Antiviral Strategies for Pandemic and Seasonal Influenza

Abstract
:1. Introduction
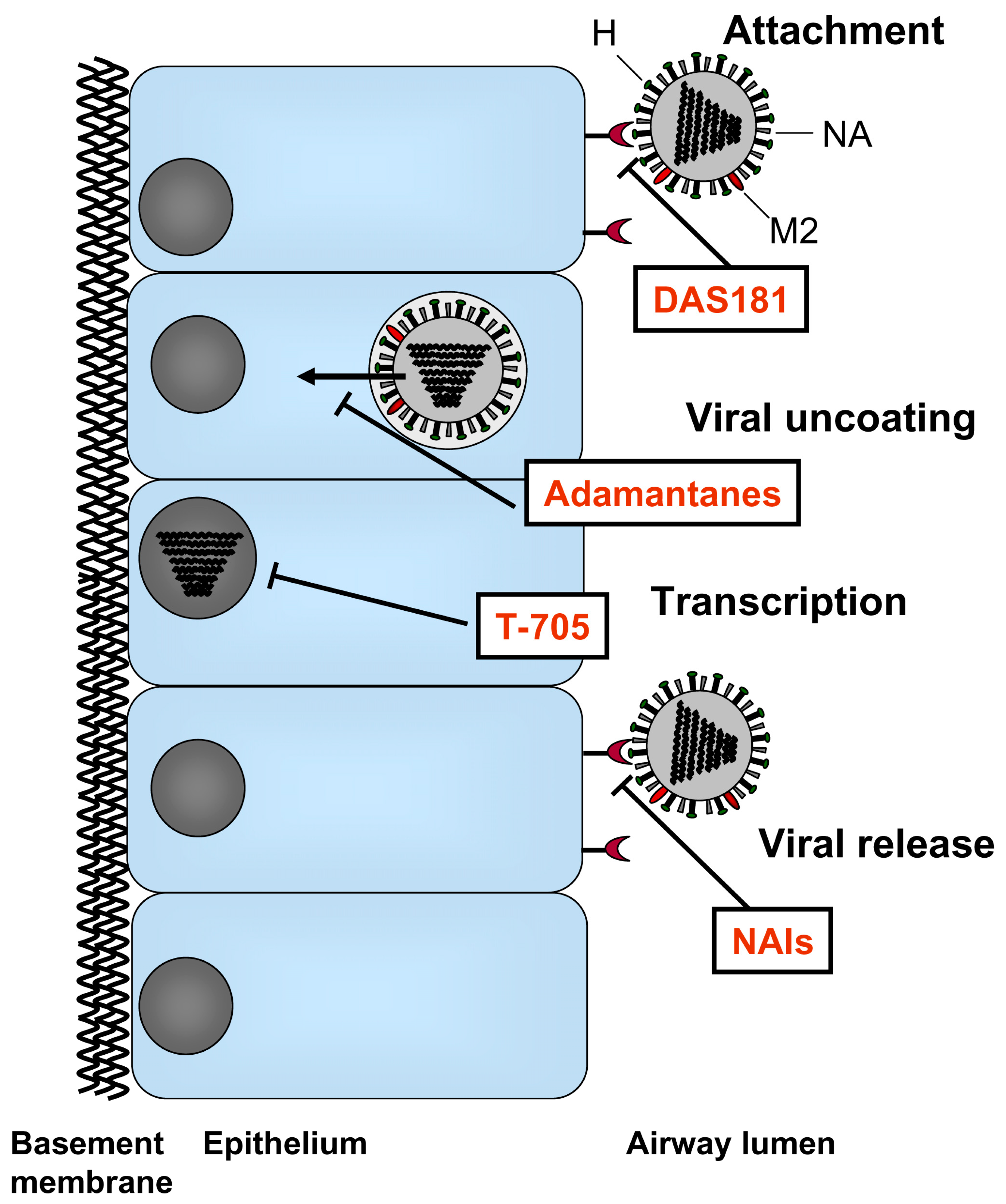
2. Currently Available Therapeutics
2.1. M2 Ion Channel Blockers
2.2. NAIs
3. New Antiviral Agents for Influenza Treatment
4. Pathogen-Targeted Approaches
4.1. NA Inhibitors
4.2. Polymerase Inhibitors
5. Host-Targeted Approaches
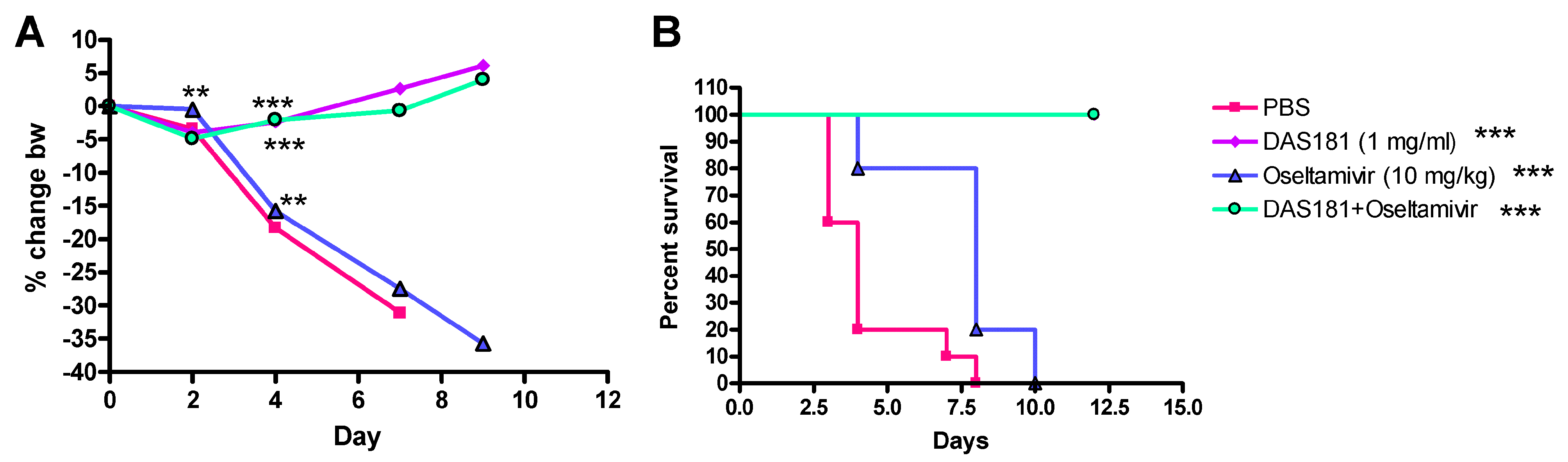
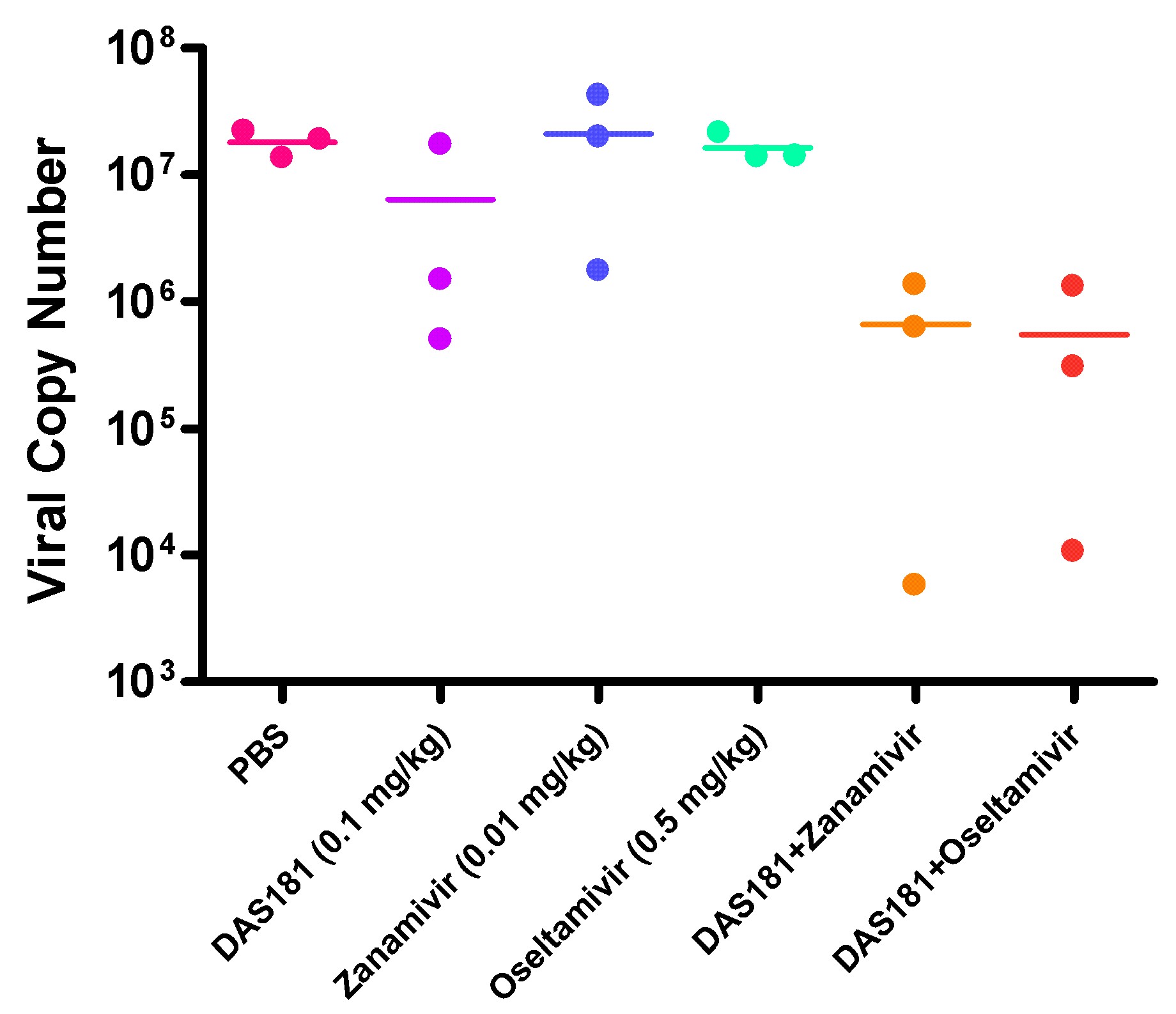
6. Combination Therapy
7. Conclusions
Acknowledgments
References and Notes
- Smith, N.M.; Bresee, J.S.; Shay, D.K.; Uyeki, T.M.; Cox, N.J.; Strikas, R.A. Prevention and Control of Influenza: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2006, 55, 1–42. [Google Scholar]
- Weinstock, D.M.; Zuccotti, G. The evolution of influenza resistance and treatment. JAMA 2009, 301, 1066–1069. [Google Scholar] [CrossRef]
- Moscona, A. Oseltamivir resistance—Disabling our influenza defenses. N. Engl. J. Med. 2005, 353, 2633–2636. [Google Scholar] [CrossRef]
- Zimmer, S.M.; Burke, D.S. Historical perspective—Emergence of influenza A (H1N1) viruses. N. Engl. J. Med. 2009, 361, 279–285. [Google Scholar] [CrossRef]
- Jefferson, T.; Demicheli, V.; Di, P.C.; Rivetti, D. Amantadine and rimantadine for influenza A in adults. Cochrane Database Syst. Rev. 2006, CD001169. [Google Scholar] [CrossRef]
- Suzuki, H.; Saito, R.; Masuda, H.; Oshitani, H.; Sato, M.; Sato, I. Emergence of amantadine-resistant influenza A viruses: Epidemiological study. J. Infect. Chemother. 2003, 9, 195–200. [Google Scholar] [CrossRef]
- Shinde, V.; Bridges, C.B.; Uyeki, T.M.; Shu, B.; Balish, A.; Xu, X.; Lindstrom, S.; Gubareva, L.V.; Deyde, V.; Garten, R.J.; et al. Triple-Reassortant Swine Influenza A (H1) in Humans in the United States, 2005–2009. N. Engl. J. Med. 2009, 360, 2616–2625. [Google Scholar] [CrossRef]
- Dawood, F.S.; Jain, S.; Finelli, L.; Shaw, M.W.; Lindstrom, S.; Garten, R.J.; Gubareva, L.V.; Xu, X.; Bridges, C.B.; Uyeki, T.M. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N. Engl. J. Med. 2009, 360, 2605–2615. [Google Scholar]
- Rungrotmongkol, T.; Intharathep, P.; Malaisree, M.; Nunthaboot, N.; Kaiyawet, N.; Sompornpisut, P.; Payungporn, S.; Poovorawan, Y.; Hannongbua, S. Susceptibility of antiviral drugs against 2009 influenza A (H1N1) virus. Biochem. Biophys. Res. Commun. 2009, 385, 390–394. [Google Scholar] [CrossRef]
- Glaxo SmithKline Inc. Relenza (Zanamivir) Inhalation Powder, 2010. Available online: http://us.gsk.com/products/assets/us_relenza.pdf (accessed on 18 August 2010).
- Roche Laboratories Inc. Tamiflu (Oseltamivir Phosphate) Capsules and for Oral Suspension, 2008. Available online: http://www.accessdata.fda.gov/drugsatfda_docs/label/2008/021087s047,%20021246s033lbl.pdf (accessed on 18 August 2010).
- Cooper, N.J.; Sutton, A.J.; Abrams, K.R.; Wailoo, A.; Turner, D.; Nicholson, K.G. Effectiveness of neuraminidase inhibitors in treatment and prevention of influenza A and B: Systematic review and meta-analyses of randomised controlled trials. BMJ 2003, 326, 1235–1242. [Google Scholar] [CrossRef]
- McGeer, A.; Green, K.A.; Plevneshi, A.; Shigayeva, A.; Siddiqi, N.; Raboud, J.; Low, D.E. Antiviral therapy and outcomes of influenza requiring hospitalization in Ontario, Canada. Clin. Infect. Dis. 2007, 45, 1568–1575. [Google Scholar] [CrossRef]
- Mishin, V.P.; Hayden, F.G.; Gubareva, L.V. Susceptibilities of antiviral-resistant influenza viruses to novel neuraminidase inhibitors. Antimicrob. Agents Chemother. 2005, 49, 4515–4520. [Google Scholar] [CrossRef]
- Dharan, N.J.; Gubareva, L.V.; Meyer, J.J.; Okomo-Adhiambo, M.; McClinton, R.C.; Marshall, S.A.; St, G.K.; Epperson, S.; Brammer, L.; Klimov, A.I.; et al. Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA 2009, 301, 1034–1041. [Google Scholar] [CrossRef]
- Oseltamivir-resistant 2009 pandemic influenza A (H1N1) virus infection in two summer campers receiving prophylaxis--North Carolina, 2009. MMWR Morb. Mortal. Wkly. Rep. 2009, 58, 969–972.
- Leung, T.W.; Tai, A.L.; Cheng, P.K.; Kong, M.S.; Lim, W. Detection of an oseltamivir-resistant pandemic influenza A/H1N1 virus in Hong Kong. J. Clin. Virol. 2009, 46, 298–299. [Google Scholar] [CrossRef]
- WHO: Oseltamivir Resistance in Immunocompromised Hospital Patients. WHO. Available online: http://www.who.int/csr/disease/swineflu/notes/briefing_20091202/en/index.html (accessed on 13 August 2010).
- WHO: Viruses Resistant to Oseltamivir (Tamiflu) Identified. WHO. Available online: http://www.who.int/csr/disease/swineflu/notes/h1n1_antiviral_resistance_20090708/en/index.html (accessed on 13 August 2010).
- CDC: Seasonal Influenza: 2008–2009 Influenza Season Week 39. CDC. Available online: http://www.cdc.gov/flu/weekly/weeklyarchives2008-2009/weekly39.htm (accessed on 13 August 2010).
- Wetherall, N.T.; Trivedi, T.; Zeller, J.; Hodges-Savola, C.; McKimm-Breschkin, J.L.; Zambon, M.; Hayden, F.G. Evaluation of neuraminidase enzyme assays using different substrates to measure susceptibility of influenza virus clinical isolates to neuraminidase inhibitors: Report of the neuraminidase inhibitor susceptibility network. J. Clin. Microbiol. 2003, 41, 742–750. [Google Scholar] [CrossRef]
- Triana-Baltzer, G.B.; Gubareva, L.V.; Klimov, A.I.; Wurtman, D.F.; Moss, R.B.; Hedlund, M.; Larson, J.L.; Belshe, R.B.; Fang, F. Inhibition of neuraminidase inhibitor-resistant influenza virus by DAS181, a novel sialidase fusion protein. PLoS ONE 2009, 4, e7838. [Google Scholar] [CrossRef]
- Lackenby, A.; Thompson, C.I.; Democratis, J. The potential impact of neuraminidase inhibitor resistant influenza. Curr. Opin. Infect. Dis. 2008, 21, 626–638. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Matrosovich, M.N.; Brenner, M.K.; Bethell, R.C.; Webster, R.G. Evidence for zanamivir resistance in an immunocompromised child infected with influenza B virus. J. Infect. Dis. 1998, 178, 1257–1262. [Google Scholar] [CrossRef]
- Beigel, J.; Bray, M. Current and future antiviral therapy of severe seasonal and avian influenza. Antiviral Res. 2008, 78, 91–102. [Google Scholar] [CrossRef]
- Boltz, D.A.; Aldridge, J.R., Jr.; Webster, R.G.; Govorkova, E.A. Drugs in development for influenza. Drugs 2010, 70, 1349–1362. [Google Scholar] [CrossRef]
- De, C.E. Antiviral agents active against influenza A viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar]
- Hsieh, H.P.; Hsu, J.T. Strategies of development of antiviral agents directed against influenza virus replication. Curr. Pharm Des. 2007, 13, 3531–3542. [Google Scholar] [CrossRef]
- Cass, L.M.; Efthymiopoulos, C.; Bye, A. Pharmacokinetics of zanamivir after intravenous, oral, inhaled or intranasal administration to healthy volunteers. Clin. Pharmacokinet. 1999, 36 (Suppl. 1), 1–11. [Google Scholar] [CrossRef]
- Calfee, D.P.; Peng, A.W.; Cass, L.M.; Lobo, M.; Hayden, F.G. Safety and efficacy of intravenous zanamivir in preventing experimental human influenza A virus infection. Antimicrob. Agents Chemother. 1999, 43, 1616–1620. [Google Scholar] [CrossRef]
- Stittelaar, K.J.; Tisdale, M.; van, A.G.; van Lavieren, R.F.; Pistoor, F.; Simon, J.; Osterhaus, A.D. Evaluation of intravenous zanamivir against experimental influenza A (H5N1) virus infection in cynomolgus macaques. Antiviral Res. 2008, 80, 225–228. [Google Scholar] [CrossRef]
- Higgs, E.S.; Hayden, F.G.; Chotpitayasunondh, T.; Whitworth, J.; Farrar, J. The Southeast Asian Influenza Clinical Research Network: Development and challenges for a new multilateral research endeavor. Antiviral Res. 2008, 78, 64–68. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Webster, R.G.; Hayden, F.G. Comparison of the activities of zanamivir, oseltamivir, and RWJ-270201 against clinical isolates of influenza virus and neuraminidase inhibitor-resistant variants. Antimicrob. Agents Chemother. 2001, 45, 3403–3408. [Google Scholar] [CrossRef]
- Bantia, S.; Arnold, C.S.; Parker, C.D.; Upshaw, R.; Chand, P. Anti-influenza virus activity of peramivir in mice with single intramuscular injection. Antiviral Res. 2006, 69, 39–45. [Google Scholar] [CrossRef]
- Barroso, L.; Treanor, J.; Gubareva, L.; Hayden, F.G. Efficacy and tolerability of the oral neuraminidase inhibitor peramivir in experimental human influenza: Randomized, controlled trials for prophylaxis and treatment. Antivir. Ther. 2005, 10, 901–910. [Google Scholar] [CrossRef]
- Kilpatrick, J.M.; Harman, L.A.; Collis, P.J.; Aitee, G.; Mead, E.; Alexander, W.J. Pharmacokinetics and safety of peramivir by intramuscular administration. In Program and abstracts of the Options for the Control of Influenza Conference VI Conference. In Proceedings of the London International Society for Influenza and Other Respiratory Virus Diseases, Toronto, Canada, June 17–23 2007; Abstract Number P916.
- BioCryst Pharmaceuticals I.: BioCryst Provides Peramivir Update and Reports First Quarter 2009 Financial Results. BioCryst Pharmaceuticals, Inc. May 8, 2009. Available online: http://investor.shareholder.com/biocryst/releasedetail.cfm?ReleaseID=382718 (accessed on 12 August 2010).
- BioCryst Pharmaceuticals I.: BioCryst Reports Preliminary Results From a Phase II Clinical Trial of Peramivir in Subjects With Acute Influenza. BioCryst Pharmaceuticals, Inc. September 19, 2007. Available online: http://investor.shareholder.com/biocryst/releasedetail.cfm?ReleaseID=264815 (accessed on 12 August 2010).
- BioCryst Pharmaceuticals I.: Data From Phase 2 Study of Peramivir in Patients Hospitalized With Influenza Presented at the XI International Symposium on Respiratory Viral Infections. BioCryst Pharmaceuticals, Inc. February 23, 2009. Available online: http://investor.shareholder.com/biocryst/releasedetail.cfm?ReleaseID=366984 (accessed on 12 August 2010).
- Hurt, A.C.; Ernest, J.; Deng, Y.M.; Iannello, P.; Besselaar, T.G.; Birch, C.; Buchy, P.; Chittaganpitch, M.; Chiu, S.C.; Dwyer, D.; et al. Emergence and spread of oseltamivir-resistant A(H1N1) influenza viruses in Oceania, South East Asia and South Africa. Antiviral Res. 2009, 83, 90–93. [Google Scholar] [CrossRef]
- Birnkrant, D.; Cox, E. The Emergency Use Authorization of peramivir for treatment of 2009 H1N1 influenza. N. Engl. J. Med. 2009, 361, 2204–2207. [Google Scholar] [CrossRef]
- Yamashita, M.; Tomozawa, T.; Kakuta, M.; Tokumitsu, A.; Nasu, H.; Kubo, S. CS-8958, a prodrug of the new neuraminidase inhibitor R-125489, shows long-acting anti-influenza virus activity. Antimicrob. Agents Chemother. 2009, 53, 3756–3764. [Google Scholar] [CrossRef]
- Biota: LANI Phase III Clinical Trials in Asia Prove Successful. Biota Press Release. August 10, 2009. Available online: http://www.biota.com.au/uploaded/154/1021542_25laniphaseiiiclinicaltri.pdf (accessed on 12 August 2010).
- Furuta, Y.; Takahashi, K.; Kuno-Maekawa, M.; Sangawa, H.; Uehara, S.; Kozaki, K.; Nomura, N.; Egawa, H.; Shiraki, K. Mechanism of action of T-705 against influenza virus. Antimicrob. Agents Chemother. 2005, 49, 981–986. [Google Scholar] [CrossRef]
- Smee, D.F.; Hurst, B.L.; Egawa, H.; Takahashi, K.; Kadota, T.; Furuta, Y. Intracellular metabolism of favipiravir (T-705) in uninfected and influenza A (H5N1) virus-infected cells. J. Antimicrob. Chemother. 2009, 64, 741–746. [Google Scholar] [CrossRef]
- Furuta, Y.; Takahashi, K.; Shiraki, K.; Sakamoto, K.; Smee, D.F.; Barnard, D.L.; Gowen, B.B.; Julander, J.G.; Morrey, J.D. T-705 (favipiravir) and related compounds: Novel broad-spectrum inhibitors of RNA viral infections. Antiviral Res. 2009, 82, 95–102. [Google Scholar] [CrossRef]
- Kiso, M.; Takahashi, K.; Sakai-Tagawa, Y.; Shinya, K.; Sakabe, S.; Le, Q.M.; Ozawa, M.; Furuta, Y.; Kawaoka, Y. T-705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 882–887. [Google Scholar] [CrossRef]
- Itoh, Y.; Shinya, K.; Kiso, M.; Watanabe, T.; Sakoda, Y.; Hatta, M.; Muramoto, Y.; Tamura, D.; Sakai-Tagawa, Y.; Noda, T.; et al. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature 2009, 460, 1021–1025. [Google Scholar] [CrossRef]
- Gowen, B.B.; Smee, D.F.; Wong, M.H.; Hall, J.O.; Jung, K.H.; Bailey, K.W.; Stevens, J.R.; Furuta, Y.; Morrey, J.D. Treatment of late stage disease in a model of arenaviral hemorrhagic fever: T-705 efficacy and reduced toxicity suggests an alternative to ribavirin. PLoS ONE 2008, 3, e3725. [Google Scholar] [CrossRef]
- Morrey, J.D.; Taro, B.S.; Siddharthan, V.; Wang, H.; Smee, D.F.; Christensen, A.J.; Furuta, Y. Efficacy of orally administered T-705 pyrazine analog on lethal West Nile virus infection in rodents. Antiviral Res. 2008, 80, 377–379. [Google Scholar] [CrossRef]
- Julander, J.G.; Shafer, K.; Smee, D.F.; Morrey, J.D.; Furuta, Y. Activity of T-705 in a hamster model of yellow fever virus infection in comparison with that of a chemically related compound, T-1106. Antimicrob. Agents Chemother. 2009, 53, 202–209. [Google Scholar] [CrossRef]
- Malakhov, M.P.; Aschenbrenner, L.M.; Smee, D.F.; Wandersee, M.K.; Sidwell, R.W.; Gubareva, L.V.; Mishin, V.P.; Hayden, F.G.; Kim, d.H.; Ing, A.; et al. Sialidase fusion protein as a novel broad-spectrum inhibitor of influenza virus infection. Antimicrob. Agents Chemother. 2006, 50, 1470–1479. [Google Scholar] [CrossRef]
- Ah-Tye, C.; Schwartz, S.; Huberman, K.; Carlin, E.; Moscona, A. Virus-receptor interactions of human parainfluenza viruses types 1, 2 and 3. Microb. Pathog. 1999, 27, 329–336. [Google Scholar] [CrossRef]
- Belser, J.A.; Lu, X.; Szretter, K.J.; Jin, X.; Aschenbrenner, L.M.; Lee, A.; Hawley, S.; Kim, d.H.; Malakhov, M.P.; Yu, M.; et al. DAS181, a novel sialidase fusion protein, protects mice from lethal avian influenza H5N1 virus infection. J. Infect. Dis. 2007, 196, 1493–1499. [Google Scholar] [CrossRef]
- Nicholls, J.M.; Aschenbrenner, L.M.; Paulson, J.C.; Campbell, E.R.; Malakhov, M.P.; Wurtman, D.F.; Yu, M.; Fang, F. Comment on: Concerns of using sialidase fusion protein as an experimental drug to combat seasonal and pandemic influenza. J. Antimicrob. Chemother. 2008, 62, 426–428. [Google Scholar] [CrossRef]
- Triana-Baltzer, G.B.; Babizki, M.; Chan, M.C.; Wong, A.C.; Aschenbrenner, L.M.; Campbell, E.R.; Li, Q.X.; Chan, R.W.; Peiris, J.S.; Nicholls, J.M.; et al. DAS181, a sialidase fusion protein, protects human airway epithelium against influenza virus infection: An in vitro pharmacodynamic analysis. J. Antimicrob. Chemother. 2010, 65, 275–284. [Google Scholar] [CrossRef]
- Chan, R.W.; Chan, M.C.; Wong, A.C.; Karamanska, R.; Dell, A.; Haslam, S.M.; Sihoe, A.D.; Chui, W.H.; Triana-Baltzer, G.; Li, Q.; et al. DAS181 inhibits H5N1 influenza virus infection of human lung tissues. Antimicrob. Agents Chemother. 2009, 53, 3935–3941. [Google Scholar] [CrossRef]
- Triana-Baltzer, G.B.; Gubareva, L.V.; Nicholls, J.M.; Pearce, M.B.; Mishin, V.P.; Belser, J.A.; Chen, L.M.; Chan, R.W.; Chan, M.C.; Hedlund, M.; et al. Novel pandemic influenza A(H1N1) viruses are potently inhibited by DAS181, a sialidase fusion protein. PLoS ONE 2009, 4, e7788. [Google Scholar] [CrossRef]
- Triana-Baltzer, G.B.; Gubareva, L.; Nicholls, J.M.; Pearce, M.B.; Mishin, V.P.; Belser, J.A.; Chen, L.M.; Chan, R.W.Y.; Chan, M.C.W.; Hedlund, M.; et al. Novel Swine-Origin A(H1N1) Influenza Viruses are Potently Inhibited by DAS181, a Sialidase Fusion Protein. PLoS ONE 2009, 4, e7788. [Google Scholar]
- Bantia, S.; Ghate, A.A.; Ananth, S.L.; Babu, Y.S.; Air, G.M.; Walsh, G.M. Generation and characterization of a mutant of influenza A virus selected with the neuraminidase inhibitor BCX-140. Antimicrob. Agents Chemother. 1998, 42, 801–807. [Google Scholar] [CrossRef]
- Barnett, J.M.; Cadman, A.; Burrell, F.M.; Madar, S.H.; Lewis, A.P.; Tisdale, M.; Bethell, R. In vitro selection and characterisation of influenza B/Beijing/1/87 isolates with altered susceptibility to zanamivir. Virology 1999, 265, 286–295. [Google Scholar] [CrossRef]
- Baum, E.Z.; Wagaman, P.C.; Ly, L.; Turchi, I.; Le, J.; Bucher, D.; Bush, K. A point mutation in influenza B neuraminidase confers resistance to peramivir and loss of slow binding. Antiviral Res. 2003, 59, 13–22. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Bethell, R.; Hart, G.J.; Murti, K.G.; Penn, C.R.; Webster, R.G. Characterization of mutants of influenza A virus selected with the neuraminidase inhibitor 4-guanidino-Neu5Ac2en. J. Virol. 1996, 70, 1818–1827. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Robinson, M.J.; Bethell, R.C.; Webster, R.G. Catalytic and framework mutations in the neuraminidase active site of influenza viruses that are resistant to 4-guanidino-Neu5Ac2en. J. Virol. 1997, 71, 3385–3390. [Google Scholar] [CrossRef]
- McKimm-Breschkin, J.L.; McDonald, M.; Blick, T.J.; Colman, P.M. Mutation in the influenza virus neuraminidase gene resulting in decreased sensitivity to the neuraminidase inhibitor 4-guanidino-Neu5Ac2en leads to instability of the enzyme. Virology 1996, 225, 240–242. [Google Scholar] [CrossRef]
- McKimm-Breschkin, J.L.; Blick, T.J.; Sahasrabudhe, A.; Tiong, T.; Marshall, D.; Hart, G.J.; Bethell, R.C.; Penn, C.R. Generation and characterization of variants of NWS/G70C influenza virus after in vitro passage in 4-amino-Neu5Ac2en and 4-guanidino-Neu5Ac2en. Antimicrob. Agents Chemother. 1996, 40, 40–46. [Google Scholar] [CrossRef]
- McKimm-Breschkin, J.L.; Sahasrabudhe, A.; Blick, T.J.; McDonald, M.; Colman, P.M.; Hart, G.J.; Bethell, R.C.; Varghese, J.N. Mutations in a conserved residue in the influenza virus neuraminidase active site decreases sensitivity to Neu5Ac2en-derived inhibitors. J. Virol. 1998, 72, 2456–2462. [Google Scholar] [CrossRef]
- McKimm-Breschkin, J.L. Resistance of influenza viruses to neuraminidase inhibitors—A review. Antiviral Res. 2000, 47, 1–17. [Google Scholar] [CrossRef]
- Molla, A.; Kati, W.; Carrick, R.; Steffy, K.; Shi, Y.; Montgomery, D.; Gusick, N.; Stoll, V.S.; Stewart, K.D.; Ng, T.I.; et al. In vitro selection and characterization of influenza A (A/N9) virus variants resistant to a novel neuraminidase inhibitor, A-315675. J. Virol. 2002, 76, 5380–5386. [Google Scholar] [CrossRef]
- Smee, D.F.; Sidwell, R.W.; Morrison, A.C.; Bailey, K.W.; Baum, E.Z.; Ly, L.; Wagaman, P.C. Characterization of an influenza A (H3N2) virus resistant to the cyclopentane neuraminidase inhibitor RWJ-270201. Antiviral Res. 2001, 52, 251–259. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Bethell, R.; Hart, G.J.; Murti, K.G.; Penn, C.R.; Webster, R.G. Characterization of mutants of influenza A virus selected with the neuraminidase inhibitor 4-guanidino-Neu5Ac2en. J. Virol. 1996, 70, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, M.; Aschenbrenner, L.M.; Jensen, K.; Larson, J.L.; Fang, F. Sialidase-based anti-influenza virus therapy protects against secondary pneumococcal infection. J. Infect. Dis. 2010, 201, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, D. The Evolving Role of Antivirals. Presented at the Influenza Congress USA 2009, Washington, DC, USA, November 18–20, 2009. [Google Scholar]
- Moss, R.B.; Davey, R.T.; Steigbigel, R.T.; Fang, F. Targeting pandemic influenza: A primer on influenza antivirals and drug resistance. J. Antimicrob. Chemother. 2010, 65, 1086–1093. [Google Scholar] [CrossRef]
- Hayden, F. Developing new antiviral agents for influenza treatment: What does the future hold? Clin. Infect. Dis. 2009, 48 (Suppl. 1), S3–S13. [Google Scholar] [CrossRef]
- Ilyushina, N.A.; Bovin, N.V.; Webster, R.G.; Govorkova, E.A. Combination chemotherapy, a potential strategy for reducing the emergence of drug-resistant influenza A variants. Antiviral Res. 2006, 70, 121–131. [Google Scholar] [CrossRef]
- Ison, M.G.; Gnann, J.W., Jr.; Nagy-Agren, S.; Treannor, J.; Paya, C.; Steigbigel, R.; Elliott, M.; Weiss, H.L.; Hayden, F.G. Safety and efficacy of nebulized zanamivir in hospitalized patients with serious influenza. Antivir. Ther. 2003, 8, 183–190. [Google Scholar] [PubMed]
- Ilyushina, N.A.; Hay, A.; Yilmaz, N.; Boon, A.C.; Webster, R.G.; Govorkova, E.A. Oseltamivir-ribavirin combination therapy for highly pathogenic H5N1 influenza virus infection in mice. Antimicrob. Agents Chemother. 2008, 52, 3889–3897. [Google Scholar] [CrossRef]
- Smee, D.F.; Hurst, B.L.; Wong, M.H.; Bailey, K.W.; Tarbet, E.B.; Morrey, J.D.; Furuta, Y. Effects of the combination of favipiravir (T-705) and oseltamivir on influenza A virus infections in mice. Antimicrob. Agents Chemother. 2010, 54, 126–133. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Candidates | Mechanism of Action | Route of Delivery | Phase of Development | Developed By |
| Long-acting NAI (CS-8958) | Inhibition of viral release | Inhaled | Phase III (Japan) clinical trial | Biota/Daiichi-Sankyo |
| Peramivir | Inhibition of viral release | i.v | Phase III clinical trial* | BioCryst Pharmaceuticals |
| T-705 (favipiravir) | Inhibition of RNA replication | Oral | Phase II clinical trial | Toyama Chemical Co., Ltd (a subsidiary of Fuji Holdings) |
| DAS181 (Fludase®) | Inhibition of host cell receptor binding | Inhaled | Phase II clinical trial | NexBio Inc. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2010 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedlund, M.; Larson, J.L.; Fang, F. Antiviral Strategies for Pandemic and Seasonal Influenza. Viruses 2010, 2, 1766-1781. https://doi.org/10.3390/v2081766
Hedlund M, Larson JL, Fang F. Antiviral Strategies for Pandemic and Seasonal Influenza. Viruses. 2010; 2(8):1766-1781. https://doi.org/10.3390/v2081766
Chicago/Turabian StyleHedlund, Maria, Jeffrey L. Larson, and Fang Fang. 2010. "Antiviral Strategies for Pandemic and Seasonal Influenza" Viruses 2, no. 8: 1766-1781. https://doi.org/10.3390/v2081766
APA StyleHedlund, M., Larson, J. L., & Fang, F. (2010). Antiviral Strategies for Pandemic and Seasonal Influenza. Viruses, 2(8), 1766-1781. https://doi.org/10.3390/v2081766