
Antiviral Activity of an Indole-Type Compound Derived from Natural Products, Identified by Virtual Screening by Interaction on Dengue Virus NS5 Protein

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Assays
2.1.1. Virtual Screening of Natural Compound Derivatives on DENV NS5 Protein
2.1.2. Selection of Compounds by Interaction on NS5 of DENV
2.2. In Vitro Assays
2.2.1. Determination of the Cytotoxic Effect of Compounds on Huh-7 Cells
2.2.2. Antiviral Screening of Compounds on NS1 Protein Production in DENV-2 Infected Cells
2.2.3. Determination of the Inhibitory Effect on NS5 Protein on the Expression Using ELISA Technique
2.2.4. Determination of the Inhibitory Effect on Viral RNA Production of DENV-2
2.3. Statistical Analysis
3. Results
3.1. The MTase and RdRp Domains Were Validated As Binding Sites
3.2. Selected Compounds by Interaction on MTase and RdRp of DENV NS5 Protein
3.3. Cytotoxic Effect Evaluation of Compounds in Huh-7 Cells
3.4. The Compounds Reduce the Production of NS1 Protein in Cells Infected with DENV-2
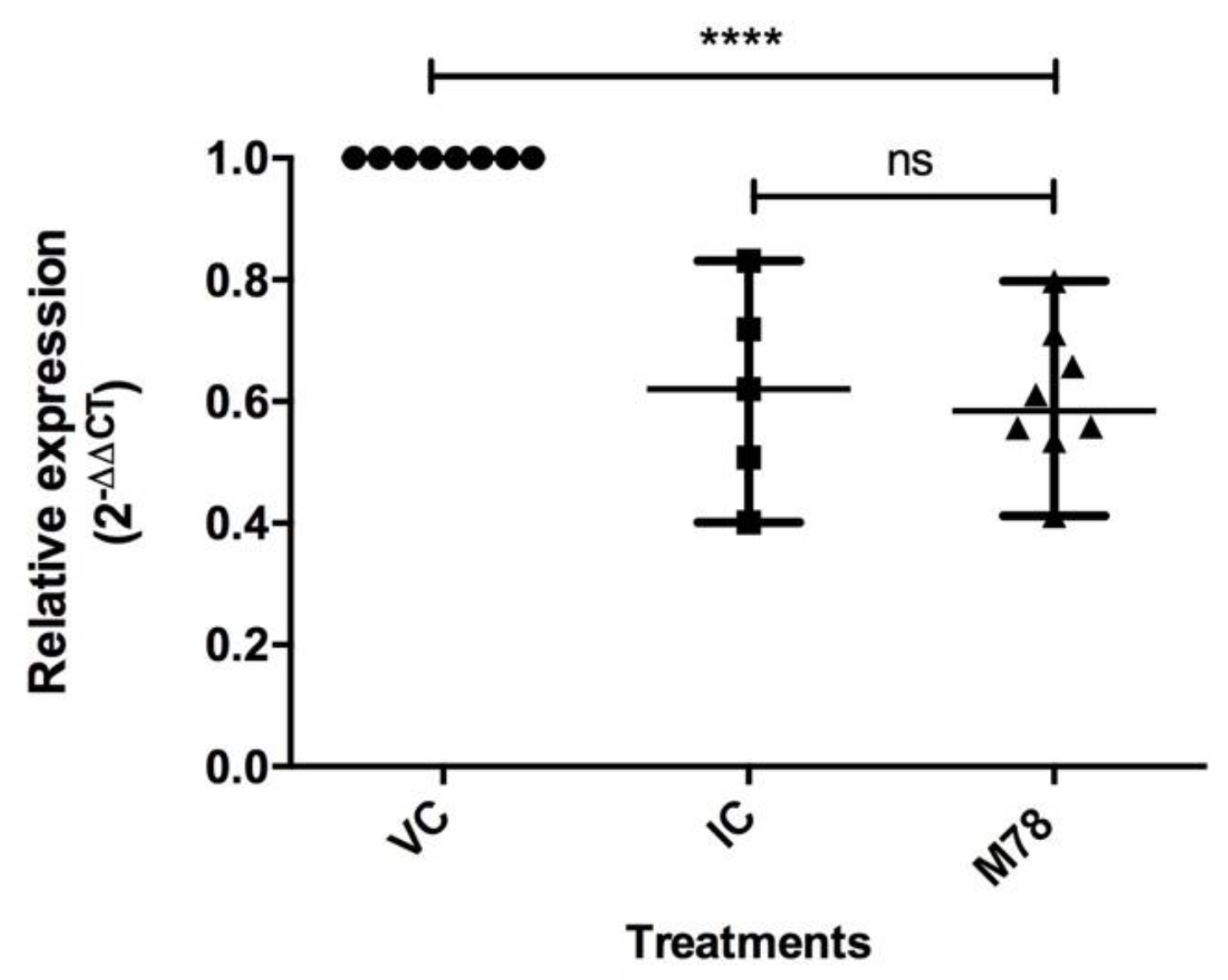
3.5. Compound M78 Affects NS5 Protein Production by Interfering with Genome Replication and/or Translation
3.6. The M78 Compound Affects The Production of DENV-2 Viral RNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uno, N.; Ross, T.M. Dengue virus and the host innate immune response. Emerg. Microbes Infect. 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bajrai, L.H.; Faizo, A.A.; Khateb, A.M.; Alkhaldy, A.A.; Rana, R.; Azhar, E.I.; Dwivedi, V.D. Pharmacophore-Model-Based Drug Repurposing for the Identification of the Potential Inhibitors Targeting the Allosteric Site in Dengue Virus NS5 RNA-Dependent RNA Polymerase. Viruses. 2022, 14, 1827. [Google Scholar] [CrossRef] [PubMed]
- Nazmi, A.; Dutta, K.; Hazra, B.; Basu, A. Role of pattern recognition receptors in flavivirus infections. Virus Res. 2014, 185, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Bhattacharjee, S. Dengue virus: Epidemiology, biology, and disease aetiology. Can. J. Microbiol. 2021, 67, 687–702. [Google Scholar] [CrossRef]
- Li, Y.; Dou, Q.; Lu, Y.; Xiang, H.; Yu, X.; Liu, S. Effects of ambient temperature and precipitation on the risk of dengue fever: A systematic review and updated meta-analysis. Environ. Res. 2020, 191, 110043. [Google Scholar] [CrossRef]
- Simmons, C.P.; Farrar, J.J.; Van Vinh Chau, N.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 423–1432. [Google Scholar] [CrossRef]
- Watterson, D.; Modhiran, N.; Young, P.R. The many faces of the flavivirus NS1 protein offer a multitude of options for inhibitor design. Antivir. Res. 2016, 130, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, P.O.; Chagas, M.A.; Rocha, W.R.; Moraes, A.H. Non-structural protein 5 (NS5) as a target for antiviral development against established and emergent flaviviruses. Curr. Opin. Virol. 2021, 50, 30–39. [Google Scholar] [CrossRef]
- Obi, J.O.; Gutiérrez-Barbosa, H.; Chua, J.V.; Deredge, D.J. Current Trends and Limitations in Dengue Antiviral Research. Trop. Med. Infect. Dis. 2021, 6, 180. [Google Scholar] [CrossRef]
- Mushtaq, M.; Naz, S.; Parang, K.; Ul-Haq, Z. Exploiting Dengue Virus Protease as a Therapeutic Target: Current Status, Challenges and Future Avenues. Curr. Med. Chem. 2021, 28, 7767–7802. [Google Scholar] [CrossRef]
- Troost, B.; Smit, J.M. Recent advances in antiviral drug development towards dengue virus. Curr. Opin. Virol. 2020, 43, 9–21. [Google Scholar] [CrossRef]
- Low, J.G.; Gatsinga, R.; Vasudevan, S.G.; Sampath, A. Dengue Antiviral Development: A Continuing Journey. Adv. Exp. Med. Biol. 2018, 1062, 319–332. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, J.; Kang, S.; Cha, H.J.; Shin, H.; Park, J.; Jang, Y.S.; Woo, J.S.; Won, C.; Min, D.H. Discovery of direct-acting antiviral agents with a graphene-based fluorescent nanosensor. Sci Adv. 2020, 6, eaaz8201. [Google Scholar] [CrossRef]
- Sagaya Jansi, R.; Khusro, A.; Agastian, P.; Alfarhan, A.; Al-Dhabi, N.A.; Arasu, M.V.; Rajagopal, R.; Barcelo, D.; Al-Tamimi, A. Emerging paradigms of viral diseases and paramount role of natural resources as antiviral agents. Sci. Total Environ. 2021, 759, 143539. [Google Scholar] [CrossRef]
- Kausar, M.A.; Ali, A.; Qiblawi, S.; Shahid, S.; Asrar Izhari, M.; Saral, A. Molecular docking based design of Dengue NS5 methyltransferase inhibitors. Bioinformation 2019, 15, 394–401. [Google Scholar] [CrossRef]
- Qamar, M.T.U.; Maryam, A.; Muneer, I.; Xing, F.; Ashfaq, U.A.; Khan, F.A.; Anwar, F.; Geesi, M.H.; Khalid, R.R.; Rauf, S.A.; et al. Computational screening of medicinal plant phytochemicals to discover potent pan-serotype inhibitors against dengue virus. Sci. Rep. 2019, 9, 1433. [Google Scholar] [CrossRef] [Green Version]
- Hariono, M.; Choi, S.B.; Roslim, R.F.; Nawi, M.S.; Tan, M.L.; Kamarulzaman, E.E.; Mohamed, N.; Yusof, R.; Othman, S.; Rahman, N.A.; et al. Thioguanine-based DENV-2 NS2B/NS3 protease inhibitors: Virtual screening, synthesis, biological evaluation and molecular modelling. PLoS ONE 2019, 14, e0210869. [Google Scholar] [CrossRef]
- Yang, C.C.; Hu, H.S.; Lin, H.M.; Wu, P.S.; Wu, R.H.; Tian, J.N.; Wu, S.H.; Tsou, L.K.; Song, J.S.; Chen, H.W.; et al. A novel flavivirus entry inhibitor, BP34610, discovered through high-throughput screening with dengue reporter viruses. Antivir. Res. 2019, 172, 104636. [Google Scholar] [CrossRef]
- Idrus, S.; Tambunan, U.; Zubaidi, A.A. Designing cyclopentapeptide inhibitor as potential antiviral drug for dengue virus ns5 methyltransferase. Bioinformation 2012, 8, 348–352. [Google Scholar] [CrossRef] [Green Version]
- García-Ariza, L.L.; Rocha-Roa, C.; Padilla-Sanabria, L.; Castaño-Osorio, J.C. Virtual Screening of Drug-Like Compounds as Potential Inhibitors of the Dengue Virus NS5 Protein. Front. Chem. 2022, 10, 637266. [Google Scholar] [CrossRef]
- Lim, S.P.; Noble, C.G.; Seh, C.C.; Soh, T.S.; El Sahili, A.; Chan, G.K.; Lescar, J.; Arora, R.; Benson, T.; Nilar, S.; et al. Potent Allosteric Dengue Virus NS5 Polymerase Inhibitors: Mechanism of Action and Resistance Profiling. PLoS Pathog. 2016, 12, e1005737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kufareva, I.; Abagyan, R. Methods of protein structure comparison. Methods Mol. Biol. 2012, 857, 231–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, U.; Tomlinson, S.M.; Fonner, J.M.; Mock, S.A.; Watowich, S.J. Identification of a novel inhibitor of dengue virus protease through use of a virtual screening drug discovery Web portal. J. Chem. Inf. Model. 2014, 54, 2816–2825. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug. Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Diamond, M.S.; Zachariah, M.; Harris, E. Mycophenolic acid inhibits dengue virus infection by preventing replication of viral RNA. Virology 2002, 304, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Kato, F.; Hishiki, T. Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms. Viruses 2016, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Chen, Y.L.; Kondreddi, R.R.; Chan, W.L.; Wang, G.; Ng, R.H.; Lim, J.Y.; Lee, W.Y.; Jeyaraj, D.A.; Niyomrattanakit, P.; et al. N-sulfonylanthranilic acid derivatives as allosteric inhibitors of dengue viral RNA-dependent RNA polymerase. J. Med. Chem. 2009, 52, 7934–7937. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Stanzione, F.; Giangreco, I.; Cole, J.C. Use of molecular docking computational tools in drug discovery. Prog. Med. Chem. 2021, 60, 273–343. [Google Scholar] [CrossRef]
- Gaurav, A.; Agrawal, N.; Al-Nema, M.; Gautam, V. Computational Approaches in the Discovery and Development of Therapeutic and Prophylactic Agents for Viral Diseases. Curr. Top. Med. Chem. 2022, 22, 2190–2206. [Google Scholar] [CrossRef]
- Potisopon, S.; Ferron, F.; Fattorini, V.; Selisko, B.; Canard, B. Substrate selectivity of Dengue and Zika virus NS5 polymerase towards 2′-modified nucleotide analogues. Antivir. Res. 2017, 140, 25–36. [Google Scholar] [CrossRef]
- Carugo, O.; Pongor, S. A normalized root-mean-square distance for comparing protein three-dimensional structures. Protein Sci. 2001, 10, 1470–1473. [Google Scholar] [CrossRef] [Green Version]
- Halder, S.K.; Ahmad, I.; Shathi, J.F.; Mim, M.M.; Hassan, M.R.; Jewel, M.J.I.; Dey, P.; Islam, M.S.; Patel, H.; Morshed, M.R.; et al. A Comprehensive Study to Unleash the Putative Inhibitors of Serotype2 of Dengue Virus: Insights from an In Silico Structure-Based Drug Discovery. Mol. Biotechnol. 2022, 1–14. [Google Scholar] [CrossRef]
- Manish, C.; Ghosh, B.P. Chapter 8—Anti-Tubercular Drug Designing Using Structural Descriptors. In Advances in Mathematical Chemistry and Applications; Subhash, C.B., Restrepo, G., José, L.V., Eds.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2015; pp. 179–190. ISBN 9781681080536. [Google Scholar]
- Hata, H.; Phuoc Tran, D.; Marzouk Sobeh, M.; Kitao, A. Binding free energy of protein/ligand complexes calculated using dissociation Parallel Cascade Selection Molecular Dynamics and Markov state model. Biophys. Physicobiol. 2021, 18, 305–316. [Google Scholar] [CrossRef]
- Fadlan, A.; Nusantoro, Y. The Effect of Energy Minimization on The Molecular Docking of Acetone-Based Oxindole Derivatives. JKPK 2021, 6, 69–77. [Google Scholar] [CrossRef]
- Ahmad, N.; Rehman, A.U.; Badshah, S.L.; Ullah, A.; Mohammad, A.; Khan, K. Molecular dynamics simulation of zika virus NS5 RNA dependent RNA polymerase with selected novel non-nucleoside inhibitors. J. Mol. Struct. 2020, 1203, 127428. [Google Scholar] [CrossRef]
- Dorababu, A. Indole—A promising pharmacophore in recent antiviral drug discovery. RSC Med. Chem. 2020, 11, 1335–1353. [Google Scholar] [CrossRef] [PubMed]
- Norwood, V.M., 4th; Huigens, R.W., 3rd. Harnessing the Chemistry of the Indole Heterocycle to Drive Discoveries in Biology and Medicine. Chembiochem 2019, 20, 2273–2297. [Google Scholar] [CrossRef] [PubMed]
- Sravanthi, T.V.; Manju, S.L. Indoles—A promising scaffold for drug development. Eur. J. Pharm. Sci. 2016, 91, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De, A.; Sarkar, S.; Majee, A. Recent advances on heterocyclic compounds with antiviral properties. Chem. Heterocycl. Compd. 2021, 57, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Giannakopoulou, E.; Pardali, V.; Frakolaki, E.; Siozos, V.; Myrianthopoulos, V.; Mikros, E.; Taylor, M.C.; Kelly, J.M.; Vassilaki, N.; Zoidis, G. Scaffold hybridization strategy towards potent hydroxamate-based inhibitors of Flaviviridae viruses and Trypanosoma species. Medchemcomm 2019, 10, 991–1006. [Google Scholar] [CrossRef] [Green Version]
- Fikatas, A.; Vervaeke, P.; Meyen, E.; Llor, N.; Ordeix, S.; Boonen, I.; Bletsa, M.; Kafetzopoulou, L.E.; Lemey, P.; Amat, M.; et al. A Novel Series of Indole Alkaloid Derivatives Inhibit Dengue and Zika Virus Infection by Interference with the Viral Replication Complex. Antimicrob. Agents Chemother. 2021, 65, e0234920. [Google Scholar] [CrossRef]
- Zhou, G.F.; Li, F.; Xue, J.X.; Qian, W.; Gu, X.R.; Zheng, C.B.; Li, C.; Yang, L.M.; Xiong, S.D.; Zhou, G.C.; et al. Antiviral effects of the fused tricyclic derivatives of indolassaine and imidazolidinone on ZIKV infection and RdRp activities of ZIKV and DENV. Virus Res. 2023, 326, 199062. [Google Scholar] [CrossRef]
- Fang, Y.; Tambo, E.; Xue, J.B.; Zhang, Y.; Zhou, X.N.; Khater, E.I.M. Detection of DENV-2 and Insect-Specific Flaviviruses in Mosquitoes Collected from Jeddah, Saudi Arabia. Front. Cell Infect. Microbiol. 2021, 11, 626368. [Google Scholar] [CrossRef]
- Nath, H.; Basu, K.; De, A.; Biswas, S. Dengue virus sustains viability of infected cells by counteracting apoptosis-mediated DNA breakage. BioRxiv 2020. [Google Scholar] [CrossRef]
- Mello, C.D.S.; Valente, L.M.M.; Wolff, T.; Lima-Junior, R.S.; Fialho, L.G.; Marinho, C.F.; Azeredo, E.L.; Oliveira-Pinto, L.M.; Pereira, R.C.A.; Siani, A.C.; et al. Decrease in Dengue virus-2 infection and reduction of cytokine/chemokine production by Uncaria guianensis in human hepatocyte cell line Huh-7. Mem. Inst. Oswaldo Cruz. 2017, 112, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Songprakhon, P.; Thaingtamtanha, T.; Limjindaporn, T.; Puttikhunt, C.; Srisawat, C.; Luangaram, P.; Dechtawewat, T.; Uthaipibull, C.; Thongsima, S.; Yenchitsomanus, P.T.; et al. Peptides targeting dengue viral nonstructural protein 1 inhibit dengue virus production. Sci. Rep. 2020, 10, 12933. [Google Scholar] [CrossRef]
- Ratanakomol, T.; Roytrakul, S.; Wikan, N.; Smith, D.R. Oroxylin A shows limited antiviral activity towards dengue virus. BMC Res. Notes 2022, 15, 154. [Google Scholar] [CrossRef]
- Kato, F.; Nio, Y.; Yagasaki, K.; Suzuki, R.; Hijikata, M.; Miura, T.; Miyazaki, I.; Tajima, S.; Lim, C.K.; Saijo, M.; et al. Identification of inhibitors of dengue viral replication using replicon cells expressing secretory luciferase. Antivir. Res. 2019, 172, 104643. [Google Scholar] [CrossRef]
- Yao, X.; Ling, Y.; Guo, S.; Wu, W.; He, S.; Zhang, Q.; Zou, M.; Nandakumar, K.S.; Chen, X.; Liu, S. Tatanan A from the Acorus calamus L. root inhibited dengue virus proliferation and infections. Phytomedicine 2018, 42, 258–267. [Google Scholar] [CrossRef]
- Hishiki, T.; Kato, F.; Tajima, S.; Toume, K.; Umezaki, M.; Takasaki, T.; Miura, T.H. An Indole Alkaloid of Uncaria rhynchophylla, Inhibits Late Step in Dengue Virus Lifecycle. Front. Microbiol. 2017, 8, 1674. [Google Scholar] [CrossRef]
- Bardiot, D.; Koukni, M.; Smets, W.; Carlens, G.; McNaughton, M.; Kaptein, S.; Dallmeier, K.; Chaltin, P.; Neyts, J.; Marchand, A. Discovery of Indole Derivatives as Novel and Potent Dengue Virus Inhibitors. J. Med. Chem. 2018, 61, 8390–8401. [Google Scholar] [CrossRef]
- Babbar, R.; Kaur, R.; Rana, P.; Arora, S.; Behl, T.; Albratty, M.; Najmi, A.; Meraya, A.M.; Alhazmi, H.A.; Singla, R.K. The Current Landscape of Bioactive Molecules against DENV: A Systematic Review. Evid. Based Complement. Alternat. Med. 2023, 2023, 2236210. [Google Scholar] [CrossRef]
- Ghasemnezhad, A.; Ghorbanzadeh, A.; Sarmast, M.K.; Ghorbanpour, M. A review on botanical, phytochemical, and pharmacological characteristics of iranian junipers (Juniperus spp.). In Plant-Derived Bioactives; Springer: Singapore, 2020; pp. 493–508. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Site | Zinc Code | Assigned Name | Binding Energy (kcal/mol) | |||
|---|---|---|---|---|---|---|
| DENV-1 | DENV-2 | DENV-2 | DENV-4 | |||
| Mtase Domain | ZINC08790808 | M08 | −11.4 | −11.4 | −11.5 | −11.5 |
| ZINC03839432 | M32 | −11.1 | −11.1 | −11.1 | −11.2 | |
| ZINC08791166 | M66 | −11.4 | −11.5 | −11.5 | −11.4 | |
| ZINC35485176 | M76 | −12.0 | −12.1 | −12.2 | −12.0 | |
| ZINC08790178 | M78 | −12.0 | −12.1 | −12.2 | −12.0 | |
| RdRp Domain | ZINC02094107 | R07 | −9.8 | −9.7 | −9.8 | −9.6 |
| ZINC04085432 | R32 | −10.1 | −10 | −10.5 | −9.9 | |
| ZINC04085246 | R46 | −9.8 | −9.7 | −10.1 | −9.2 | |
| ZINC12884853 | R53 | −9.8 | −9.8 | −10.1 | −9.3 | |
| ZINC20611155 | R55 | −9.8 | −9.6 | −9.8 | −10.0 | |
| Both domain | ZINC08790125 | MR25 | MTase −11.8 | −11.9 | −12.1 | −11.9 |
| RdRp −10.1 | −9.8 | −10.0 | −9.7 | |||
| ZINC08791241 | MR41 | MTase −11.2 | −11.3 | −11.4 | −11.3 | |
| RdRp −10.0 | −9.9 | −10.4 | −9.4 | |||
| ZINC12885588 | MR88 | MTase −11.2 | −11.1 | −11.3 | −11.2 | |
| RdRp −10.4 | −10.3 | −10.8 | −10.2 | |||
| ZINC04086794 | MR94 | MTase −11.1 | −11.2 | −11.4 | −11.1 | |
| RdRp −10.2 | −10.4 | −10.4 | −10.2 | |||
| ZINC08791299 | MR99 | MTase −11.3 | −11.3 | −11.5 | −11.3 | |
| RdRp −10.2 | −10.1 | −10.4 | −9.9 | |||
| Compound | Structure | CC50 (µM) |
|---|---|---|
| M66 |  | 44.08 |
| M76 |  | >100 |
| M78 |  | 60.77 |
| R07 |  | >100 |
| R32 |  | >100 |
| R53 |  | 30 |
| R55 |  | >100 |
| MR25 |  | >100 |
| MR41 |  | >100 |
| MR94 |  | 46.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Ariza, L.L.; González-Rivillas, N.; Díaz-Aguirre, C.J.; Rocha-Roa, C.; Padilla-Sanabria, L.; Castaño-Osorio, J.C. Antiviral Activity of an Indole-Type Compound Derived from Natural Products, Identified by Virtual Screening by Interaction on Dengue Virus NS5 Protein. Viruses 2023, 15, 1563. https://doi.org/10.3390/v15071563
García-Ariza LL, González-Rivillas N, Díaz-Aguirre CJ, Rocha-Roa C, Padilla-Sanabria L, Castaño-Osorio JC. Antiviral Activity of an Indole-Type Compound Derived from Natural Products, Identified by Virtual Screening by Interaction on Dengue Virus NS5 Protein. Viruses. 2023; 15(7):1563. https://doi.org/10.3390/v15071563
Chicago/Turabian StyleGarcía-Ariza, Leidy Lorena, Natalia González-Rivillas, Cindy Johanna Díaz-Aguirre, Cristian Rocha-Roa, Leonardo Padilla-Sanabria, and Jhon Carlos Castaño-Osorio. 2023. "Antiviral Activity of an Indole-Type Compound Derived from Natural Products, Identified by Virtual Screening by Interaction on Dengue Virus NS5 Protein" Viruses 15, no. 7: 1563. https://doi.org/10.3390/v15071563
APA StyleGarcía-Ariza, L. L., González-Rivillas, N., Díaz-Aguirre, C. J., Rocha-Roa, C., Padilla-Sanabria, L., & Castaño-Osorio, J. C. (2023). Antiviral Activity of an Indole-Type Compound Derived from Natural Products, Identified by Virtual Screening by Interaction on Dengue Virus NS5 Protein. Viruses, 15(7), 1563. https://doi.org/10.3390/v15071563