
The Modulation of Immune Responses in Tilapinevirus tilapiae-Infected Fish Cells through MAPK/ERK Signalling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Infection
2.2. Inhibition of MAPK-ERK Signalling
2.3. Western Blot Analysis
2.4. RNA Extraction, cDNA Construction and qPCR
2.5. Statistical Analysis
3. Results
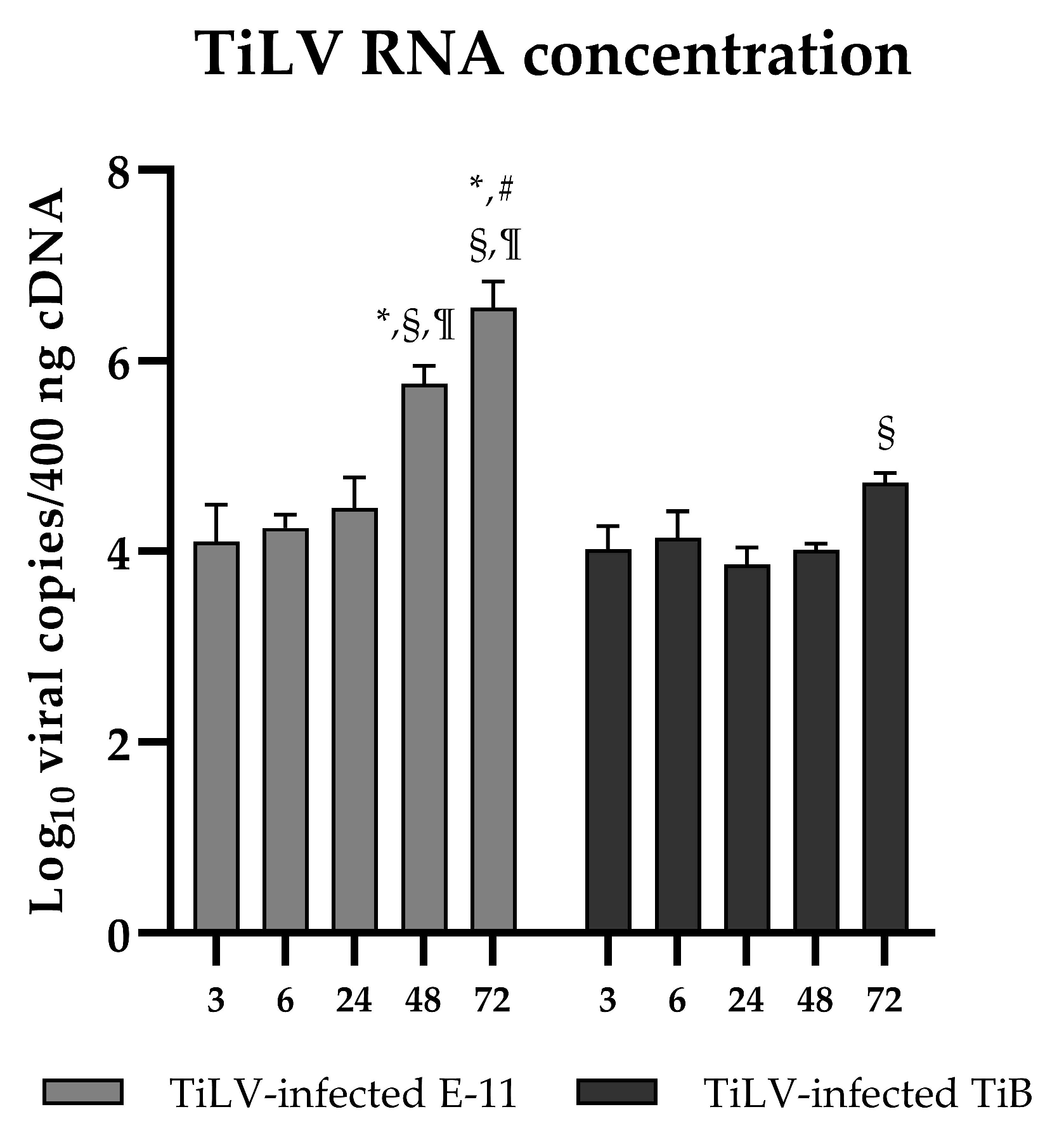
3.1. Different Cell Responses to TiLV Infection in E-11 and TiB Cells
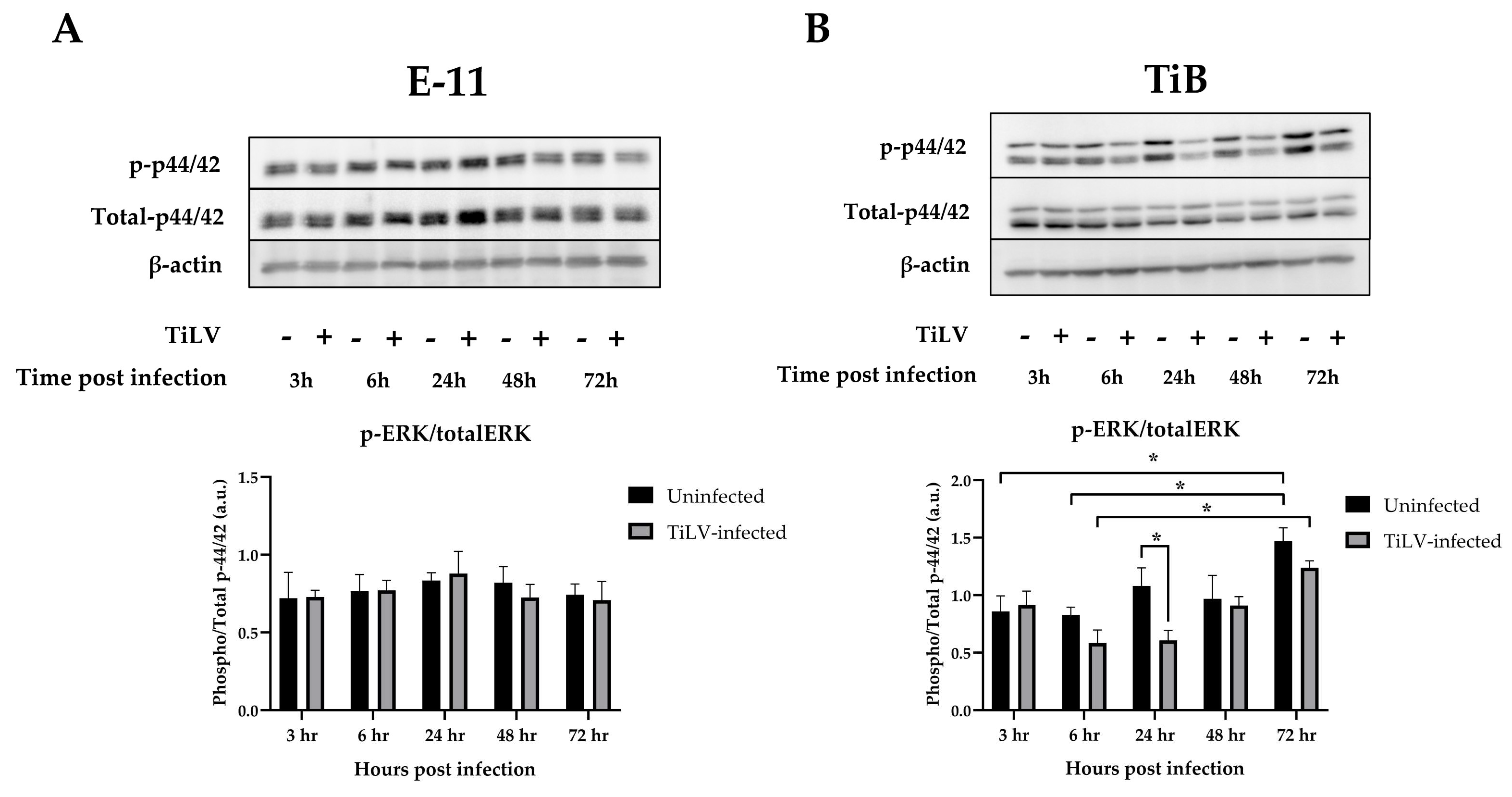
3.2. Inihibition of ERK Phosphorylation in TiB Cells following TiLV Infection
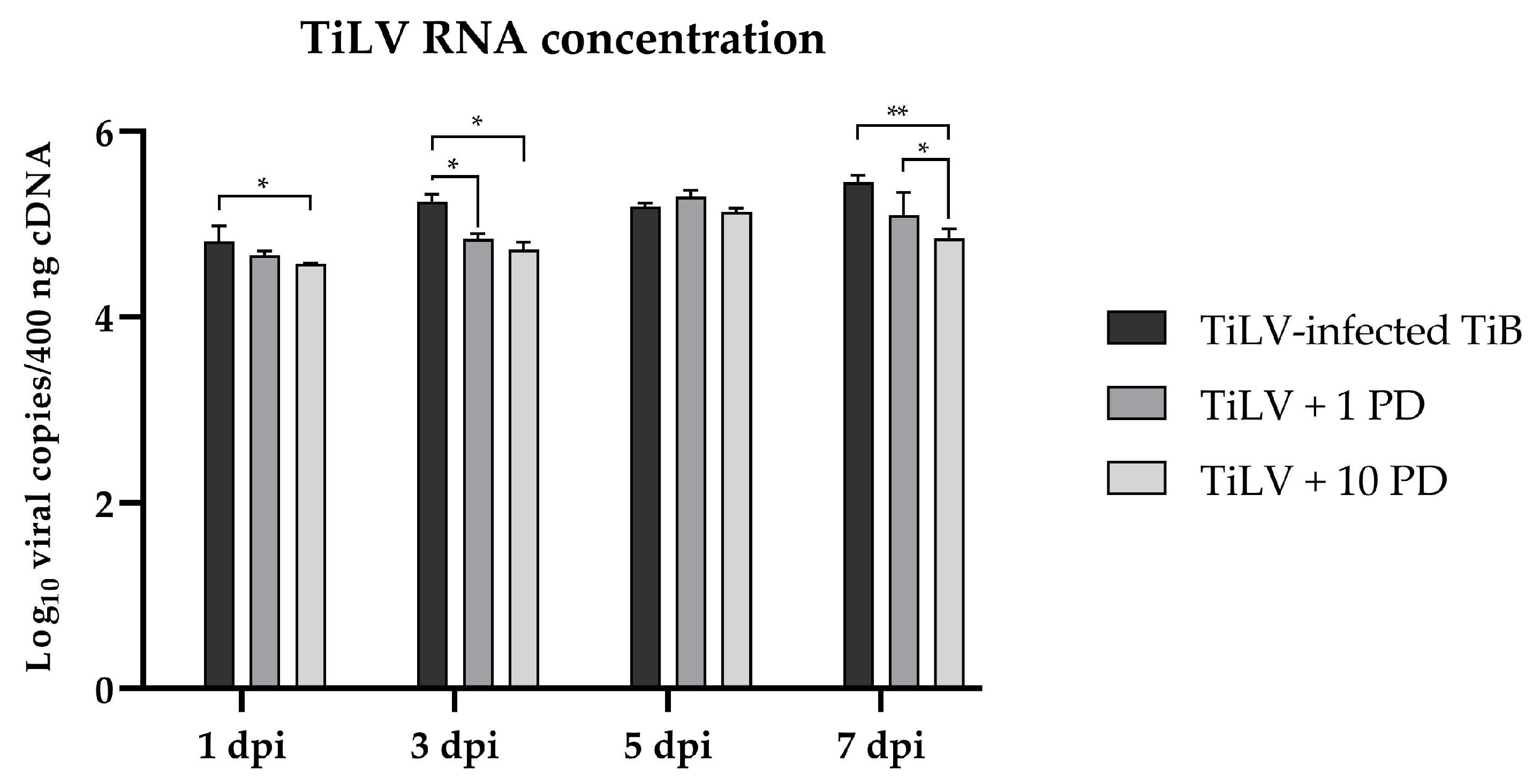
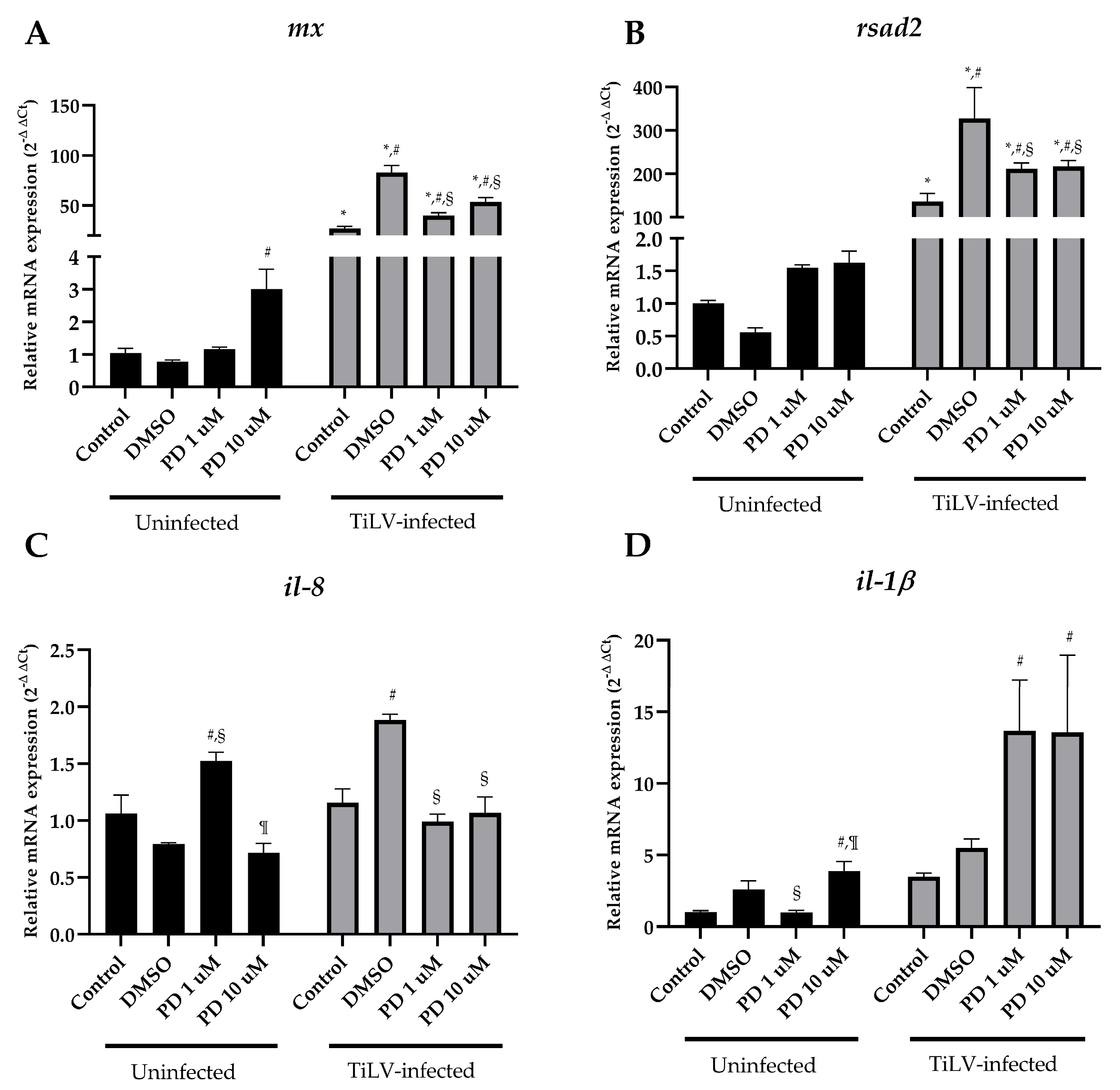
3.3. Suppression of ERK-Signalling Reduces Viral Copies and Altered Immune-Related Genes during Early TiLV Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Debnath, P.P.; Dinh-Hung, N.; Taengphu, S.; Nguyen, V.V.; Delamare-Deboutteville, J.; Senapin, S.; Vishnumurthy Mohan, C.; Dong, H.T.; Rodkhum, C. Tilapia Lake virus was not detected in non-tilapine species within tilapia polyculture systems of Bangladesh. J. Fish. Dis. 2022, 45, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Surachetpong, W.; Roy, S.R.K.; Nicholson, P. Tilapia Lake virus: The story so far. J. Fish. Dis. 2020, 43, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.; Nguyen, V.T.H.; Bui, H.C.N.; Tran, Y.B.T.; Tran, H.T.T.; Le, T.T.T.; Vu, H.T.T.; Ngo, T.P.H. Tilapia Lake Virus (TiLV) from Vietnam is genetically distantly related to TiLV strains from other countries. J. Fish. Dis. 2022, 45, 1389–1401. [Google Scholar] [CrossRef]
- ICTV Virus Taxonomy: 2021 Release. Available online: https://ictv.global/taxonomy (accessed on 22 February 2022).
- Eyngor, M.; Zamostiano, R.; Kembou Tsofack, J.E.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E.; et al. Identification of a novel RNA virus lethal to tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, H.W.; Kabuusu, R.; Beltran, S.; Reyes, E.; Lince, J.A.; del Pozo, J. Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): A case report. J. Fish. Dis. 2014, 37, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.K.; Sakulpolwat, S.; Sukdanon, S.; Lertwanakarn, T.; Waiyamitra, P.; Piewbang, C.; Pierezan, F.; Techangamsuwan, S.; Soto, E.; Surachetpong, W. Tilapia Lake virus (TiLV) causes severe anaemia and systemic disease in tilapia. J. Fish. Dis. 2023. [Google Scholar] [CrossRef]
- He, T.; Zhang, Y.-Z.; Gao, L.-H.; Miao, B.; Zheng, J.-S.; Pu, D.-C.; Zhang, Q.-Q.; Zeng, W.-W.; Wang, D.-S.; Su, S.-Q.; et al. Identification and pathogenetic study of tilapia lake virus (TiLV) isolated from naturally diseased tilapia. Aquaculture 2023, 565, 739166. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Dachavichitlead, W.; Surachetpong, W. Experimental infection of Tilapia Lake virus (TiLV) in Nile tilapia (Oreochromis niloticus) and red tilapia (Oreochromis spp.). Vet. Microbiol. 2017, 207, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zheng, S.; Wang, Y.; Wang, Q.; Li, Y.; Yin, J.; Ren, Y.; Shi, C.; Zhao, Z.; Jiang, Z.; et al. Susceptibilities of ten fish cell lines to infection with Tilapia Lake virus. Microb. Pathog. 2022, 166, 105510. [Google Scholar] [CrossRef]
- Thangaraj, R.S.; Ravi, C.; Kumar, R.; Dharmaratnam, A.; Valaparambil Saidmuhammed, B.; Pradhan, P.K.; Sood, N. Derivation of two tilapia (Oreochromis niloticus) cell lines for efficient propagation of Tilapia Lake Virus (TiLV). Aquaculture 2018, 492, 206–214. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Zeng, W.; Yin, J.; Li, Y.; Ren, Y.; Shi, C.; Bergmann, S.M.; Zhu, X. Establishment and characterization of a cell line from tilapia brain for detection of Tilapia Lake virus. J. Fish. Dis. 2018, 41, 1803–1809. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Rastogi, A.; Criollo Joaquin, M.P.; Verma, D.K.; Rathore, G.; Swaminathan, T.R.; Paria, A.; Pradhan, P.K.; Sood, N. Establishment and characterization of a continuous cell line from heart of Nile tilapia (Oreochromis niloticus) and its susceptibility to tilapia lake virus. J. Virol. Methods 2021, 287, 113989. [Google Scholar] [CrossRef]
- Waiyamitra, P.; Zoral, M.A.; Saengtienchai, A.; Luengnaruemitchai, A.; Decamp, O.; Gorgoglione, B.; Surachetpong, W. Probiotics modulate tilapia resistance and immune response against Tilapia Lake virus infection. Pathogens 2020, 9, 919. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Yamkasem, J.; Surachetpong, W.; Lin, Y.J.; You, S.H.; Lu, T.H.; Chen, C.Y.; Wang, W.M.; Liao, C.M. Assessing the effect of probiotics on tilapia lake virus-infected tilapia: Transmission and immune response. J. Fish. Dis. 2022, 45, 1117–1132. [Google Scholar] [CrossRef] [PubMed]
- Lertwanakarn, T.; Trongwongsa, P.; Yingsakmongkol, S.; Khemthong, M.; Tattiyapong, P.; Surachetpong, W. Antiviral Activity of Ribavirin against Tilapia tilapinevirus in Fish Cells. Pathogens 2021, 10, 1616. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Kitiyodom, S.; Yata, T.; Jantharadej, K.; Adamek, M.; Surachetpong, W. Chitosan nanoparticle immersion vaccine offers protection against Tilapia Lake virus in laboratory and field studies. Fish. Shellfish. Immunol. 2022, 131, 972–979. [Google Scholar] [CrossRef]
- Yu, N.-T.; Zeng, W.-W.; Xiong, Z.; Liu, Z.-X. A high efficacy DNA vaccine against Tilapia Lake virus in Nile tilapia (Oreochromis niloticus). Aquac. Rep. 2022, 24, 101166. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, Y.; Chen, X.; Wang, Q.; Bergmann, S.M.; Yang, Y.; Wang, Y.; Li, B.; Lv, Y.; Li, H.; et al. Potency and efficacy of VP20-based vaccine against tilapia lake virus using different prime-boost vaccination regimens in tilapia. Aquaculture 2021, 539, 736654. [Google Scholar] [CrossRef]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Shepherd, E.G.; Nelin, L.D. MAPK phosphatases—Regulating the immune response. Nat. Rev. Immunol. 2007, 7, 202–212. [Google Scholar] [CrossRef]
- Qiang, J.; Tao, Y.-F.; Zhu, J.-H.; Lu, S.-Q.; Cao, Z.-M.; Ma, J.-L.; He, J.; Xu, P. Effects of heat stress on follicular development and atresia in Nile tilapia (Oreochromis niloticus) during one reproductive cycle and its potential regulation by autophagy and apoptosis. Aquaculture 2022, 555, 738171. [Google Scholar] [CrossRef]
- Xu, Z.; Gan, L.; Li, T.; Xu, C.; Chen, K.; Wang, X.; Qin, J.G.; Chen, L.; Li, E. Transcriptome profiling and molecular pathway analysis of genes in association with salinity adaptation in Nile tilapia (Oreochromis niloticus). PLoS ONE 2015, 10, e0136506. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Megens, H.J.; Mengistu, S.B.; Bastiaansen, J.W.M.; Mulder, H.A.; Benzie, J.A.H.; Groenen, M.A.M.; Komen, H. Genome-wide association analysis of adaptation to oxygen stress in Nile tilapia (Oreochromis niloticus). BMC Genomics 2021, 22, 426. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Y.; Li, C.; Ai, K.; Li, K.; Li, H.; Yang, J. The evolutionarily conserved MAPK/Erk signaling promotes ancestral T-cell immunity in fish via c-Myc-mediated glycolysis. J. Biol. Chem. 2020, 295, 3000–3016. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.-G.; Zou, Z.-Y.; Xiao, W.; Zhu, J.-L.; Li, D.-Y.; Yu, J.; Yang, H.; Chen, B.-L.; Ma, Y.-H. Antibacterial ability of two species of tilapia is related to inducibility of the p38 mitogen-activated protein kinase pathway. Aquac. Res. 2021, 52, 456–462. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, Y.; Zhang, X. Suppression of coronavirus replication by inhibition of the MEK signaling pathway. J. Virol. 2007, 81, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Khandelwal, N.; Thachamvally, R.; Tripathi, B.N.; Barua, S.; Kashyap, S.K.; Maherchandani, S.; Kumar, N. Role of MAPK/MNK1 signaling in virus replication. Virus Res. 2018, 253, 48–61. [Google Scholar] [CrossRef]
- Luo, H.; Yanagawa, B.; Zhang, J.; Luo, Z.; Zhang, M.; Esfandiarei, M.; Carthy, C.; Wilson, J.E.; Yang, D.; McManus, B.M. Coxsackievirus B3 replication is reduced by inhibition of the extracellular signal-regulated kinase (ERK) signaling pathway. J. Virol. 2002, 76, 3365–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Márquez, N.; Calzado, M.A.; Sánchez-Duffhues, G.; Pérez, M.; Minassi, A.; Pagani, A.; Appendino, G.; Diaz, L.; Muñoz-Fernández, M.A.; Muñoz, E. Differential effects of phorbol-13-monoesters on human immunodeficiency virus reactivation. Biochem. Pharmacol. 2008, 75, 1370–1380. [Google Scholar] [CrossRef]
- Pleschka, S. RNA viruses and the mitogenic Raf/MEK/ERK signal transduction cascade. Biol. Chem. 2008, 389, 1273–1282. [Google Scholar] [CrossRef]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza virus propagation is impaired by inhibition of the Raf/MEK/ERK signalling cascade. Nat. Cell. Biol. 2001, 3, 301–305. [Google Scholar] [CrossRef]
- Yang, X.; Chen, Y.; Gabuzda, D. ERK MAP kinase links cytokine signals to activation of latent HIV-1 infection by stimulating a cooperative interaction of AP-1 and NF-kappaB. J. Biol. Chem. 1999, 274, 27981–27988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piewbang, C.; Tattiyapong, P.; Khemthong, M.; Lachroje, S.; Boonrungsiman, S.; Kasantikul, T.; Surachetpong, W.; Techangamsuwan, S. Dual infections of tilapia parvovirus (TiPV) and tilapia lake virus (TiLV) in multiple tilapia farms: Their impacts, genetic diversity, viral tropism, and pathological effects. Aquaculture 2022, 550, 737887. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Sirikanchana, K.; Surachetpong, W. Development and validation of a reverse transcription quantitative polymerase chain reaction for Tilapia Lake virus detection in clinical samples and experimentally challenged fish. J. Fish. Dis. 2018, 41, 255–261. [Google Scholar] [CrossRef]
- Sood, N.; Verma, D.K.; Paria, A.; Yadav, S.C.; Yadav, M.K.; Bedekar, M.K.; Kumar, S.; Swaminathan, T.R.; Mohan, C.V.; Rajendran, K.V.; et al. Transcriptome analysis of liver elucidates key immune-related pathways in Nile tilapia Oreochromis niloticus following infection with Tilapia Lake virus. Fish. Shellfish. Immunol. 2021, 111, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Abu Rass, R.; Kembou-Ringert, J.E.; Zamostiano, R.; Eldar, A.; Ehrlich, M.; Bacharach, E. Mapping of Tilapia Lake virus entry pathways with inhibitors reveals dependence on dynamin activity and cholesterol but not endosomal acidification. Front. Cell. Dev. Biol. 2022, 10, 1075364. [Google Scholar] [CrossRef]
- Zampieri, C.A.; Fortin, J.F.; Nolan, G.P.; Nabel, G.J. The ERK mitogen-activated protein kinase pathway contributes to Ebola virus glycoprotein-induced cytotoxicity. J. Virol. 2007, 81, 1230–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planz, O.; Pleschka, S.; Ludwig, S. MEK-specific inhibitor U0126 blocks spread of Borna disease virus in cultured cells. J. Virol. 2001, 75, 4871–4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planz, O.; Pleschka, S.; Wolff, T. Borna disease virus: A unique pathogen and its interaction with intracellular signalling pathways. Cell. Microbiol. 2009, 11, 872–879. [Google Scholar] [CrossRef]
- Barber, S.A.; Bruett, L.; Douglass, B.R.; Herbst, D.S.; Zink, M.C.; Clements, J.E. Visna virus-induced activation of MAPK is required for virus replication and correlates with virus-induced neuropathology. J. Virol. 2002, 76, 817–828. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; Hijikata, M.; Shimotohno, K. Enhancement of internal ribosome entry site-mediated translation and replication of hepatitis C virus by PD98059. Virology 2005, 340, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Kuang, E.; Fu, B.; Liang, Q.; Myoung, J.; Zhu, F. Phosphorylation of eukaryotic translation initiation factor 4B (EIF4B) by open reading frame 45/p90 ribosomal S6 kinase (ORF45/RSK) signaling axis facilitates protein translation during Kaposi sarcoma-associated herpesvirus (KSHV) lytic replication. J. Biol. Chem. 2011, 286, 41171–41182. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.D.; Sonenberg, N. Regulation of cap-dependent translation by eIF4E inhibitory proteins. Nature 2005, 433, 477–480. [Google Scholar] [CrossRef]
- Mugimba, K.K.; Lamkhannat, M.; Dubey, S.; Mutoloki, S.; Munang’andu, H.M.; Evensen, Ø. Tilapia Lake virus downplays innate immune responses during early stage of infection in Nile tilapia (Oreochromis niloticus). Sci. Rep. 2020, 10, 20364. [Google Scholar] [CrossRef]
- Pierezan, F.; Yun, S.; Piewbang, C.; Surachetpong, W.; Soto, E. Pathogenesis and immune response of Nile tilapia (Oreochromis niloticus) exposed to Tilapia Lake virus by intragastric route. Fish. Shellfish. Immunol. 2020, 107, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Rakus, K.; Mojzesz, M.; Widziolek, M.; Pooranachandran, N.; Teitge, F.; Surachetpong, W.; Chadzinska, M.; Steinhagen, D.; Adamek, M. Antiviral response of adult zebrafish (Danio rerio) during tilapia Lake virus (TiLV) infection. Fish. Shellfish. Immunol. 2020, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Villaizan, M.D.; Chico, V.; Perez, L. Fish innate immune response to viral infection—An overview of five major antiviral genes. Viruses 2022, 14, 1546. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhao, T.; Ai, K.; Zhang, Y.; Li, H.; Yang, J. c-Raf participates in adaptive immune response of Nile tilapia via regulating lymphocyte activation. Fish. Shellfish. Immunol. 2019, 86, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhao, T.; Zhang, Y.; Ai, K.; Li, H.; Yang, J. Involvement of H-Ras in the adaptive immunity of Nile tilapia by regulating lymphocyte activation. Fish. Shellfish. Immunol. 2019, 89, 281–289. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lertwanakarn, T.; Khemthong, M.; Tattiyapong, P.; Surachetpong, W. The Modulation of Immune Responses in Tilapinevirus tilapiae-Infected Fish Cells through MAPK/ERK Signalling. Viruses 2023, 15, 900. https://doi.org/10.3390/v15040900
Lertwanakarn T, Khemthong M, Tattiyapong P, Surachetpong W. The Modulation of Immune Responses in Tilapinevirus tilapiae-Infected Fish Cells through MAPK/ERK Signalling. Viruses. 2023; 15(4):900. https://doi.org/10.3390/v15040900
Chicago/Turabian StyleLertwanakarn, Tuchakorn, Matepiya Khemthong, Puntanut Tattiyapong, and Win Surachetpong. 2023. "The Modulation of Immune Responses in Tilapinevirus tilapiae-Infected Fish Cells through MAPK/ERK Signalling" Viruses 15, no. 4: 900. https://doi.org/10.3390/v15040900
APA StyleLertwanakarn, T., Khemthong, M., Tattiyapong, P., & Surachetpong, W. (2023). The Modulation of Immune Responses in Tilapinevirus tilapiae-Infected Fish Cells through MAPK/ERK Signalling. Viruses, 15(4), 900. https://doi.org/10.3390/v15040900