Mechanisms and Factors that Influence High Frequency Retroviral Recombination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
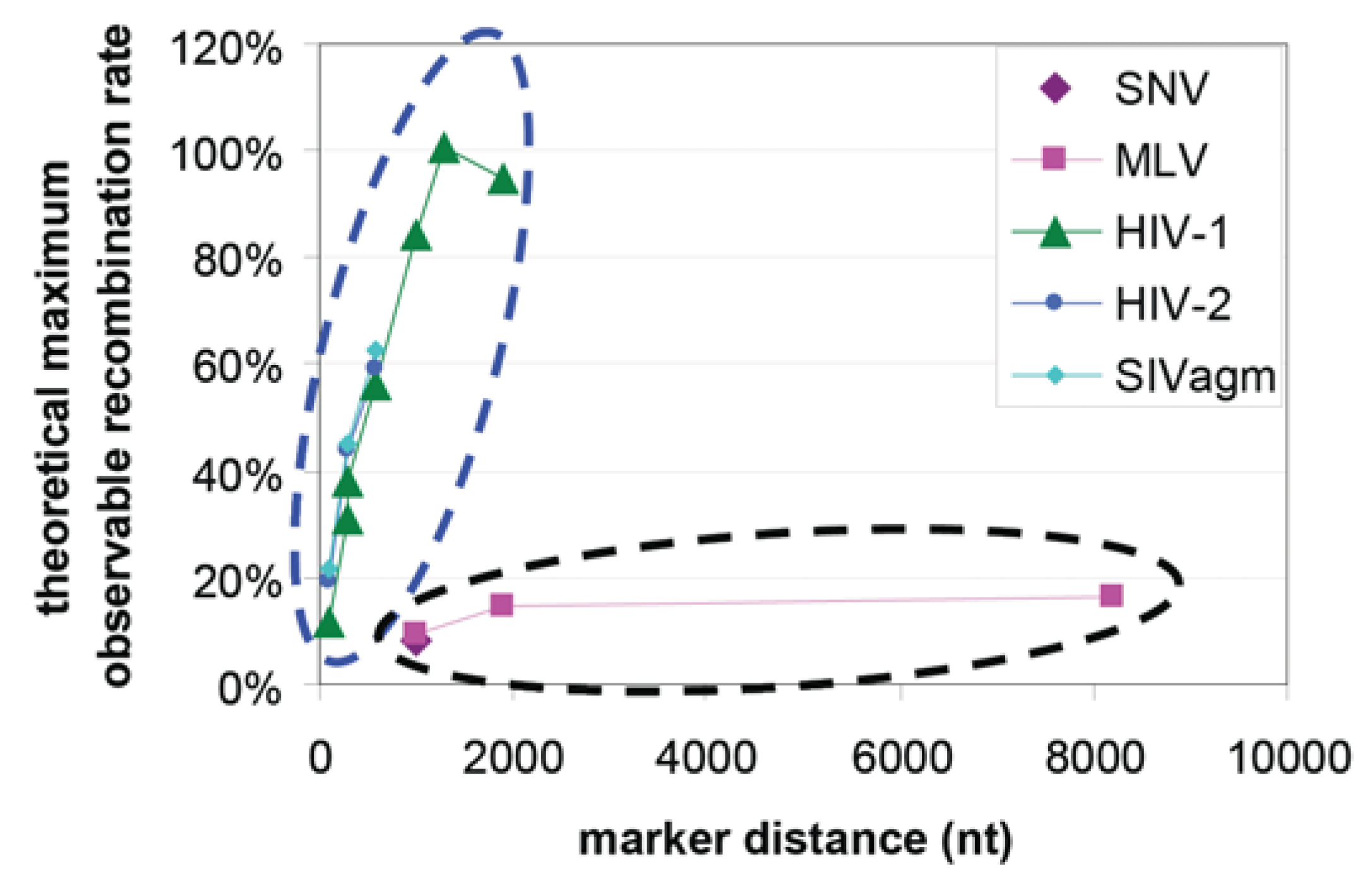
:1. Introduction
1.1. Retroviral Replication Cycle
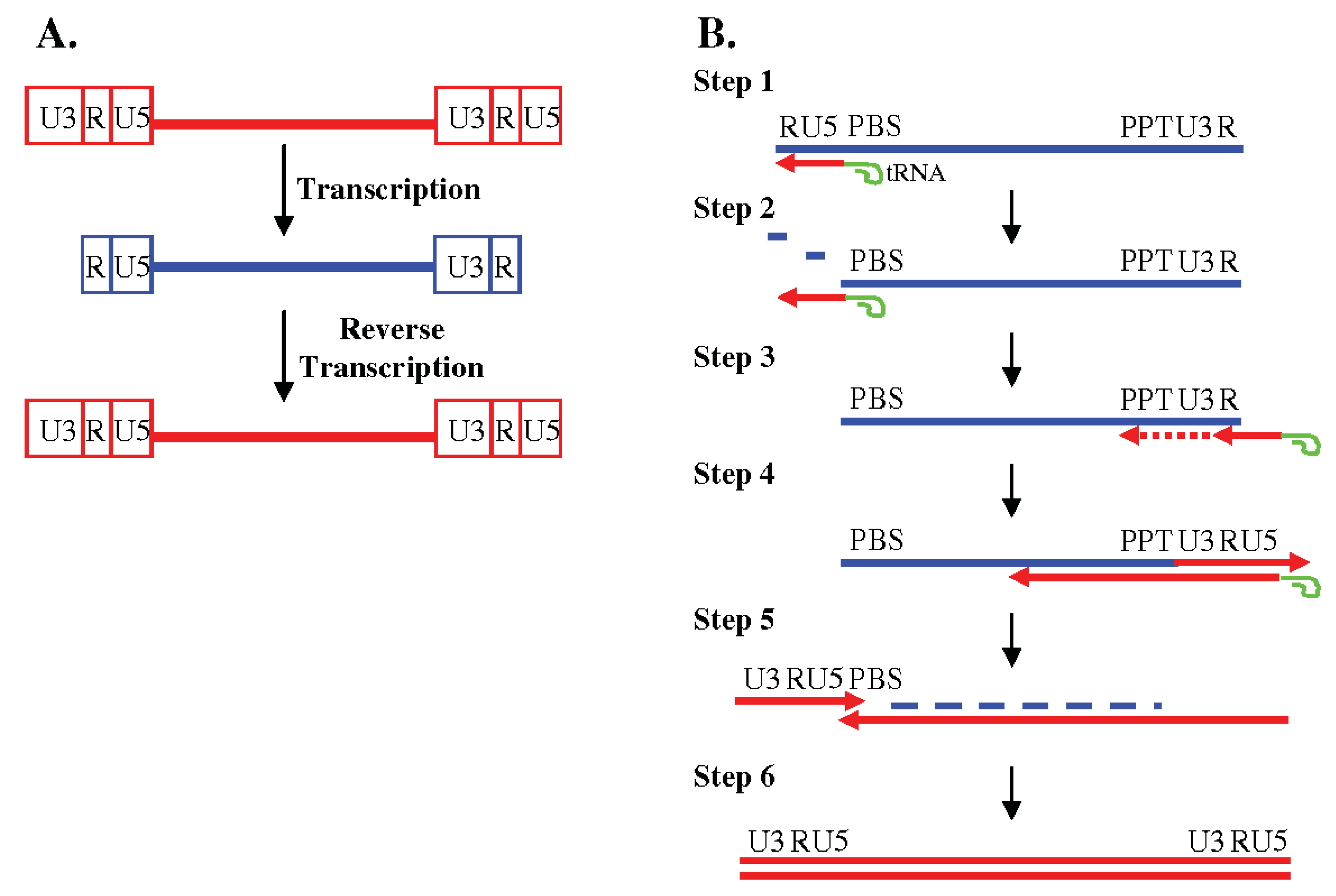
1.2. Reverse Transcription of the Viral RNA into DNA
1.3. Retroviral Recombination Models
1.4. Requirements for the Generation of Observable Recombination Events
2. Template Switching During Reverse Transcription
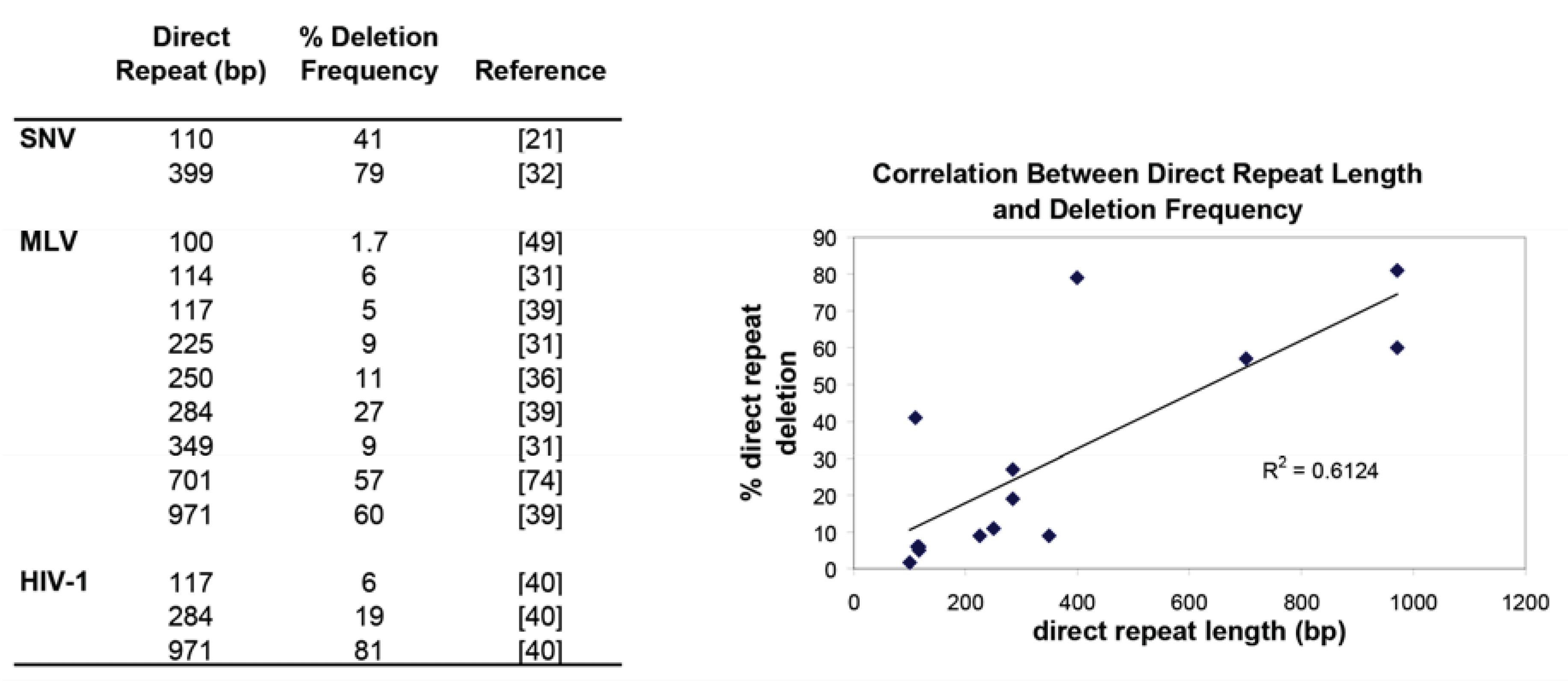
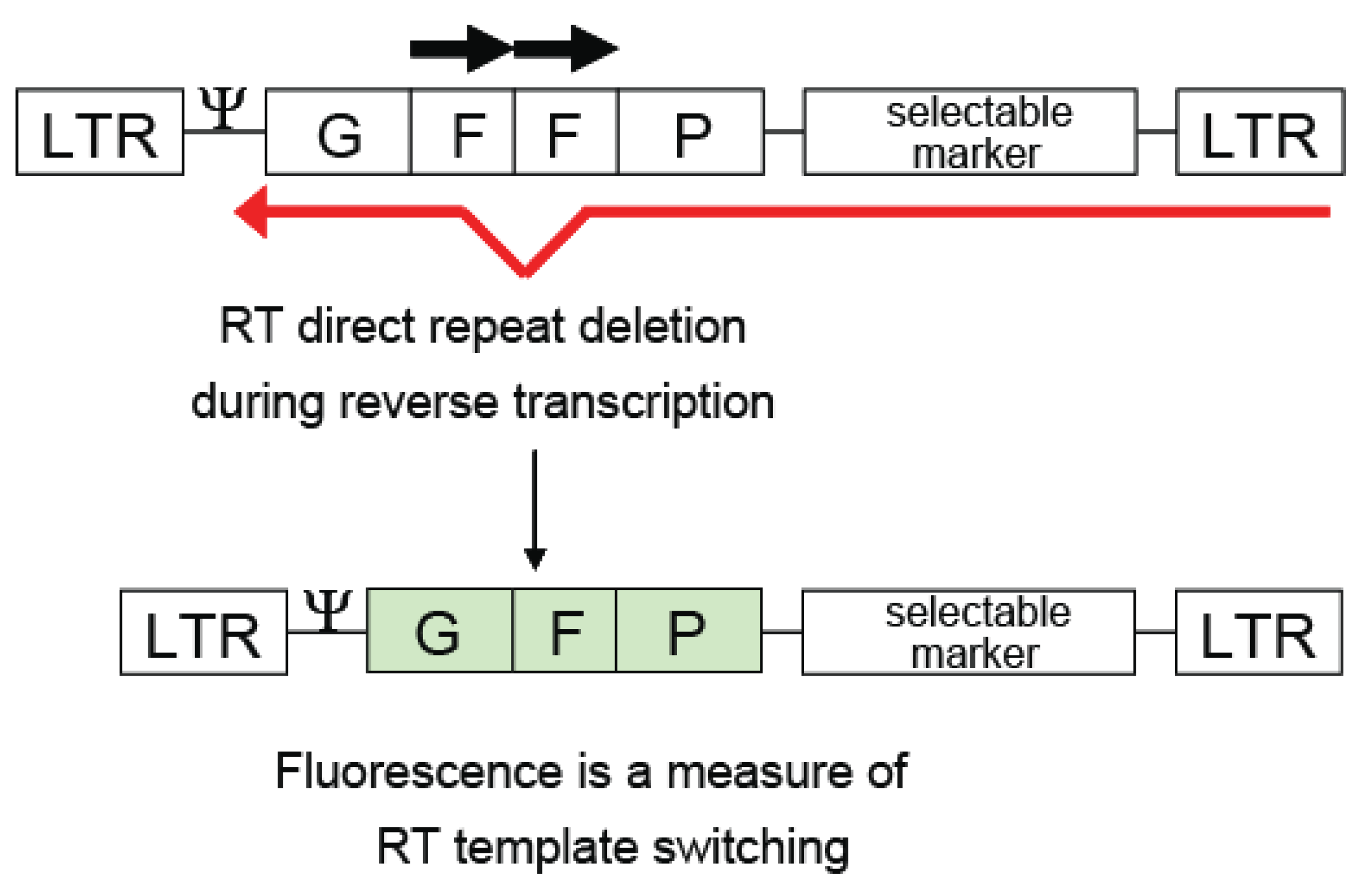
2.1. In Vivo Direct Repeat Deletion Reveals that Template Switching Occurs Frequently During Reverse Transcription and is Dependent Upon the Size and Distance Between the Direct Repeats
2.2. Frequency of Direct Repeat Deletion in Vivo Is Influenced by Nucleotide Pools and Mutations in RT
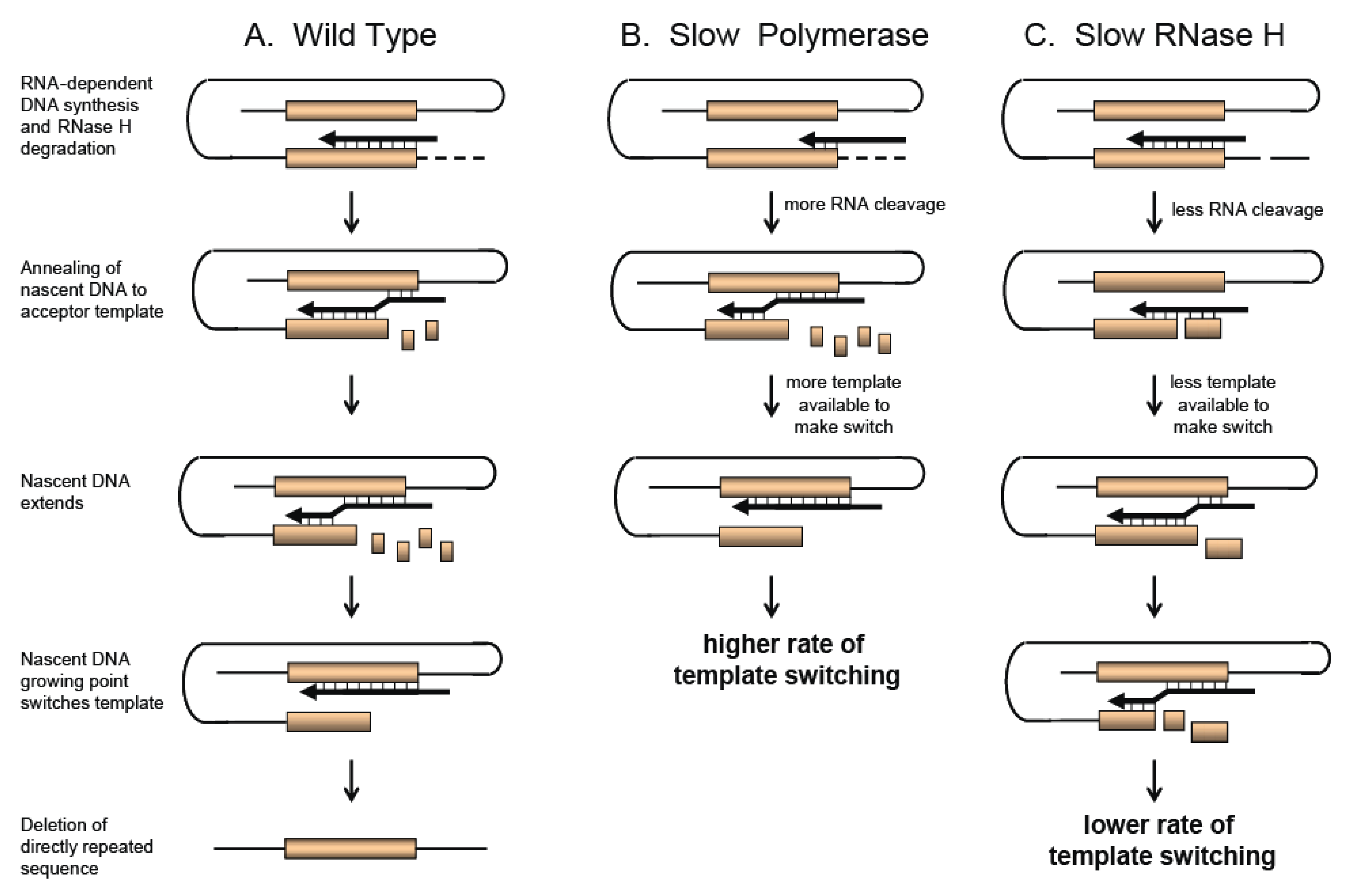
2.3. In Vitro Studies Dissecting the Biochemical Nature of Template Switching
2.4. The Use of Directly Repeated Sequences for the Development of Viral Vectors for Gene Therapy and for Probing In Vivo Reverse Transcription
3. Recombination: Rates, RNA Copackaging, and Fidelity
3.1. Recombination Rates in a Single Replication Cycle
3.2. RNA Copackaging and Its Impact on Recombination Rates
3.3. Fidelity of Recombination and Its Contribution to the Overall Mutation Frequency
4. Effects of Sequence Diversity and RNA Secondary Structures on Recombination
4.1. The Effects of Sequence Diversity on HIV-1 Recombination
4.2. The Effects of RNA Secondary Structure on HIV-1 Recombination
5. Fitness of the Progeny Recombinant
6. Recombination in the HIV-1 Epidemics
6.1. Intersubtype HIV-1 Recombinants: Evidence of Recombination-Generated Variation in Current HIV- Isolates
6.2. Detection, Replicative Fitness, and Drug-Resistance of the Intersubtype Recombinants
6.3. Recombinants and Variation Create Challenges to HIV-1 Vaccine Development
7. Conclusions
Acknowledgements
References and Notes
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Anderson, J.P.; Bradac, J.A.; Carr, J.K.; Foley, B.; Funkhouser, R.K.; Gao, F.; Hahn, B.H.; Kalish, M.L.; Kuiken, C.; et al. HIV-1 nomenclature proposal. Science 2000, 288, 55–56. [Google Scholar] [CrossRef]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global trends in molecular epidemiology of HIV-1 during 2000–2007. AIDS 2011, 25, 679–689. [Google Scholar] [CrossRef]
- Coffin, J.M.; Hughes, S.H.; Varmus, H. Retroviruses; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1997; pp. 121–160. [Google Scholar]
- Yu, S.F.; Baldwin, D.N.; Gwynn, S.R.; Yendapalli, S.; Linial, M.L. Human foamy virus replication: A pathway distinct from that of retroviruses and hepadnaviruses. Science 1996, 271, 1579–1582. [Google Scholar] [CrossRef]
- Hulme, A.E.; Perez, O.; Hope, T.J. Complementary assays reveal a relationship between HIV-1 uncoating and reverse transcription. Proc. Natl. Acad. Sci. U. S. A. 2011, 1014522108. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Retroviral recombination and reverse transcription. Science 1990, 250, 1227–1233. [Google Scholar] [CrossRef]
- Yu, H.; Jetzt, A.E.; Ron, Y.; Preston, B.D.; Dougherty, J.P. The nature of human immunodeficiency virus type 1 strand transfers. J. Biol. Chem. 1998, 273, 28384–28391. [Google Scholar] [CrossRef]
- van Wamel, J.L.; Berkhout, B. The first strand transfer during HIV-1 reverse transcription can occur either intramolecularly or intermolecularly. Virology 1998, 244, 245–251. [Google Scholar] [CrossRef]
- Coffin, J.M. Structure, replication, and recombination of retrovirus genomes: Some unifying hypotheses. J. Gen. Virol. 1979, 42, 1–26. [Google Scholar] [CrossRef]
- Hwang, C.K.; Svarovskaia, E.S.; Pathak, V.K. Dynamic copy choice: Steady state between murine leukemia virus polymerase and polymerase-dependent RNase H activity determines frequency of in vivo template switching. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 12209–12214. [Google Scholar] [CrossRef]
- Junghans, R.P.; Boone, L.R.; Skalka, A.M. Products of reverse transcription in avian retrovirus analyzed by electron microscopy. J. Virol. 1982, 43, 544–554. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Genetic consequences of packaging two RNA genomes in one retroviral particle: Pseudodiploidy and high rate of genetic recombination. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 1556–1560. [Google Scholar] [CrossRef]
- Pathak, V.K.; Hu, W.S. "Might as Well Jump!" Template switching by retroviral reverse transcriptase, defective genome formation, and recombination. Semin. Virol. 1997, 8, 141–150. [Google Scholar] [CrossRef]
- Zhuang, J.; Jetzt, A.E.; Sun, G.; Yu, H.; Klarmann, G.; Ron, Y.; Preston, B.D.; Dougherty, J.P. Human immunodeficiency virus type 1 recombination: Rate, fidelity, and putative hot spots. J. Virol. 2002, 76, 11273–11282. [Google Scholar] [CrossRef]
- Galetto, R.; Moumen, A.; Giacomoni, V.; Veron, M.; Charneau, P.; Negroni, M. The structure of HIV-1 genomic RNA in the gp120 gene determines a recombination hot spot in vivo. J. Biol. Chem. 2004, 279, 36625–36632. [Google Scholar] [CrossRef]
- Jetzt, A.E.; Yu, H.; Klarmann, G.J.; Ron, Y.; Preston, B.D.; Dougherty, J.P. High rate of recombination throughout the human immunodeficiency virus type 1 genome. J. Virol. 2000, 74, 1234–1240. [Google Scholar] [CrossRef]
- Hu, W.S.; Bowman, E.H.; Delviks, K.A.; Pathak, V.K. Homologous recombination occurs in a distinct retroviral subpopulation and exhibits high negative interference. J. Virol. 1997, 71, 6028–6036. [Google Scholar] [CrossRef]
- Katz, R.A.; Skalka, A.M. Generation of diversity in retroviruses. Annu. Rev. Genet. 1990, 24, 409–445. [Google Scholar] [CrossRef]
- Temin, H.M. Sex and recombination in retroviruses. Trends Genet. 1991, 7, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.K.; Temin, H.M. Broad spectrum of in vivo forward mutations, hypermutations, and mutational hotspots in a retroviral shuttle vector after a single replication cycle: Deletions and deletions with insertions. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 6024–6028. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, S.; Varela-Echavarria, A.; Ron, Y.; Preston, B.D.; Dougherty, J.P. Genetic rearrangements occurring during a single cycle of murine leukemia virus vector replication: Characterization and implications. J. Virol. 1995, 69, 7991–8000. [Google Scholar] [CrossRef] [PubMed]
- Czernilofsky, A.P.; Levinson, A.D.; Varmus, H.E.; Bishop, J.M.; Tischer, E.; Goodman, H.M. Nucleotide sequence of an avian sarcoma virus oncogene (src) and proposed amino acid sequence for gene product. Nature 1980, 287, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.; Kosik, E. Mutagenesis of the region between env and src of the SR-A strain of Rous sarcoma virus for the purpose of constructing helper-independent vectors. Virology 1984, 136, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Omer, C.A.; Pogue-Geile, K.; Guntaka, R.; Staskus, K.A.; Faras, A.J. Involvement of directly repeated sequences in the generation of deletions of the avian sarcoma virus src gene. J. Virol. 1983, 47, 380–382. [Google Scholar] [CrossRef]
- Rhode, B.W.; Emerman, M.; Temin, H.M. Instability of large direct repeats in retrovirus vectors. J. Virol. 1987, 61, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Lovmand, S.; Kjeldgaard, N.O.; Jorgensen, P.; Pedersen, F.S. Enhancer functions in U3 of Akv virus: A role for cooperativity of a tandem repeat unit and its flanking DNA sequences. J. Virol. 1990, 64, 3185–3191. [Google Scholar] [CrossRef]
- Pedersen, K.; Lovmand, S.; Jorgensen, E.C.; Pedersen, F.S.; Jorgensen, P. Efficient replication and expression of murine leukemia virus with major deletions in the enhancer region of U3. Virology 1992, 187, 821–824. [Google Scholar] [CrossRef]
- Pathak, V.K.; Temin, H.M. Broad spectrum of in vivo forward mutations, hypermutations, and mutational hotspots in a retroviral shuttle vector after a single replication cycle: Substitutions, frameshifts, and hypermutations. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 6019–6023. [Google Scholar] [CrossRef]
- Julias, J.G.; Hash, D.; Pathak, V.K. E- vectors: Development of novel self-inactivating and self-activating retroviral vectors for safer gene therapy. J. Virol. 1995, 69, 6839–6846. [Google Scholar] [CrossRef]
- Delviks, K.A.; Pathak, V.K. Effect of distance between homologous sequences and 3' homology on the frequency of retroviral reverse transcriptase template switching. J. Virol. 1999, 73, 7923–7932. [Google Scholar] [CrossRef]
- Bowman, R.R.; Hu, W.S.; Pathak, V.K. Relative rates of retroviral reverse transcriptase template switching during RNA- and DNA-dependent DNA synthesis. J. Virol. 1998, 72, 5198–5206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Temin, H.M. Retrovirus recombination depends on the length of sequence identity and is not error prone. J. Virol. 1994, 68, 2409–2414. [Google Scholar] [CrossRef]
- An, W.; Telesnitsky, A. Effects of varying sequence similarity on the frequency of repeat deletion during reverse transcription of a human immunodeficiency virus type 1 vector. J. Virol. 2002, 76, 7897–7902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Temin, H.M. Rate and mechanism of nonhomologous recombination during a single cycle of retroviral replication. Science 1993, 259, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Delviks, K.A.; Hwang, C.K.; Pathak, V.K. Structural determinants of murine leukemia virus reverse transcriptase that affect the frequency of template switching. J. Virol. 2000, 74, 7171–7178. [Google Scholar] [CrossRef]
- Nikolenko, G.N.; Svarovskaia, E.S.; Delviks, K.A.; Pathak, V.K. Antiretroviral drug resistance mutations in human immunodeficiency virus type 1 reverse transcriptase increase template-switching frequency. J. Virol. 2004, 78, 8761–8770. [Google Scholar] [CrossRef]
- Julias, J.G.; Pathak, V.K. Deoxyribonucleoside triphosphate pool imbalances in vivo are associated with an increased retroviral mutation rate. J. Virol. 1998, 72, 7941–7949. [Google Scholar] [CrossRef]
- Pfeiffer, J.K.; Topping, R.S.; Shin, N.H.; Telesnitsky, A. Altering the intracellular environment increases the frequency of tandem repeat deletion during Moloney murine leukemia virus reverse transcription. J. Virol. 1999, 73, 8441–8447. [Google Scholar] [CrossRef]
- An, W.; Telesnitsky, A. Frequency of direct repeat deletion in a human immunodeficiency virus type 1 vector during reverse transcription in human cells. Virology 2001, 286, 475–482. [Google Scholar] [CrossRef]
- Nikolenko, G.N.; Palmer, S.; Maldarelli, F.; Mellors, J.W.; Coffin, J.M.; Pathak, V.K. Mechanism for nucleoside analog-mediated abrogation of HIV-1 replication: Balance between RNase H activity and nucleotide excision. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 2093–2098. [Google Scholar] [CrossRef]
- Gao, L.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Insights into the multiple roles of pausing in HIV-1 reverse transcriptase-promoted strand transfers. J. Biol. Chem. 2007, 282, 6222–6231. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.J.; Buiser, R.G.; Mallaber, L.M.; Fay, P.J.; Bambara, R.A. Parameters that influence processive synthesis and site-specific termination by human immunodeficiency virus reverse transcriptase on RNA and DNA templates. Biochim. Biophys. Acta 1992, 1131, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Harrison, G.P.; Mayo, M.S.; Hunter, E.; Lever, A.M. Pausing of reverse transcriptase on retroviral RNA templates is influenced by secondary structures both 5' and 3' of the catalytic site. Nucl. Acids Res. 1998, 26, 3433–3442. [Google Scholar] [CrossRef] [PubMed]
- Klasens, B.I.; Huthoff, H.T.; Das, A.T.; Jeeninga, R.E.; Berkhout, B. The effect of template RNA structure on elongation by HIV-1 reverse transcriptase. Biochim. Biophys. Acta 1999, 1444, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Lapadat-Tapolsky, M.; De Rocquigny, H.; Van Gent, D.; Roques, B.; Plasterk, R.; Darlix, J.L. Interactions between HIV-1 nucleocapsid protein and viral DNA may have important functions in the viral life cycle. Nucl. Acids Res. 1993, 21, 831–839. [Google Scholar] [CrossRef]
- Suo, Z.; Johnson, K.A. Effect of RNA secondary structure on the kinetics of DNA synthesis catalyzed by HIV-1 reverse transcriptase. Biochemistry 1997, 36, 12459–12467. [Google Scholar] [CrossRef]
- Wu, W.; Blumberg, B.M.; Fay, P.J.; Bambara, R.A. Strand transfer mediated by human immunodeficiency virus reverse transcriptase in vitro is promoted by pausing and results in misincorporation. J. Biol. Chem. 1995, 270, 325–332. [Google Scholar] [CrossRef]
- Zhang, W.H.; Hwang, C.K.; Hu, W.S.; Gorelick, R.J.; Pathak, V.K. Zinc finger domain of murine leukemia virus nucleocapsid protein enhances the rate of viral DNA synthesis in vivo. J. Virol. 2002, 76, 7473–7484. [Google Scholar] [CrossRef]
- Lanciault, C.; Champoux, J.J. Pausing during reverse transcription increases the rate of retroviral recombination. J. Virol. 2006, 80, 2483–2494. [Google Scholar] [CrossRef]
- Klarmann, G.J.; Schauber, C.A.; Preston, B.D. Template-directed pausing of DNA synthesis by HIV-1 reverse transcriptase during polymerization of HIV-1 sequences in vitro. J. Biol. Chem. 1993, 268, 9793–9802. [Google Scholar] [CrossRef]
- Shen, W.; Gao, L.; Balakrishnan, M.; Bambara, R.A. A recombination hot spot in HIV-1 contains guanosine runs that can form a G-quartet structure and promote strand transfer in vitro. J. Biol. Chem. 2009, 284, 33883–33893. [Google Scholar] [CrossRef] [PubMed]
- Brincat, J.L.; Pfeiffer, J.K.; Telesnitsky, A. RNase H activity is required for high-frequency repeat deletion during Moloney murine leukemia virus replication. J. Virol. 2002, 76, 88–95. [Google Scholar] [CrossRef]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Barr, R.; Pathak, V.K. Mutations in human immunodeficiency virus type 1 RNase H primer grip enhance 3'-azido-3'-deoxythymidine resistance. J. Virol. 2007, 81, 6837–6845. [Google Scholar] [CrossRef] [PubMed]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Boyer, P.L.; Hughes, S.H.; Coffin, J.M.; Jere, A.; Pathak, V.K. HIV-1 reverse transcriptase connection subdomain mutations reduce template RNA degradation and enhance AZT excision. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 10943–10948. [Google Scholar] [CrossRef] [PubMed]
- Nikolenko, G.N.; Delviks-Frankenberry, K.A.; Palmer, S.; Maldarelli, F.; Fivash, M.J., Jr.; Coffin, J.M.; Pathak, V.K. Mutations in the connection domain of HIV-1 reverse transcriptase increase 3'-azido-3'-deoxythymidine resistance. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Negroni, M.; Buc, H. Copy-choice recombination by reverse transcriptases: Reshuffling of genetic markers mediated by RNA chaperones. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 6385–6390. [Google Scholar] [CrossRef]
- Roda, R.H.; Balakrishnan, M.; Hanson, M.N.; Wohrl, B.M.; Le Grice, S.F.; Roques, B.P.; Gorelick, R.J.; Bambara, R.A. Role of the reverse transcriptase, nucleocapsid protein, and template structure in the two-step transfer mechanism in retroviral recombination. J. Biol. Chem. 2003, 278, 31536–31546. [Google Scholar] [CrossRef]
- Roda, R.H.; Balakrishnan, M.; Kim, J.K.; Roques, B.P.; Fay, P.J.; Bambara, R.A. Strand transfer occurs in retroviruses by a pause-initiated two-step mechanism. J. Biol. Chem. 2002, 277, 46900–46911. [Google Scholar] [CrossRef]
- Chen, Y.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Steps of the acceptor invasion mechanism for HIV-1 minus strand strong stop transfer. J. Biol. Chem. 2003, 278, 38368–38375. [Google Scholar] [CrossRef]
- Hanson, M.N.; Balakrishnan, M.; Roques, B.P.; Bambara, R.A. Effects of donor and acceptor RNA structures on the mechanism of strand transfer by HIV-1 reverse transcriptase. J. Mol. Biol. 2005, 353, 772–787. [Google Scholar] [CrossRef]
- DeStefano, J.J.; Mallaber, L.M.; Rodriguez-Rodriguez, L.; Fay, P.J.; Bambara, R.A. Requirements for strand transfer between internal regions of heteropolymer templates by human immunodeficiency virus reverse transcriptase. J. Virol. 1992, 66, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- DeStefano, J.J.; Bambara, R.A.; Fay, P.J. The mechanism of human immunodeficiency virus reverse transcriptase-catalyzed strand transfer from internal regions of heteropolymeric RNA templates. J. Biol. Chem. 1994, 269, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Operario, D.J.; Balakrishnan, M.; Bambara, R.A.; Kim, B. Reduced dNTP interaction of human immunodeficiency virus type 1 reverse transcriptase promotes strand transfer. J. Biol. Chem. 2006, 281, 32113–32121. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Fay, P.J.; Bambara, R.A. The kissing hairpin sequence promotes recombination within the HIV-I 5' leader region. J. Biol. Chem. 2001, 276, 36482–36492. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Roques, B.P.; Fay, P.J.; Bambara, R.A. Template dimerization promotes an acceptor invasion-induced transfer mechanism during human immunodeficiency virus type 1 minus-strand synthesis. J. Virol. 2003, 77, 4710–4721. [Google Scholar] [CrossRef]
- Tanese, N.; Telesnitsky, A.; Goff, S.P. Abortive reverse transcription by mutants of Moloney murine leukemia virus deficient in the reverse transcriptase-associated RNase H function. J. Virol. 1991, 65, 4387–4397. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, M.; Schulze, T.; Larder, B.A.; Moelling, K. Mutations within the RNase H domain of human immunodeficiency virus type 1 reverse transcriptase abolish virus infectivity. J. Gen. Virol. 1991, 72, 59–66. [Google Scholar] [CrossRef]
- Purohit, V.; Balakrishnan, M.; Kim, B.; Bambara, R.A. Evidence that HIV-1 reverse transcriptase employs the DNA 3' end-directed primary/secondary RNase H cleavage mechanism during synthesis and strand transfer. J. Biol. Chem. 2005, 280, 40534–40543. [Google Scholar] [CrossRef]
- DeStefano, J.J.; Mallaber, L.M.; Fay, P.J.; Bambara, R.A. Determinants of the RNase H cleavage specificity of human immunodeficiency virus reverse transcriptase. Nucl. Acids Res. 1993, 21, 4330–4338. [Google Scholar] [CrossRef]
- Furfine, E.S.; Reardon, J.E. Reverse transcriptase.RNase H from the human immunodeficiency virus. Relationship of the DNA polymerase and RNA hydrolysis activities. J. Biol. Chem. 1991, 266, 406–412. [Google Scholar] [CrossRef]
- Schultz, S.J.; Zhang, M.; Champoux, J.J. Recognition of internal cleavage sites by retroviral RNases H. J. Mol. Biol. 2004, 344, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Balakrishnan, M.; Palaniappan, C.; Fay, P.J.; Bambara, R.A. The sequential mechanism of HIV reverse transcriptase RNase H. J. Biol. Chem. 2000, 275, 37664–37671. [Google Scholar] [CrossRef] [PubMed]
- Delviks, K.A.; Hu, W.S.; Pathak, V.K. Psi- vectors: Murine leukemia virus-based self-inactivating and self-activating retroviral vectors. J. Virol. 1997, 71, 6218–6224. [Google Scholar] [CrossRef] [PubMed]
- Delviks, K.A.; Pathak, V.K. Development of murine leukemia virus-based self-activating vectors that efficiently delete the selectable drug resistance gene during reverse transcription. J. Virol. 1999, 73, 8837–8842. [Google Scholar] [CrossRef]
- Liu, S.; Abbondanzieri, E.A.; Rausch, J.W.; Le Grice, S.F.; Zhuang, X. Slide into action: Dynamic shuttling of HIV reverse transcriptase on nucleic acid substrates. Science 2008, 322, 1092–1097. [Google Scholar] [CrossRef]
- Anderson, J.A.; Bowman, E.H.; Hu, W.S. Retroviral recombination rates do not increase linearly with marker distance and are limited by the size of the recombining subpopulation. J. Virol. 1998, 72, 1195–1202. [Google Scholar] [CrossRef]
- Rhodes, T.; Wargo, H.; Hu, W.S. High rates of human immunodeficiency virus type 1 recombination: Near-random segregation of markers one kilobase apart in one round of viral replication. J. Virol. 2003, 77, 11193–11200. [Google Scholar] [CrossRef]
- Rhodes, T.D.; Nikolaitchik, O.; Chen, J.; Powell, D.; Hu, W.S. Genetic recombination of human immunodeficiency virus type 1 in one round of viral replication: Effects of genetic distance, target cells, accessory genes, and lack of high negative interference in crossover events. J. Virol. 2005, 79, 1666–1677. [Google Scholar] [CrossRef]
- Chen, J.; Powell, D.; Hu, W.S. High frequency of genetic recombination is a common feature of primate lentivirus replication. J. Virol. 2006, 80, 9651–9658. [Google Scholar] [CrossRef]
- Anderson, J.A.; Pathak, V.K.; Hu, W.S. Effect of the murine leukemia virus extended packaging signal on the rates and locations of retroviral recombination. J. Virol. 2000, 74, 6953–6963. [Google Scholar] [CrossRef]
- Zhuang, J.; Mukherjee, S.; Ron, Y.; Dougherty, J.P. High rate of genetic recombination in murine leukemia virus: Implications for influencing proviral ploidy. J. Virol. 2006, 80, 6706–6711. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.P.; Rhodes, T.D.; Chen, J.; Fu, W.; Hu, W.S. Identification of a major restriction in HIV-1 intersubtype recombination. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 9002–9007. [Google Scholar] [CrossRef]
- Clever, J.L.; Wong, M.L.; Parslow, T.G. Requirements for kissing-loop-mediated dimerization of human immunodeficiency virus RNA. J. Virol. 1996, 70, 5902–5908. [Google Scholar] [CrossRef] [PubMed]
- Muriaux, D.; Fosse, P.; Paoletti, J. A kissing complex together with a stable dimer is involved in the HIV-1Lai RNA dimerization process in vitro. Biochemistry 1996, 35, 5075–5082. [Google Scholar] [CrossRef] [PubMed]
- Paillart, J.C.; Skripkin, E.; Ehresmann, B.; Ehresmann, C.; Marquet, R. A loop-loop "kissing" complex is the essential part of the dimer linkage of genomic HIV-1 RNA. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 5572–5577. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.P.S.; Chen, J.; Nikolaitchik, O.A.; Hu, W.-S. Molecular determinants of HIV-1 intersubtype recombination potential. Virology 2007, 363, 437–446. [Google Scholar] [CrossRef]
- Moore, M.D.; Fu, W.; Nikolaitchik, O.; Chen, J.; Ptak, R.G.; Hu, W.S. Dimer initiation signal of human immunodeficiency virus type 1: Its role in partner selection during RNA copackaging and its effects on recombination. J. Virol. 2007, 81, 4002–4011. [Google Scholar] [CrossRef]
- Hussein, I.T.; Ni, N.; Galli, A.; Chen, J.; Moore, M.D.; Hu, W.S. Delineation of the preferences and requirements of the human immunodeficiency virus type 1 dimerization initiation signal by using an in vivo cell-based selection approach. J. Virol. 2010, 84, 6866–6875. [Google Scholar] [CrossRef]
- Moore, M.D.; Nikolaitchik, O.A.; Chen, J.; Hammarskjold, M.L.; Rekosh, D.; Hu, W.S. Probing the HIV-1 genomic RNA trafficking pathway and dimerization by genetic recombination and single virion analyses. PLoS Pathog. 2009, 5, e1000627. [Google Scholar] [CrossRef]
- Moore, M.D.; Hu, W.S. HIV-1 RNA dimerization: It takes two to tango. AIDS Rev. 2009, 11, 91–102. [Google Scholar]
- Paillart, J.C.; Shehu-Xhilaga, M.; Marquet, R.; Mak, J. Dimerization of retroviral RNA genomes: An inseparable pair. Nat. Rev. Microbiol. 2004, 2, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nikolaitchik, O.; Singh, J.; Wright, A.; Bencsics, C.E.; Coffin, J.M.; Ni, N.; Lockett, S.; Pathak, V.K.; Hu, W.S. High efficiency of HIV-1 genomic RNA packaging and heterozygote formation revealed by single virion analysis. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 13535–13540. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.A.; An, W.; King, S.R.; Telesnitsky, A. Nonrandom dimerization of murine leukemia virus genomic RNAs. J. Virol. 2004, 78, 12129–12139. [Google Scholar] [CrossRef] [PubMed]
- Kharytonchyk, S.A.; Kireyeva, A.I.; Osipovich, A.B.; Fomin, I.K. Evidence for preferential copackaging of Moloney murine leukemia virus genomic RNAs transcribed in the same chromosomal site. Retrovirology 2005, 2, 3. [Google Scholar] [CrossRef]
- Dang, Q.; Chen, J.; Unutmaz, D.; Coffin, J.M.; Pathak, V.K.; Powell, D.; KewalRamani, V.N.; Maldarelli, F.; Hu, W.S. Nonrandom HIV-1 infection and double infection via direct and cell-mediated pathways. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 632–637. [Google Scholar] [CrossRef]
- Chen, J.; Dang, Q.; Unutmaz, D.; Pathak, V.K.; Maldarelli, F.; Powell, D.; Hu, W.S. Mechanisms of nonrandom human immunodeficiency virus type 1 infection and double infection: Preference in virus entry is important but is not the sole factor. J. Virol. 2005, 79, 4140–4149. [Google Scholar] [CrossRef]
- Del Portillo, A.; Tripodi, J.; Najfeld, V.; Wodarz, D.; Levy, D.N.; Chen, B.K. Multiploid Inheritance of HIV-1 during Cell-to-Cell Infection. J. Virol. 2011, 85, 7169–7176. [Google Scholar] [CrossRef]
- Jung, A.; Maier, R.; Vartanian, J.P.; Bocharov, G.; Jung, V.; Fischer, U.; Meese, E.; Wain-Hobson, S.; Meyerhans, A. Recombination: Multiply infected spleen cells in HIV patients. Nature 2002, 418, 144. [Google Scholar] [CrossRef]
- Josefsson, L.; King, M.S.; Makitalo, B.; Brannstrom, J.; Shao, W.; Maldarelli, F.; Kearney, M.F.; Hu, W.S.; Chen, J.; Gaines, H.; et al. Majority of CD4+ T cells from peripheral blood of HIV-1-infected individuals contain only one HIV DNA molecule. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 11199–11204. [Google Scholar] [CrossRef]
- Chin, M.P.; Lee, S.K.; Chen, J.; Nikolaitchik, O.A.; Powell, D.A.; Fivash, M.J., Jr.; Hu, W.S. Long-range recombination gradient between HIV-1 subtypes B and C variants caused by sequence differences in the dimerization initiation signal region. J. Mol. Biol. 2008, 377, 1324–1333. [Google Scholar] [CrossRef]
- Nikolaitchik, O.A.; Galli, A.; Moore, M.D.; Pathak, V.K.; Hu, W.-S. Multiple barriers to recombination between divergent HIV-1 variants revealed by a dual-marker recombination assay. J. Mol. Biol. 2011, 407, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Galli, A.; Kearney, M.; Nikolaitchik, O.A.; Yu, S.; Chin, M.P.S.; Maldarelli, F.; Coffin, J.M.; Pathak, V.K.; Hu, W.-S. Patterns of human immunodeficiency virus type 1 recombination ex vivo provide evidence for coadaptation of distant sites, resulting in purifying selection for intersubtype recombinants during replication. J. Virol. 2010, 84, 7651–7661. [Google Scholar] [CrossRef] [PubMed]
- Baird, H.A.; Galetto, R.; Gao, Y.; Simon-Loriere, E.; Abreha, M.; Archer, J.; Fan, J.; Robertson, D.L.; Arts, E.J.; Negroni, M. Sequence determinants of breakpoint location during HIV-1 intersubtype recombination. Nucl. Acids Res. 2006, 34, 5203–5216. [Google Scholar] [CrossRef]
- Baird, H.A.; Gao, Y.; Galetto, R.; Lalonde, M.; Anthony, R.M.; Giacomoni, V.; Abreha, M.; Destefano, J.J.; Negroni, M.; Arts, E.J. Influence of sequence identity and unique breakpoints on the frequency of intersubtype HIV-1 recombination. Retrovirology 2006, 3, 91. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Galetto, R.; Hamoudi, M.; Archer, J.; Lefeuvre, P.; Martin, D.P.; Robertson, D.L.; Negroni, M. Molecular mechanisms of recombination restriction in the envelope gene of the human immunodeficiency virus. PLoS Pathog. 2009, 5, e1000418. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Paraskevis, D.; Vandamme, A.M.; Magiorkinis, E.; Sypsa, V.; Hatzakis, A. In vivo characteristics of human immunodeficiency virus type 1 intersubtype recombination: Determination of hot spots and correlation with sequence similarity. J. Gen. Virol. 2003, 84, 2715–2722. [Google Scholar] [CrossRef]
- Archer, J.; Pinney, J.W.; Fan, J.; Simon-Loriere, E.; Arts, E.J.; Negroni, M.; Robertson, D.L. Identifying the important HIV-1 recombination breakpoints. PLoS Comput. Biol. 2008, 4, e1000178. [Google Scholar] [CrossRef]
- Fan, J.; Negroni, M.; Robertson, D.L. The distribution of HIV-1 recombination breakpoints. Infect. Genet. Evol. 2007, 7, 717–723. [Google Scholar] [CrossRef]
- Galli, A.; Lai, A.; Corvasce, S.; Saladini, F.; Riva, C.; Deho, L.; Caramma, I.; Franzetti, M.; Romano, L.; Galli, M.; et al. Recombination analysis and structure prediction show correlation between breakpoint clusters and RNA hairpins in the pol gene of human immunodeficiency virus type 1 unique recombinant forms. J. Gen. Virol. 2008, 89, 3119–3125. [Google Scholar] [CrossRef]
- Dykes, C.; Balakrishnan, M.; Planelles, V.; Zhu, Y.; Bambara, R.A.; Demeter, L.M. Identification of a preferred region for recombination and mutation in HIV-1 gag. Virology 2004, 326, 262–279. [Google Scholar] [CrossRef]
- Galetto, R.; Giacomoni, V.; Veron, M.; Negroni, M. Dissection of a circumscribed recombination hot spot in HIV-1 after a single infectious cycle. J. Biol. Chem. 2006, 281, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Palaniappan, C.; Wu, W.; Fay, P.J.; Bambara, R.A. Evidence for a unique mechanism of strand transfer from the transactivation response region of HIV-1. J. Biol. Chem. 1997, 272, 16769–16777. [Google Scholar] [CrossRef] [PubMed]
- Moumen, A.; Polomack, L.; Unge, T.; Veron, M.; Buc, H.; Negroni, M. Evidence for a mechanism of recombination during reverse transcription dependent on the structure of the acceptor RNA. J. Biol. Chem. 2003, 278, 15973–15982. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.M.; Dang, K.K.; Gorelick, R.J.; Leonard, C.W.; Bess Jr, J.W.; Swanstrom, R.; Burch, C.L.; Weeks, K.M. Architecture and secondary structure of an entire HIV-1 RNA genome. Nature 2009, 460, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Martin, D.P.; Weeks, K.M.; Negroni, M. RNA structures facilitate recombination-mediated gene swapping in HIV-1. J. Virol. 2010, 84, 12675–12682. [Google Scholar] [CrossRef] [PubMed]
- McCutchan, F.E. Global epidemiology of HIV. J. Med. Virol. 2006, 78, S7–S12. [Google Scholar] [CrossRef]
- Thomson, M.M.; Najera, R. Travel and the introduction of human immunodeficiency virus type 1 non-B subtype genetic forms into Western countries. Clin. Infect. Dis. 2001, 32, 1732–1737. [Google Scholar] [CrossRef]
- Thomson, M.M.; Najera, R. Molecular epidemiology of HIV-1 variants in the global AIDS pandemic: An update. AIDS Rev. 2005, 7, 210–224. [Google Scholar]
- Thomson, M.M.; Perez-Alvarez, L.; Najera, R. Molecular epidemiology of HIV-1 genetic forms and its significance for vaccine development and therapy. Lancet Infect. Dis. 2002, 2, 461–471. [Google Scholar] [CrossRef]
- Los Alamos HIV Sequence Database. Available online: http://www.hiv.lanl.gov/ (accessed on 8 August 2011).
- Oelrichs, R.B.; Crowe, S.M. The molecular epidemiology of HIV-1 in South and East Asia. Curr. HIV Res. 2003, 1, 239–248. [Google Scholar] [CrossRef]
- Abecasis, A.B.; Lemey, P.; Vidal, N.; de Oliveira, T.; Peeters, M.; Camacho, R.; Shapiro, B.; Rambaut, A.; Vandamme, A.-M. Recombination confounds the early evolutionary history of human immunodeficiency virus type 1: Subtype G is a circulating recombinant form. J. Virol. 2007, 81, 8543–8551. [Google Scholar] [CrossRef] [PubMed]
- Tee, K.K.; Pybus, O.G.; Li, X.J.; Han, X.; Shang, H.; Kamarulzaman, A.; Takebe, Y. Temporal and spatial dynamics of human immunodeficiency virus type 1 circulating recombinant forms 08_BC and 07_BC in Asia. J. Virol. 2008, 82, 9206–9215. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, M.T.; Ruibal, I.; Villahermosa, M.L.; Diaz, H.; Delgado, E.; Parga, E.V.; Perez-Alvarez, L.; de Armas, M.B.; Cuevas, L.; Medrano, L.; Noa, E.; Osmanov, S.; Najera, R.; Thomson, M.M. High HIV-1 genetic diversity in Cuba. AIDS 2002, 16, 1643–1653. [Google Scholar] [CrossRef]
- Delgado, E.; Thomson, M.M.; Villahermosa, M.L.; Sierra, M.; Ocampo, A.; Miralles, C.; Rodriguez-Perez, R.; Diz-Aren, J.; Ojea-de Castro, R.; Losada, E.; et al. Identification of a newly characterized HIV-1 BG intersubtype circulating recombinant form in Galicia, Spain, which exhibits a pseudotype-like virion structure. J. Acquir. Immune Defic. Syndr. 2002, 29, 536–543. [Google Scholar] [CrossRef]
- Thomson, M.M.; Delgado, E.; Manjón, N.; Ocampo, A.; Villahermosa, M.L.; Mariño, A.; Herrero, I.; Cuevas, M.T.; Vázquez-de Parga, E.; Pérez-Alvarez, L.; et al. Spanish Group for Antiretroviral Studies in Galicia HIV-1 genetic diversity in Galicia Spain: BG intersubtype recombinant viruses circulating among injecting drug users. AIDS 2001, 15, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. AIDS 2006, 20, W13–W23. [Google Scholar] [CrossRef]
- Bourlet, T.; Signori-Schmuck, A.; Roche, L.; Icard, V.; Saoudin, H.; Trabaud, M.A.; Tardy, J.C.; Morand, P.; Pozzetto, B.; Ecochard, R.; et al. HIV-1 load comparison using four commercial real-time assays. J. Clin. Microbiol. 2011, 49, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Church, D.; Gregson, D.; Lloyd, T.; Klein, M.; Beckthold, B.; Laupland, K.; Gill, M.J. Comparison of the RealTime HIV-1, COBAS TaqMan 48 v1.0, Easy Q v1.2, and Versant v3.0 assays for Determination of HIV-1 Viral Loads in a Cohort of Canadian Patients with Diverse HIV Subtype Infections. J. Clin. Microbiol. 2010, 49, 118–124. [Google Scholar] [CrossRef]
- Foglieni, B.; Candotti, D.; Guarnori, I.; Raffaele, L.; Berzuini, A.; Spreafico, M.; Orani, A.; Rossotti, R.; Rossi, D.; Allain, J.P.; et al. A cluster of human immunodeficiency virus Type 1 recombinant form escaping detection by commercial genomic amplification assays. Transfusion 2011, 51, 719–730. [Google Scholar] [CrossRef]
- Korenromp, E.L.; Williams, B.G.; Schmid, G.P.; Dye, C. Clinical prognostic value of RNA viral load and CD4 cell counts during untreated HIV-1 infection—A quantitative review. PLoS One 2009, 4, e5950. [Google Scholar] [CrossRef]
- Mellors, J.W.; Rinaldo, C.R., Jr.; Gupta, P.; White, R.M.; Todd, J.A.; Kingsley, L.A. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science 1996, 272, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Arien, K.K.; Abraha, A.; Quinones-Mateu, M.E.; Kestens, L.; Vanham, G.; Arts, E.J. The replicative fitness of primary human immunodeficiency virus type 1 (HIV-1) group M, HIV-1 group O, and HIV-2 isolates. J. Virol. 2005, 79, 8979–8990. [Google Scholar] [CrossRef] [PubMed]
- Njai, H.F.; Gali, Y.; Vanham, G.; Clybergh, C.; Jennes, W.; Vidal, N.; Butel, C.; Mpoudi-Ngolle, E.; Peeters, M.; Arien, K.K. The predominance of human immunodeficiency virus type 1 (HIV-1) circulating recombinant form 02 (CRF02_AG) in West Central Africa may be related to its replicative fitness. Retrovirology 2006, 3, 40. [Google Scholar] [CrossRef]
- Konings, F.A.; Burda, S.T.; Urbanski, M.M.; Zhong, P.; Nadas, A.; Nyambi, P.N. Human immunodeficiency virus type 1 (HIV-1) circulating recombinant form 02_AG (CRF02_AG) has a higher in vitro replicative capacity than its parental subtypes A and G. J. Med. Virol. 2006, 78, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.Y.; Takebe, Y.; Weniger, B.G.; Luo, C.C.; Kalish, M.L.; Auwanit, W.; Yamazaki, S.; Gayle, H.D.; Young, N.L.; Schochetman, G. Independent introduction of two major HIV-1 genotypes into distinct high-risk populations in Thailand. Lancet 1993, 341, 1171–1174. [Google Scholar] [CrossRef]
- Hudgens, M.G.; Longini, I.M., Jr.; Vanichseni, S.; Hu, D.J.; Kitayaporn, D.; Mock, P.A.; Halloran, M.E.; Satten, G.A.; Choopanya, K.; Mastro, T.D. Subtype-specific transmission probabilities for human immunodeficiency virus type 1 among injecting drug users in Bangkok, Thailand. Am. J. Epidemiol. 2002, 155, 159–168. [Google Scholar] [CrossRef]
- Laurent, C.; Bourgeois, A.; Faye, M.A.; Mougnutou, R.; Seydi, M.; Gueye, M.; Liegeois, F.; Kane, C.T.; Butel, C.; Mbuagbaw, J.; et al. No difference in clinical progression between patients infected with the predominant human immunodeficiency virus type 1 circulating recombinant form (CRF) 02_AG strain and patients not infected with CRF02_AG, in Western and West-Central Africa: A four-year prospective multicenter study. J. Infect. Dis. 2002, 186, 486–492. [Google Scholar]
- Easterbrook, P.J.; Smith, M.; Mullen, J.; O'Shea, S.; Chrystie, I.; de Ruiter, A.; Tatt, I.D.; Geretti, A.M.; Zuckerman, M. Impact of HIV-1 viral subtype on disease progression and response to antiretroviral therapy. J. Int. AIDS Soc. 2010, 13, 4. [Google Scholar] [CrossRef]
- Baeten, J.M.; Chohan, B.; Lavreys, L.; Chohan, V.; McClelland, R.S.; Certain, L.; Mandaliya, K.; Jaoko, W.; Overbaugh, J. HIV-1 subtype D infection is associated with faster disease progression than subtype A in spite of similar plasma HIV-1 loads. J. Infect. Dis. 2007, 195, 1177–1180. [Google Scholar] [CrossRef]
- Vergne, L.; Peeters, M.; Mpoudi-Ngole, E.; Bourgeois, A.; Liegeois, F.; Toure-Kane, C.; Mboup, S.; Mulanga-Kabeya, C.; Saman, E.; Jourdan, J.; et al. Genetic diversity of protease and reverse transcriptase sequences in non-subtype-b human immunodeficiency virus type 1 strains: Evidence of many minor drug resistance mutations in treatment-naive patients. J. Clin. Microbiol. 2000, 38, 3919–3925. [Google Scholar] [CrossRef]
- Kantor, R.; Katzenstein, D.A.; Efron, B.; Carvalho, A.P.; Wynhoven, B.; Cane, P.; Clarke, J.; Sirivichayakul, S.; Soares, M.A.; Snoeck, J.; et al. Impact of HIV-1 subtype and antiretroviral therapy on protease and reverse transcriptase genotype: Results of a global collaboration. PLoS Med. 2005, 2, e112. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M. HIV-1 subtypes: Epidemiology and significance for HIV management. Curr. Opin. Infect. Dis. 2006, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M.; Harrison, L.; Green, H.; Sabin, C.; Hill, T.; Fearnhill, E.; Pillay, D.; Dunn, D. Effect of HIV-1 subtype on virologic and immunologic response to starting highly active antiretroviral therapy. Clin. Infect. Dis. 2009, 48, 1296–1305. [Google Scholar] [CrossRef]
- Bannister, W.P.; Ruiz, L.; Loveday, C.; Vella, S.; Zilmer, K.; Kjaer, J.; Knysz, B.; Phillips, A.N.; Mocroft, A. HIV-1 subtypes and response to combination antiretroviral therapy in Europe. Antivir. Ther. 2006, 11, 707–715. [Google Scholar] [PubMed]
- Bocket, L.; Cheret, A.; Deuffic-Burban, S.; Choisy, P.; Gerard, Y.; de la Tribonniere, X.; Viget, N.; Ajana, F.; Goffard, A.; Barin, F.; Mouton, Y.; Yazdanpanah, Y. Impact of human immunodeficiency virus type 1 subtype on first-line antiretroviral therapy effectiveness. Antivir. Ther. 2005, 10, 247–254. [Google Scholar] [CrossRef]
- Delviks-Frankenberry, K.A.; Nikolenko, G.N.; Maldarelli, F.; Hase, S.; Takebe, Y.; Pathak, V.K. Subtype-specific differences in the human immunodeficiency virus type 1 reverse transcriptase connection subdomain of CRF01_AE are associated with higher levels of resistance to 3'-azido-3'-deoxythymidine. J. Virol. 2009, 83, 8502–8513. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Posada, D.; Crandall, K.A.; Holmes, E.C. The causes and consequences of HIV evolution. Nat. Rev. Genet. 2004, 5, 52–61. [Google Scholar] [CrossRef]
- Batorsky, R.; Kearney, M.F.; Palmer, S.E.; Maldarelli, F.; Rouzine, I.M.; Coffin, J.M. Estimate of effective recombination rate and average selection coefficient for HIV in chronic infection. Proc. Natl. Acad. Sci. U. S. A. 2011. [Google Scholar] [CrossRef]
- Kearney, M.; Maldarelli, F.; Shao, W.; Margolick, J.B.; Daar, E.S.; Mellors, J.W.; Rao, V.; Coffin, J.M.; Palmer, S. Human immunodeficiency virus type 1 population genetics and adaptation in newly infected individuals. J. Virol. 2009, 83, 2715–2727. [Google Scholar] [CrossRef]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 7552–7557. [Google Scholar] [CrossRef]
- Li, H.; Bar, K.J.; Wang, S.; Decker, J.M.; Chen, Y.; Sun, C.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Learn, G.H.; Morgan, C.J.; et al. High Multiplicity Infection by HIV-1 in men who have sex with men. PLoS Pathog. 2010, 6, e1000890. [Google Scholar] [CrossRef] [PubMed]
- Streeck, H.; Li, B.; Poon, A.F.; Schneidewind, A.; Gladden, A.D.; Power, K.A.; Daskalakis, D.; Bazner, S.; Zuniga, R.; Brander, C.; et al. Immune-driven recombination and loss of control after HIV superinfection. J. Exp. Med. 2008, 205, 1789–1796. [Google Scholar] [CrossRef]
- Reynolds, M.R.; Weiler, A.M.; Piaskowski, S.M.; Kolar, H.L.; Hessell, A.J.; Weiker, M.; Weisgrau, K.L.; Leon, E.J.; Rogers, W.E.; Makowsky, R.; et al. Macaques vaccinated with simian immunodeficiency virus SIVmac239Delta nef delay acquisition and control replication after repeated low-dose heterologous SIV challenge. J. Virol. 2010, 84, 9190–9199. [Google Scholar] [CrossRef] [PubMed]
- Gundlach, B.R.; Lewis, M.G.; Sopper, S.; Schnell, T.; Sodroski, J.; Stahl-Hennig, C.; Uberla, K. Evidence for recombination of live, attenuated immunodeficiency virus vaccine with challenge virus to a more virulent strain. J. Virol. 2000, 74, 3537–3542. [Google Scholar] [CrossRef] [PubMed]
- Metzner, K.J.; Moretto, W.J.; Donahoe, S.M.; Jin, X.; Gettie, A.; Montefiori, D.C.; Marx, P.A.; Binley, J.M.; Nixon, D.F.; Connor, R.I. Evaluation of CD8+ T-cell and antibody responses following transient increased viraemia in rhesus macaques infected with live, attenuated simian immunodeficiency virus. J. Gen. Virol. 2005, 86, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.R.; Weiler, A.M.; Weisgrau, K.L.; Piaskowski, S.M.; Furlott, J.R.; Weinfurter, J.T.; Kaizu, M.; Soma, T.; Leon, E.J.; MacNair, C.; et al. Macaques vaccinated with live-attenuated SIV control replication of heterologous virus. J. Exp. Med. 2008, 205, 2537–2550. [Google Scholar] [CrossRef]
- Fischer, W.; Perkins, S.; Theiler, J.; Bhattacharya, T.; Yusim, K.; Funkhouser, R.; Kuiken, C.; Haynes, B.; Letvin, N.L.; Walker, B.D.; et al. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nat. Med. 2007, 13, 100–106. [Google Scholar] [CrossRef]
- Nickle, D.C.; Rolland, M.; Jensen, M.A.; Pond, S.L.; Deng, W.; Seligman, M.; Heckerman, D.; Mullins, J.I.; Jojic, N. Coping with viral diversity in HIV vaccine design. PLoS Comput. Biol. 2007, 3, e75. [Google Scholar] [CrossRef]
- Santra, S.; Liao, H.X.; Zhang, R.; Muldoon, M.; Watson, S.; Fischer, W.; Theiler, J.; Szinger, J.; Balachandran, H.; Buzby, A.; et al. Mosaic vaccines elicit CD8+ T lymphocyte responses that confer enhanced immune coverage of diverse HIV strains in monkeys. Nat. Med. 2010, 16, 324–328. [Google Scholar] [CrossRef]
- Barouch, D.H.; O'Brien, K.L.; Simmons, N.L.; King, S.L.; Abbink, P.; Maxfield, L.F.; Sun, Y.H.; La Porte, A.; Riggs, A.M.; Lynch, D.M.; et al. Mosaic HIV-1 vaccines expand the breadth and depth of cellular immune responses in rhesus monkeys. Nat. Med. 2010, 16, 319–323. [Google Scholar] [CrossRef]





© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Delviks-Frankenberry, K.; Galli, A.; Nikolaitchik, O.; Mens, H.; Pathak, V.K.; Hu, W.-S. Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses 2011, 3, 1650-1680. https://doi.org/10.3390/v3091650
Delviks-Frankenberry K, Galli A, Nikolaitchik O, Mens H, Pathak VK, Hu W-S. Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses. 2011; 3(9):1650-1680. https://doi.org/10.3390/v3091650
Chicago/Turabian StyleDelviks-Frankenberry, Krista, Andrea Galli, Olga Nikolaitchik, Helene Mens, Vinay K. Pathak, and Wei-Shau Hu. 2011. "Mechanisms and Factors that Influence High Frequency Retroviral Recombination" Viruses 3, no. 9: 1650-1680. https://doi.org/10.3390/v3091650
APA StyleDelviks-Frankenberry, K., Galli, A., Nikolaitchik, O., Mens, H., Pathak, V. K., & Hu, W. -S. (2011). Mechanisms and Factors that Influence High Frequency Retroviral Recombination. Viruses, 3(9), 1650-1680. https://doi.org/10.3390/v3091650