mutations associated with ribavirin treatment failure result in altered viral fitness and ribavirin sensitivity. J. Hepatol. 2016, 65, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Debing, Y.; Zhou, X.; Yin, Y.; Xu, L.; Herrera Carrillo, E.; Brandsma, J.H.; Poot, R.A.; Berkhout, B.; et al. Biological or pharmacological activation of protein kinase C alpha constrains hepatitis E virus replication. Antiviral. Res. 2017, 140, 1–12. [Google Scholar] [CrossRef]
- Todt, D.; Friesland, M.; Moeller, N.; Praditya, D.; Kinast, V.; Bruggemann, Y.; Knegendorf, L.; Burkard, T.; Steinmann, J.; Burm, R.; et al. Robust hepatitis E virus infection and transcriptional response in human hepatocytes. Proc. Natl. Acad. Sci. USA 2020, 117, 1731–1741. [Google Scholar] [CrossRef]
- Shukla, P.; Nguyen, H.T.; Faulk, K.; Mather, K.; Torian, U.; Engle, R.E.; Emerson, S.U. Adaptation of a genotype 3 hepatitis E virus to efficient growth in cell culture depends on an inserted human gene segment acquired by recombination. J. Virol. 2012, 86, 5697–5707. [Google Scholar] [CrossRef]
- Ding, Q.; Nimgaonkar, I.; Archer, N.F.; Bram, Y.; Heller, B.; Schwartz, R.E.; Ploss, A. Identification of the intragenomic promoter controlling hepatitis E virus subgenomic RNA transcription. mBio 2018, 9, e00769-18. [Google Scholar] [CrossRef]
- Debing, Y.; Mishra, N.; Verbeken, E.; Ramaekers, K.; Dallmeier, K.; Neyts, J. A rat model for hepatitis E virus. Dis. Model Mech. 2016, 9, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Shiota, T.; Li, T.C.; Yoshizaki, S.; Kato, T.; Wakita, T.; Ishii, K. The hepatitis E virus capsid C-terminal region is essential for the viral life cycle: Implication for viral genome encapsidation and particle stabilization. J. Virol. 2013, 87, 6031–6036. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Mahsoub, H.M.; Li, W.; Heffron, C.L.; Tian, D.; Hassebroek, A.M.; LeBroith, T.; Meng, X.J. Ribavirin treatment failure-associated mutation, Y1320H, in the RNA-dependent RNA polymerase of genotype 3 hepatitis E virus (HEV) enhances virus replication in a rabbit HEV infection model. mBio. 2023, 14, e0337222. [Google Scholar] [CrossRef]
- Metzger, K.; Bentaleb, C.; Hervouet, K.; Alexandre, V.; Montpellier, C.; Saliou, J.M.; Ferrie, M.; Camuzet, C.; Rouille, Y.; Lecoeur, C.; et al. Processing and subcellular localization of the hepatitis E virus replicase: Identification of candidate viral factories. Front. Microbiol. 2022, 13, 828636. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Li, Y.; Li, Y.; Yu, P.; Li, P.; Donkers, J.M.; van de Graaf, S.F.J.; de Man, R.A.; Peppelenbosch, M.P.; Pan, Q. FDA-drug screening identifies deptropine inhibiting hepatitis E virus involving the NF-kappaB-RIPK1-caspase axis. Antiviral Res. 2019, 170, 104588. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, P.; Li, Y.; Zhang, R.; Yu, P.; Ma, Z.; Kainov, D.E.; de Man, R.A.; Peppelenbosch, M.P.; Pan, Q. Drug screening identified gemcitabine inhibiting hepatitis E virus by inducing interferon-like response via activation of STAT1 phosphorylation. Antiviral Res. 2020, 184, 104967. [Google Scholar] [CrossRef]
- Nimgaonkar, I.; Archer, N.F.; Becher, I.; Shahrad, M.; LeDesma, R.A.; Mateus, A.; Caballero-Gomez, J.; Berneshawi, A.R.; Ding, Q.; Douam, F.; et al. Isocotoin suppresses hepatitis E virus replication through inhibition of heat shock protein 90. Antiviral Res. 2021, 185, 104997. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, X.; Debing, Y.; Chen, K.; van der Laan, L.J.; Neyts, J.; Janssen, H.L.; Metselaar, H.J.; Peppelenbosch, M.P.; Pan, Q. Calcineurin inhibitors stimulate and mycophenolic acid inhibits replication of hepatitis E virus. Gastroenterology 2014, 146, 1775–1783. [Google Scholar] [CrossRef]
- Dao Thi, V.L.; Debing, Y.; Wu, X.; Rice, C.M.; Neyts, J.; Moradpour, D.; Gouttenoire, J. Sofosbuvir inhibits hepatitis E virus replication in vitro and results in an additive effect when combined with ribavirin. Gastroenterology 2016, 150, 82–85.e84. [Google Scholar] [CrossRef]
- Kamar, N.; Abravanel, F.; Behrendt, P.; Hofmann, J.; Pageaux, G.P.; Barbet, C.; Moal, V.; Couzi, L.; Horvatits, T.; de Man, R.A.; et al. Ribavirin for hepatitis E virus infection after organ transplantation: A large European retrospective multicenter study. Clin. Inf. Dis. 2020, 71, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Gorris, M.; van der Lecq, B.M.; van Erpecum, K.J.; de Bruijne, J. Treatment for chronic hepatitis E virus infection: A systematic review and meta-analysis. J. Viral Hepat. 2021, 28, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Xu, L.; Yin, Y.; Peppelenbosch, M.P.; Pan, Q.; Wang, W. Nucleoside analogue 2’-C-methylcytidine inhibits hepatitis E virus replication but antagonizes ribavirin. Arch. Virol. 2017, 162, 2989–2996. [Google Scholar] [CrossRef] [PubMed]
- van der Valk, M.; Zaaijer, H.L.; Kater, A.P.; Schinkel, J. Sofosbuvir shows antiviral activity in a patient with chronic hepatitis E virus infection. J. Hepatol. 2017, 66, 242–243. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Sato, J.; Sahara-Miura, Y.; Yoshida, S.; Kurakata, H.; Hosoya, T. C6-deoxy coelenterazine analogues as an efficient substrate for glow luminescence reaction of nanoKAZ: The mutated catalytic 19 kDa component of Oplophorus luciferase. Biochem. Biophys. Res. Commun. 2013, 437, 23–28. [Google Scholar] [CrossRef]
- Inouye, S.; Sato, J.; Sahara-Miura, Y.; Yoshida, S.; Hosoya, T. Luminescence enhancement of the catalytic 19 kDa protein (KAZ) of Oplophorus luciferase by three amino acid substitutions. Biochem. Biophys. Res. Commun. 2014, 445, 157–162. [Google Scholar] [CrossRef]
- Inouye, S.; Sato, J.; Sahara-Miura, Y.; Hosoya, T.; Suzuki, T. Unconventional secretion of the mutated 19 kDa protein of Oplophorus luciferase (nanoKAZ) in mammalian cells. Biochem. Biophys. Res. Commun. 2014, 450, 1313–1319. [Google Scholar] [CrossRef]
- Tomabechi, Y.; Hosoya, T.; Ehara, H.; Sekine, S.I.; Shirouzu, M.; Inouye, S. Crystal structure of nanoKAZ: The mutated 19 kDa component of Oplophorus luciferase catalyzing the bioluminescent reaction with coelenterazine. Biochem. Biophys. Res. Commun. 2016, 470, 88–93. [Google Scholar] [CrossRef]
- Bouquet, J.; Cheval, J.; Rogee, S.; Pavio, N.; Eloit, M. Identical consensus sequence and conserved genomic polymorphism of hepatitis E virus during controlled interspecies transmission. J. Virol. 2012, 86, 6238–6245. [Google Scholar] [CrossRef]
- Takeuchi, S.; Yamazaki, Y.; Sato, K.; Takizawa, D.; Yamada, M.; Okamoto, H. Full genome analysis of a European-type genotype 3 hepatitis E virus variant obtained from a Japanese patient with autochthonous acute hepatitis E. J. Med. Virol. 2015, 87, 1067–1071. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Torian, U.; Faulk, K.; Mather, K.; Engle, R.E.; Thompson, E.; Bonkovsky, H.L.; Emerson, S.U. A naturally occurring human/hepatitis E recombinant virus predominates in serum but not in faeces of a chronic hepatitis E patient and has a growth advantage in cell culture. J. Gen. Virol. 2012, 93, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, S.; Abravanel, F.; Dubois, M.; Sandres-Saune, K.; Mansuy, J.M.; Rostaing, L.; Kamar, N.; Izopet, J. Characterization of the polyproline region of the hepatitis E virus in immunocompromised patients. J. Virol. 2014, 88, 12017–12025. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Reetz, J.; Ulrich, R.G.; Machnowska, P.; Sachsenroder, J.; Nickel, P.; Hofmann, J. An ORF1-rearranged hepatitis E virus derived from a chronically infected patient efficiently replicates in cell culture. J. Viral Hepat. 2014, 21, 447–456. [Google Scholar] [CrossRef]
- Peralta, B.; Mateu, E.; Casas, M.; de Deus, N.; Martin, M.; Pina, S. Genetic characterization of the complete coding regions of genotype 3 hepatitis E virus isolated from Spanish swine herds. Virus Res. 2009, 139, 111–116. [Google Scholar] [CrossRef]
- Szkolnicka, D.; Pollan, A.; da Silva, N.; Oechslin, N.; Gouttenoire, J.; Moradpour, D. Recombinant hepatitis E viruses harboring tags in the ORF1 protein. J. Virol. 2019, 93, e00459-19. [Google Scholar] [CrossRef]
- Zhang, F.; Guo, H.; Zhang, J.; Chen, Q.; Fang, Q. Identification of the caveolae/raft-mediated endocytosis as the primary entry pathway for aquareovirus. Virology 2018, 513, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Zhang, R.; Yu, P.; Li, Y.; Pan, Q.; Li, Y. The macrolide antibiotic azithromycin potently inhibits hepatitis E virus in cell culture models. Int. J. Antimicrob. Agents 2021, 58, 106383. [Google Scholar] [CrossRef]
- Schwinn, M.K.; Machleidt, T.; Zimmerman, K.; Eggers, C.T.; Dixon, A.S.; Hurst, R.; Hall, M.P.; Encell, L.P.; Binkowski, B.F.; Wood, K.V. CRISPR-mediated tagging of endogenous proteins with a luminescent peptide. ACS Chem. Biol. 2018, 13, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, G.; Zhang, Y.; Lin, X.; Zhao, X.; Cui, Q.; Rong, L.; Du, R. Development of an HiBiT-tagged reporter H3N2 influenza A virus and its utility as an antiviral screening platform. J. Med. Virol. 2023, 95, e28345. [Google Scholar] [CrossRef]
- Nishiyama, T.; Umezawa, K.; Yamada, K.; Takahashi, M.; Kunita, S.; Mulyanto; Kii, I.; Okamoto, H. The capsid (ORF2) protein of hepatitis E virus in feces is C-terminally truncated. Pathogens 2021, 11, 24. [Google Scholar] [CrossRef]
- Graff, J.; Nguyen, H.; Yu, C.; Elkins, W.R.; St Claire, M.; Purcell, R.H.; Emerson, S.U. The open reading frame 3 gene of hepatitis E virus contains a cis-reactive element and encodes a protein required for infection of macaques. J. Virol. 2005, 79, 6680–6689. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.; Xiang, G.; Gong, M.; Yang, R.; Qin, J.; Li, Y.; Nan, Y.; Yang, Y.; Zhang, Q.C.; Ding, Q. Identification of functional cis-acting RNA elements in the hepatitis E virus genome required for viral replication. PLoS Pathog. 2020, 16, e1008488. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Structural and molecular biology of hepatitis E virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Nagashima, S.; Jirintai, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Kouki, T.; Yashiro, T.; Okamoto, H. Hepatitis E virus egress depends on the exosomal pathway, with secretory exosomes derived from multivesicular bodies. J. Gen. Virol. 2014, 95, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Takahashi, M.; Jirintai, S.; Tanaka, T.; Nishizawa, T.; Yasuda, J.; Okamoto, H. Tumour susceptibility gene 101 and the vacuolar protein sorting pathway are required for the release of hepatitis E virions. J. Gen. Virol. 2011, 92, 2838–2848. [Google Scholar] [CrossRef] [PubMed]





| Strain (Genotype) | Insertion Site | Strategy to Select Insertion Site | Tag | Application | Reference |
|---|---|---|---|---|---|
| HEV83-2-27 (HEV-3) | HVR | Transposon-mediated random insertion coupled with selection in a subgenomic replicon system | Hemagglutinin or NanoLuc |
| [96] |
| JE03-1760F/P10 (HEV-3) | HVR | Based on insertion sites that have been reported in HEV-3-infected humans or pigs with ≥30-aa insertion | nanoKAZ |
| [61] |
| Strain (Genotype) | Tag | Strategy | Application | Findings | Reference |
|---|---|---|---|---|---|
| JE03-1760F/P10 (HEV-3) | FLAG | Two tandem glycine–serine linker sequences and FLAG tag were inserted at the 3′ end of the ORF2 sequence, followed by two stop codons and the 3′-terminal 60 nt ORF2 sequence (nt 7091–7151). |
| ORF2 proteins associated with enveloped HEV have an intact C-terminus, but those associated with non-enveloped HEV are C-terminally truncated. | [101] |
| JE03-1760F/P10 (HEV-3) | HiBiT | Two tandem glycine–serine linker sequences and HiBiT tag were inserted at the 3′ end of the ORF2 sequence, followed by two stop codons and the 3′-terminal 60 nt ORF2 sequence (nt 7091–7151). |
| The reporter HEV replicated efficiently in PLC/PRF/5 cells, produced membrane-associated particles, and was genetically stable and infectious. | [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Primadharsini, P.P.; Nagashima, S.; Nishiyama, T.; Okamoto, H. Three Distinct Reporter Systems of Hepatitis E Virus and Their Utility as Drug Screening Platforms. Viruses 2023, 15, 1989. https://doi.org/10.3390/v15101989
Primadharsini PP, Nagashima S, Nishiyama T, Okamoto H. Three Distinct Reporter Systems of Hepatitis E Virus and Their Utility as Drug Screening Platforms. Viruses. 2023; 15(10):1989. https://doi.org/10.3390/v15101989
Chicago/Turabian StylePrimadharsini, Putu Prathiwi, Shigeo Nagashima, Takashi Nishiyama, and Hiroaki Okamoto. 2023. "Three Distinct Reporter Systems of Hepatitis E Virus and Their Utility as Drug Screening Platforms" Viruses 15, no. 10: 1989. https://doi.org/10.3390/v15101989
APA StylePrimadharsini, P. P., Nagashima, S., Nishiyama, T., & Okamoto, H. (2023). Three Distinct Reporter Systems of Hepatitis E Virus and Their Utility as Drug Screening Platforms. Viruses, 15(10), 1989. https://doi.org/10.3390/v15101989