
A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Monocyte-Derived Macrophages
2.2. Generation of CD16 Expressing SK6 Cells
2.3. Viruses
2.4. Antibody-Dependent Enhancement of Infection
2.5. Vaccination Challenge Experiment
2.6. Virological Analyses
2.7. Serum Neutralization Assay
2.8. Statistics
3. Results
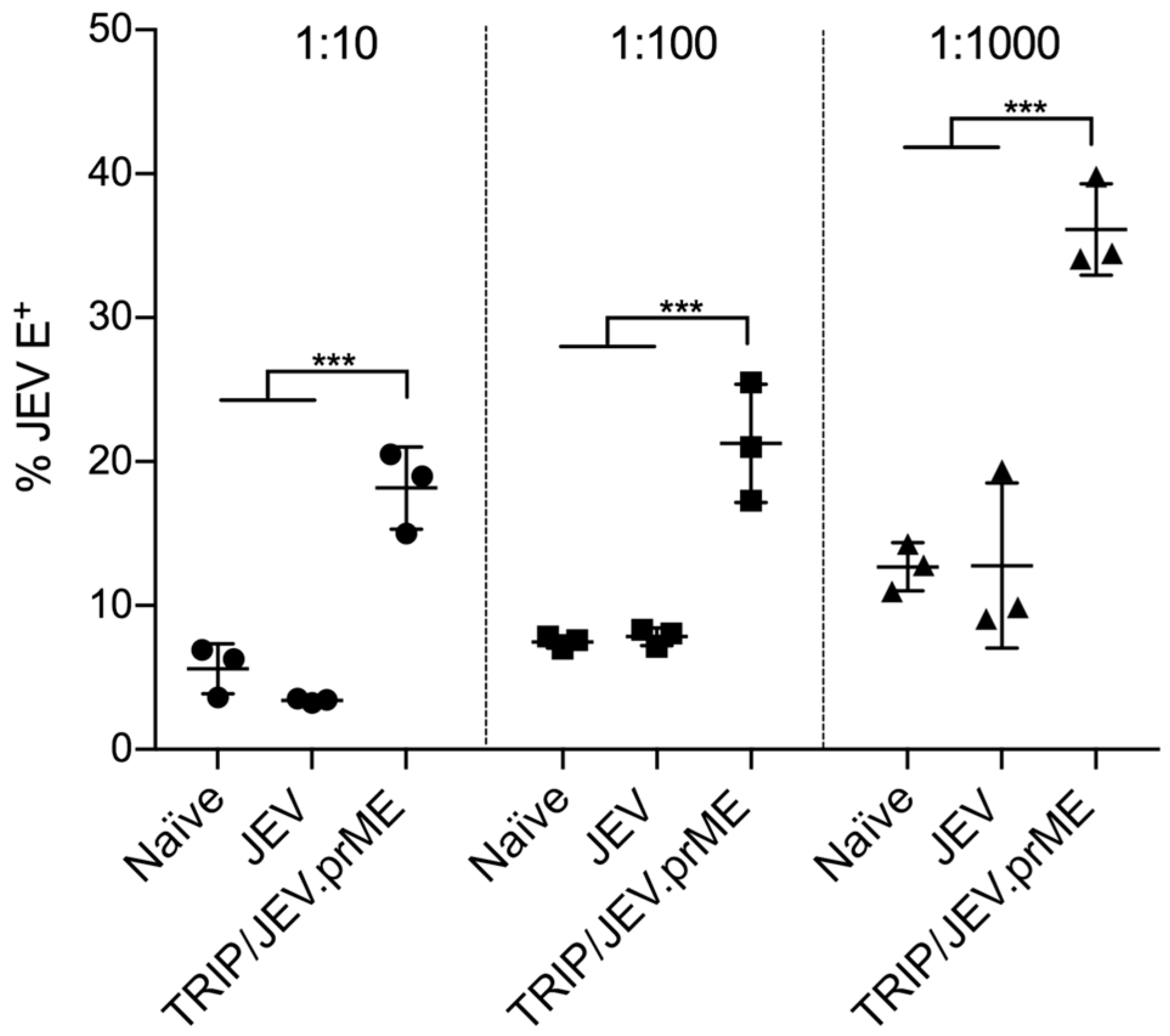
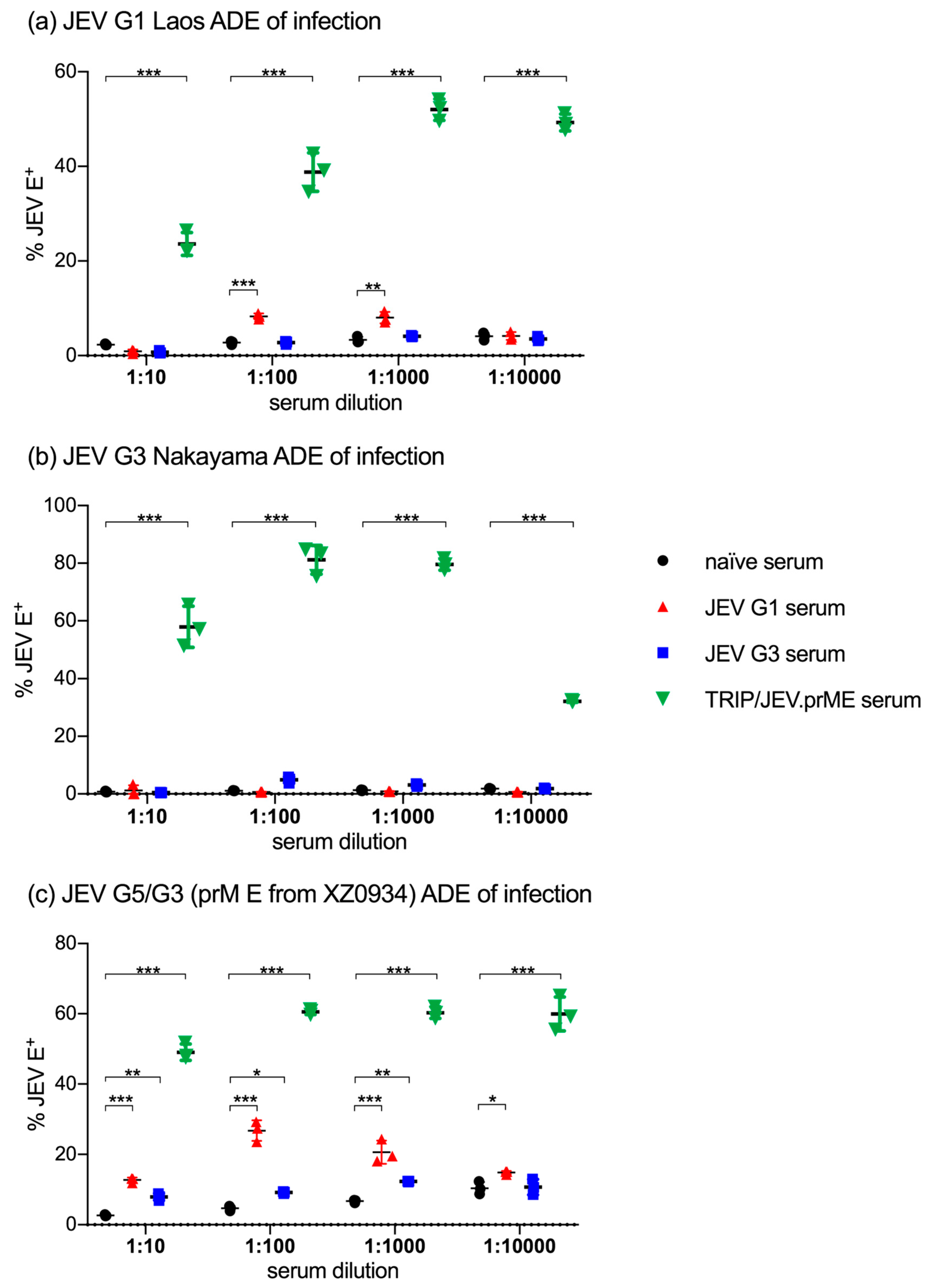
3.1. TRIP/JEV.prME Induces ADE of Macrophage Infection
3.2. TRIP/JEV.prME-Antibodies Strongly Enhance JEV Infection of Cells Expressing FcγRIII
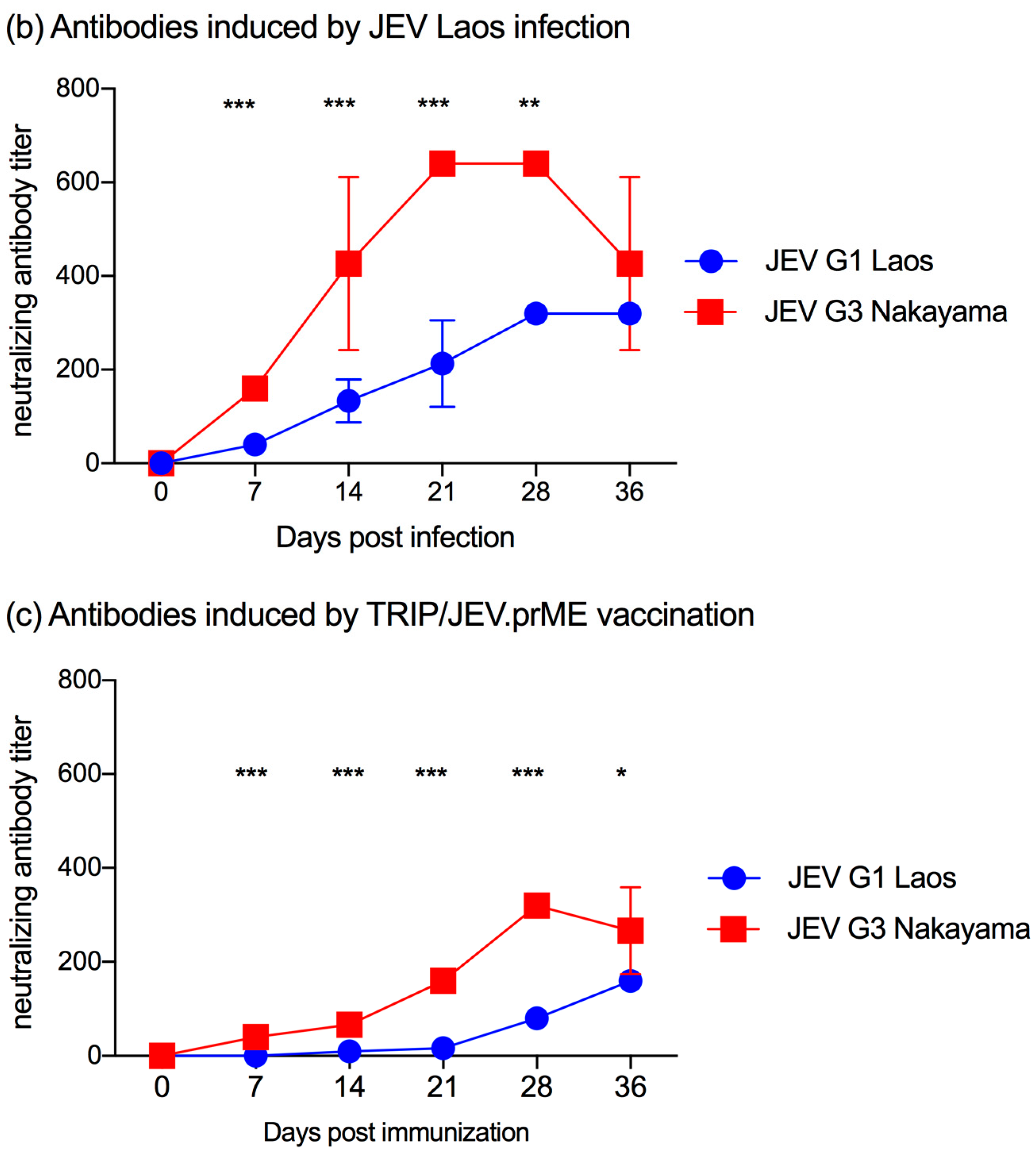
3.3. Antibody Responses Induced by TRIP/JEV.prME Vaccine and JEV Infection
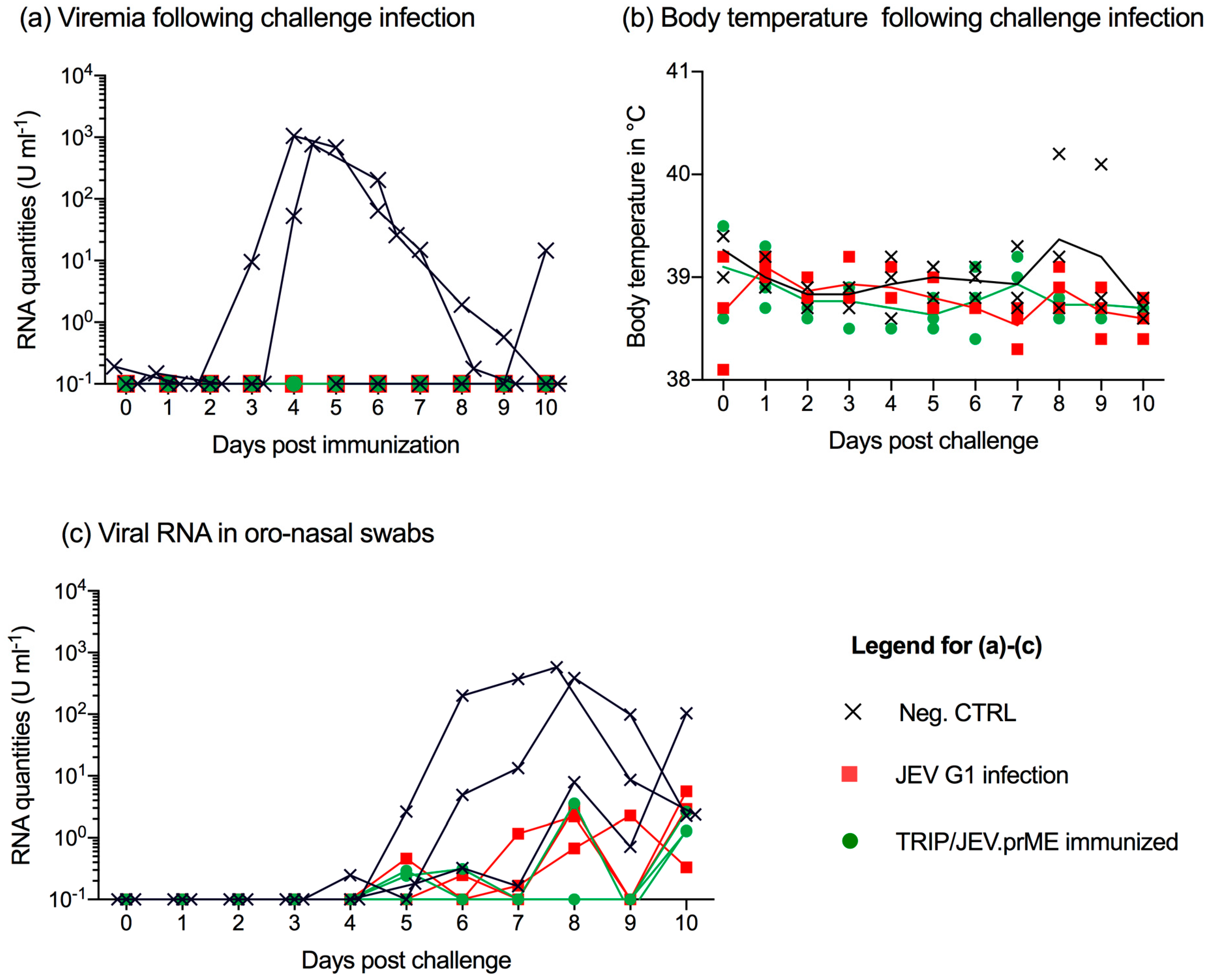
3.4. TRIP/JEV.prME Vaccine and Previous JEV Infection Induce Protection against Viremia
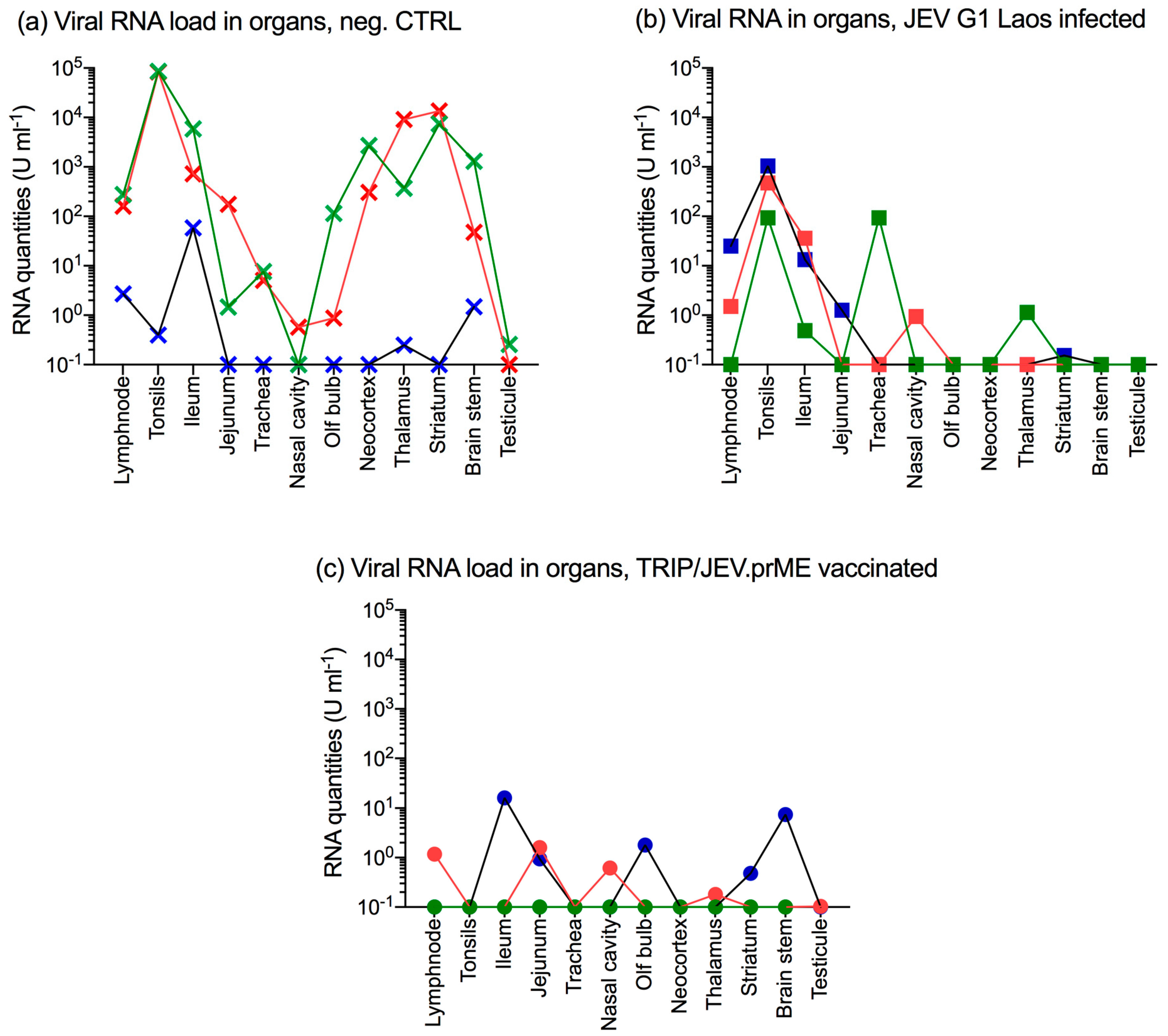
3.5. JEV Immunization Does Not Completely Prevent Oro-Nasal Shedding of Challenge Virus
3.6. JEV Immunization Does Not Provide Sterile Immunity
3.7. JEV G1 Persistence after JEV G3 Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weaver, S.C.; Barrett, A.D. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Impoinvil, D.E.; Baylis, M.; Solomon, T. Japanese encephalitis: On the One Health agenda. Curr. Top. Microbiol. Immunol. 2013, 365, 205–247. [Google Scholar] [PubMed]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccines Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Hernandez-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.P. Review of climate, landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.Y.; Balasuriya, U.B.; Lee, C.K. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58–77. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, H.; Wang, H.; Fu, S.; Guo, Z.; Liang, G. Southernmost Asia is the source of Japanese encephalitis virus (genotype 1) diversity from which the viruses disperse and evolve throughout Asia. PLoS Negl. Trop. Dis. 2013, 7, e2459. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Ward, M.J.; Brown, A.J.; Barrett, A.D. Phylogeography of Japanese encephalitis virus: Genotype is associated with climate. PLoS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar] [CrossRef] [PubMed]
- Takhampunya, R.; Kim, H.C.; Tippayachai, B.; Kengluecha, A.; Klein, T.A.; Lee, W.J.; Grieco, J.; Evans, B.P. Emergence of Japanese encephalitis virus genotype V in the Republic of Korea. Virol. J. 2011, 8, 449. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Fu, S.H.; Chen, W.X.; Wang, H.Y.; Guo, Y.H.; Liu, Q.Y.; Li, Y.X.; Luo, H.M.; Da, W.; Duo Ji, D.Z.; et al. Genotype V Japanese encephalitis virus is emerging. PLoS Negl. Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef] [PubMed]
- Dubischar-Kastner, K.; Kanesa-Thasan, N. Vaccinating against Japanese encephalitis virus: What have we learned from recent clinical trials? Expert. Rev. Vaccines 2012, 11, 1159–1161. [Google Scholar] [CrossRef] [PubMed]
- Larena, M.; Prow, N.A.; Hall, R.A.; Petrovsky, N.; Lobigs, M. JE-ADVAX vaccine protection against Japanese encephalitis virus mediated by memory B cells in the absence of CD8(+) T cells and pre-exposure neutralizing antibody. J. Virol. 2013, 87, 4395–4402. [Google Scholar] [CrossRef] [PubMed]
- Kimura-Kuroda, J.; Yasui, K. Protection of mice against Japanese encephalitis virus by passive administration with monoclonal antibodies. J. Immunol. 1988, 141, 3606–3610. [Google Scholar] [PubMed]
- Van Gessel, Y.; Klade, C.S.; Putnak, R.; Formica, A.; Krasaesub, S.; Spruth, M.; Cena, B.; Tungtaeng, A.; Gettayacamin, M.; Dewasthaly, S. Correlation of protection against Japanese encephalitis virus and JE vaccine (IXIARO((R)) induced neutralizing antibody titers. Vaccine 2011, 29, 5925–5931. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.R.; Whitehead, S.S. Immune response to dengue virus and prospects for a vaccine. Annu. Rev. Immunol. 2011, 29, 587–619. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P. Dengue Antibody and Zika: Friend or Foe? Trends Immunol. 2016, 37, 635–636. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Foo, S.S.; Bruzzone, R.; Dinh, L.V.; King, N.J.; Mahalingam, S. Fc receptors in antibody-dependent enhancement of viral infections. Immunol. Rev. 2015, 268, 340–364. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.J.; Smith, D.W.; Broom, A.K.; Mackenzie, J.S.; Hall, R.A.; Shellam, G.R.; McMinn, P.C. Antibody-dependent enhancement of Murray Valley encephalitis virus virulence in mice. J. Gen. Virol. 2003, 84, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Buckley, A. Antibody-dependent enhancement of yellow fever and Japanese encephalitis virus neurovirulence. J. Gen. Virol. 1989, 70, 1605–1608. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Moyer, J.T.; Izumi, T.; Gresser, I.; Mc, C.J. Ecologic studies of Japanese encephalitis virus in Japan. VI. Swine infection. Am. J. Trop. Med. Hyg. 1959, 8, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, M.E.; Garcia-Nicolas, O.; Brechbuhl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, M.E.; Garcia-Nicolas, O.; Brechbuhl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Vet. Res. 2016, 47, 34. [Google Scholar] [CrossRef] [PubMed]
- De Wispelaere, M.; Ricklin, M.; Souque, P.; Frenkiel, M.P.; Paulous, S.; Garcia-Nicolas, O.; Summerfield, A.; Charneau, P.; Despres, P.A. Lentiviral Vector Expressing Japanese Encephalitis Virus-like Particles Elicits Broad Neutralizing Antibody Response in Pigs. PLoS Negl. Trop. Dis. 2015, 9, e0004081. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Nicolas, O.; Baumann, A.; Vielle, N.J.; Gomez-Laguna, J.; Quereda, J.J.; Pallares, F.J.; Ramis, G.; Carrasco, L.; Summerfield, A. Virulence and genotype-associated infectivity of interferon-treated macrophages by porcine reproductive and respiratory syndrome viruses. Virus Res. 2014, 179, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.P.; Neuhaus, V.; Guzylack-Piriou, L.; Ruggli, N.; McCullough, K.C.; Summerfield, A. Toll-like receptor 7 and MyD88 knockdown by lentivirus-mediated RNA interference to porcine dendritic cell subsets. Gene Ther. 2007, 14, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [PubMed]
- Aubry, F.; Vongsouvath, M.; Nougairede, A.; Phetsouvanh, R.; Sibounheuang, B.; Charrel, R.; Rattanavong, S.; Phommasone, K.; Sengvilaipraserth, O.; de Lamballerie, X.; et al. Complete Genome of a Genotype I Japanese Encephalitis Virus Isolated from a Patient with Encephalitis in Vientiane, Lao PDR. Genome Announc. 2013, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.K.; Kweon, C.H.; Kim, B.H.; Lim, S.I.; Kim, S.H.; Kwon, J.H.; Han, H.R. TaqMan reverse transcription polymerase chain reaction for the detection of Japanese encephalitis virus. J. Vet. Sci. 2004, 5, 345–351. [Google Scholar] [PubMed]
- Pierson, T.C.; Xu, Q.; Nelson, S.; Oliphant, T.; Nybakken, G.E.; Fremont, D.H.; Diamond, M.S. The stoichiometry of antibody-mediated neutralization and enhancement of West Nile virus infection. Cell. Host Microbe 2007, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science 2010, 328, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Rodenhuis-Zybert, I.A.; van der Schaar, H.M.; da Silva Voorham, J.M.; van der Ende-Metselaar, H.; Lei, H.Y.; Wilschut, J.; Smit, J.M. Immature dengue virus: A veiled pathogen? PLoS Pathog. 2010, 6, e1000718. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.; Jost, C.A.; Xu, Q.; Ess, J.; Martin, J.E.; Oliphant, T.; Whitehead, S.S.; Durbin, A.P.; Graham, B.S.; Diamond, M.S.; et al. Maturation of West Nile virus modulates sensitivity to antibody-mediated neutralization. PLoS Pathog. 2008, 4, e1000060. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. Degrees of maturity: The complex structure and biology of flaviviruses. Curr. Opin. Virol. 2012, 2, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Wikan, N.; Libsittikul, S.; Yoksan, S.; Auewarakul, P.; Smith, D.R. Delayed antibody dependent enhancement of low passage dengue virus 4 isolates. BMC Res. Notes 2015, 8, 399. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Mukherjee, S.; Kuhn, R.J.; Pierson, T.C. Combined effects of the structural heterogeneity and dynamics of flaviviruses on antibody recognition. J. Virol. 2014, 88, 11726–11737. [Google Scholar] [CrossRef] [PubMed]
- He, R.T.; Innis, B.L.; Nisalak, A.; Usawattanakul, W.; Wang, S.; Kalayanarooj, S.; Anderson, R. Antibodies that block virus attachment to Vero cells are a major component of the human neutralizing antibody response against dengue virus type 2. J. Med. Virol. 1995, 45, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Putnak, R.; Barvir, D.A.; Burrous, J.M.; Dubois, D.R.; D’Andrea, V.M.; Hoke, C.H.; Sadoff, J.C.; Eckels, K.H. Development of a purified, inactivated, dengue-2 virus vaccine prototype in Vero cells: Immunogenicity and protection in mice and rhesus monkeys. J. Infect. Dis. 1996, 174, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.R.; Dowd, K.A.; Engle, M.; Tesh, R.B.; Johnson, S.; Pierson, T.C.; Diamond, M.S. Poorly neutralizing cross-reactive antibodies against the fusion loop of West Nile virus envelope protein protect in vivo via Fcgamma receptor and complement-dependent effector mechanisms. J. Virol. 2011, 85, 11567–11580. [Google Scholar] [CrossRef] [PubMed]
- Rodenhuis-Zybert, I.A.; da Silva Voorham, J.M.; Torres, S.; van de Pol, D.; Smit, J.M. Antibodies against immature virions are not a discriminating factor for dengue disease severity. PLoS Negl. Trop. Dis. 2015, 9, e0003564. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum | JEV Strain | FRNT50 |
|---|---|---|
| JEV G1 antisera | G1 | 1:80 |
| G3 | 1:320 | |
| G3/G5 | 1:160 | |
| JEV G3 antisera | G1 | 1:20 |
| G3 | 1:320–640 | |
| G3/G5 | 1:80 | |
| TRIP/JEV.prME antisera | G1 | 1:40 |
| G3 | 1:160 | |
| G3/G5 | 1:60 |
| Specificity | Primer and Probe Sequence (5′-3′) | Concentration (nM) | |
|---|---|---|---|
| 3′ NTR JEV | forward | GGTGTAAGGACTAGAGGTTAGAGG | 200 |
| reverse | ATTCCCAGGTGTCAATATGCTGTT | 200 | |
| probe | FAM-CCCGTGGAAACAACATCATGCGGC-BHQ-1 | 100 | |
| JEV G1 Laos | forward | GACAGGATAAAGTCATGTGCGT | 200 |
| reverse | CCTGACGTTGGTCTTTCAAC | 200 | |
| probe | FAM-CCGTCTCGGAAGCAGGTCCC-BHQ-1 | 100 | |
| JEV G3 Nakayama | forward | CAGGGTCATCTAGTGTGATTTAAGG | 1600 |
| reverse | CAGTCCTCCTGGGACTGAGA | 1600 | |
| probe | FAM-TGCTGGCCTGACTCCATATGCA-BHQ-1 | 200 |
| Group/Pig Number 1 | Nakayama-Specific RT-qPCR | Laos-Specific RT-qPCR |
|---|---|---|
| Neg. CTRL, #1516 | 5.2 × 100 | negative |
| Neg. CTRL, #1517 | 4.6 × 105 | negative |
| Neg. CTRL, #1518 | 2.0 × 105 | negative |
| JEV G1, #1512 | negative | 3.2 × 101 |
| JEV G1, #1513 | negative | 1.7 × 101 |
| JEV G1, #1521 | negative | 2.9 × 100 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Nicolás, O.; Ricklin, M.E.; Liniger, M.; Vielle, N.J.; Python, S.; Souque, P.; Charneau, P.; Summerfield, A. A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs. Viruses 2017, 9, 124. https://doi.org/10.3390/v9050124
García-Nicolás O, Ricklin ME, Liniger M, Vielle NJ, Python S, Souque P, Charneau P, Summerfield A. A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs. Viruses. 2017; 9(5):124. https://doi.org/10.3390/v9050124
Chicago/Turabian StyleGarcía-Nicolás, Obdulio, Meret E. Ricklin, Matthias Liniger, Nathalie J. Vielle, Sylvie Python, Philippe Souque, Pierre Charneau, and Artur Summerfield. 2017. "A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs" Viruses 9, no. 5: 124. https://doi.org/10.3390/v9050124
APA StyleGarcía-Nicolás, O., Ricklin, M. E., Liniger, M., Vielle, N. J., Python, S., Souque, P., Charneau, P., & Summerfield, A. (2017). A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs. Viruses, 9(5), 124. https://doi.org/10.3390/v9050124