1. Introduction
Crystals are widely distributed in plants, with calcium oxalate being the most common, although crystals occasionally contain magnesium, sulfur, silicon, and other chemical elements. Some organic substances are birefringent and could be detected in polarizing microscopes, e.g., starch and lignified cell walls. To determine the composition of crystals, anatomical studies are not enough, also the determination of the elemental composition is necessary [
1,
2,
3,
4]. The taxonomic significance of the shape and localization of crystals is known; they can characterize species, genera, and even suprageneric taxa; crystal characters have phylogenetic signals in some taxa [
5,
6]. Crystal features are widely used in the identification of wood [
7] and bark [
3].
The most important work on crystals in the fruits of the Apiaceae family was published at the end of the 19th century by J. Rompel [
8]. The author studied about 200 species from 86 genera of Apiaceae and 2 genera of Araliaceae. He identified three types of crystals, differing in morphology and location in the pericarp: (1) Type
Hydrocotyle, where one layer of cells with simple crystals is located in the innermost part of the mesocarp, directly on the lignified endocarp; (2) Type
Sanicula, where druses of crystals are scattered throughout the pericarp, including the commissure area; and (3) Type
Scandix, simple or complex crystals are located around the columella and throughout the commissure, but are absent in other parts of the pericarp. Rompel used the system proposed by G. Bentham [
9] to present his data; the presence of each type of crystal or their absence turned out to be closely correlated with the tribes or subtribes of this system.
Additionally, Drude [
10] studied fruit anatomy and crystals in many Apiaceae species, using crystal features to build a system of the family that botanists accepted for 100 years. He assigned genera with
Hydrocotyle-type crystals to the subfamily Hydrocotyloideae, those with
Sanicula-type crystals to the subfamily Saniculoiddeae, and in the subfamily Apioideae,
Scandix-type crystals are characteristic of the tribe Scandiceae. Despite the abundance of molecular data and the change in our understanding of the family system, crystals remain an important feature in the delimitation of taxa [
11].
In the decades that followed Rompel’s [
5] publication, there were few new reports of crystals. This is partly due to the fact that many methods of anatomical studies involve the treatment of the material with acids that dissolve calcium oxalate. Purposeful searches for crystals were not carried out. There have been reports of calcium oxalate crystals in
Ammi majus [
12,
13] and
Bilacunaria microcarpa [
14]. An intensive study of the anatomy of Apiaceae fruits is currently being carried out by botanists at the University of Johannesburg, where crystals were first identified in the fruits of many South African genera [
15,
16,
17,
18,
19,
20]. Based on new data on the distribution of crystals, anatomy, morphology, and molecular phylogenetics, a new division of the Apiaceae family into tribes has been proposed [
21].
Crystals were found in the fruits and leaves of the Madagascar genus
Billburttia, however, they had a shape not characteristic of calcium oxalate, did not dissolve in acid, and energy-dispersive analysis showed the absence of calcium in them [
22]. These crystals could have resulted from the precipitation of some organic matter.
We have lately studied [
23] the anatomy of
Modesciadium involucratum,
Stoibrax dichotomum, and
S. pomelianum fruits using bright field light microscopy and have identified small crystals on the commissural side and in the exocarp. The genera
Ammi,
Modesciadium, and
Stoibrax have also recently been studied by molecular phylogenetics, all of which have been assigned to the clade Apieae:
Stoibrax to subclade A, and
Ammi and
Modesciadium to subclade B3 [
24,
25]. The authors noted that they used acids to prepare anatomical slides of the fruits, and it was therefore impossible to determine the presence/absence of calcium oxalate crystals.
In this paper, we present the results of studying such an important taxonomic feature as the presence, shape, and localization crystals in the fruits of Apiaceae. Some species of the B3 subclade of the Apieae clade were studied: Ammi majus, Modesciadium involucratum, Deverra aphylla, and Rutheopsis tortuosa. We used conventional and polarizing optical microscopy, as well as scanning electron microscopy in natural environment mode (ESEM) and usual SEM. The chemical composition of the crystals and tissues of the fruits was studied using energy-dispersive spectroscopy (EDS). Additionally, species with classical Sanicula-type (Sanicula rubriflora) and Scandix-type (Chaerophyllum bulbosum) crystals were studied.
Drude’s [
10] system of the family Apiaceae was accepted by many botanists in the twentieth century, but the accumulation of morphological and especially molecular data required its revision. The recognition and circumscription of tribes remain largely unsettled [
11]. For this reason, we use the informal category “clade” for family subdivisions.
3. Results
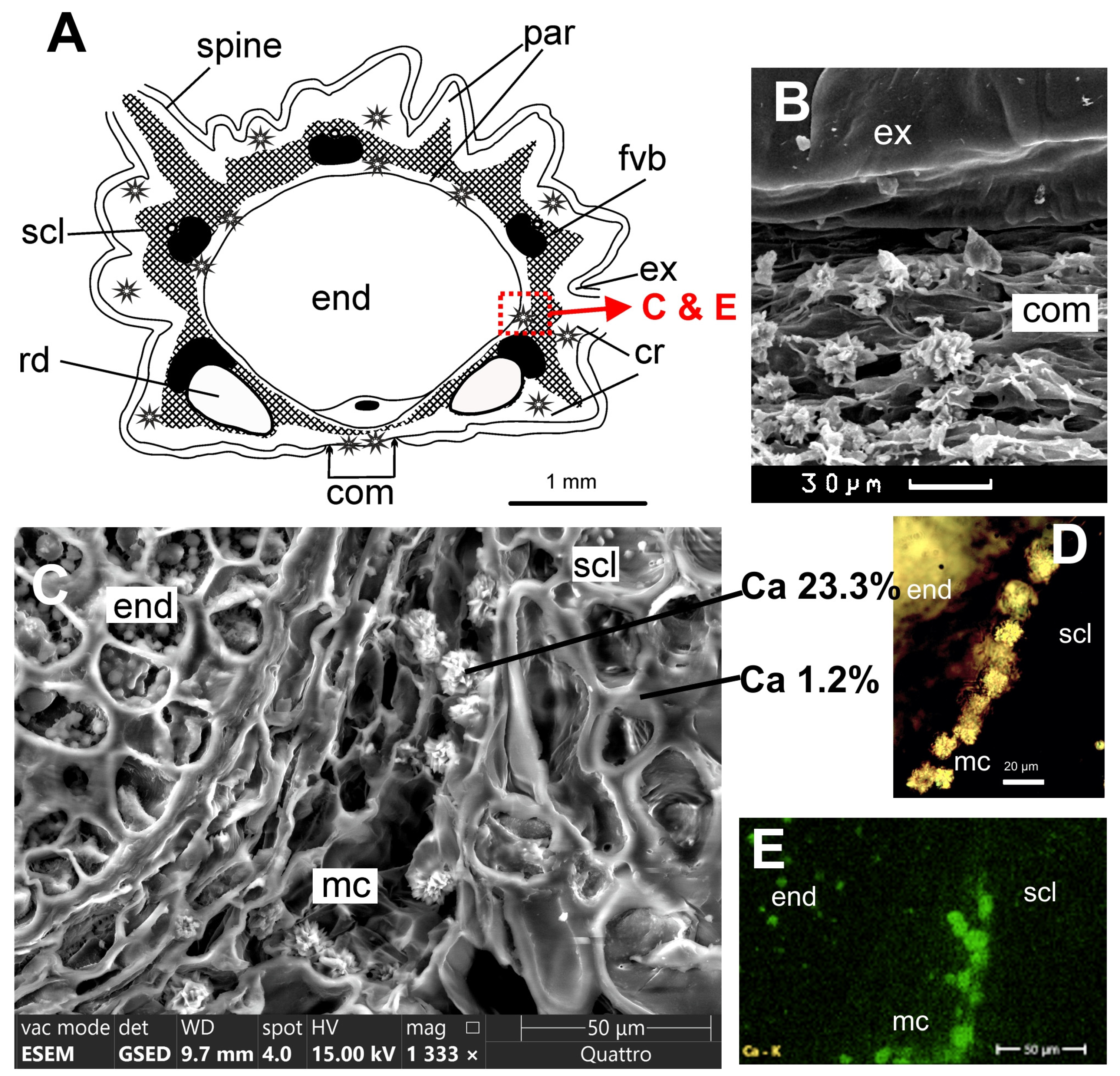
Clade Saniculeae:
Sanicula rubriflora F. Schmidt ex Maxim. (
Figure 1).
Fruits of two ovate mericarps, covered with hook-shaped spines; 4–6 mm long, 3.5–4 mm broad; primary ribs equal, on the surface not prominent; mericarps not compressed dorsally; commissure narrow. Outer cell walls of exocarp thick (10–15 µm). Spines have longitudinal bundles of lignified cells, which are connected to rib vascular bundles; so the mesocarp on the transverse section consists of non-lignified parenchyma cells and a continuous layer of sclerenchyma; non-lignified parenchyma forms a thin layer under the sclerenchyma and fills the outer part of the mesocarp. Vallecular and commissural secretory ducts are obsolete; one or both marginal rib secretory ducts are very large, and the remaining rib ducts are small.
Crystals on commissure are clearly visible in an optical stereomicroscope at a magnification of 20–40× as a white powder. A scanning microscope shows large druses up to 10 µm in size on the commissure surface, cell walls were destroyed during fruit dehiscence. On the transverse sections, a lot of druses are visible in the inner layers of the mesocarp parenchyma, and scattered druses in the outer parenchyma of the mesocarp.
ESEM and EDS. Two maps and 15 spectra were studied. The druses are composed of carbon, oxygen, and calcium. On the elemental maps of transverse sections of the mericarp (inner part of pericarp with druses and some area of seed), the calcium content is 2.4–3.5%. The spectra of druses show calcium content from 9.9% to 23.7%, sclerenchyma 1.2–1.7%, endosperm cells 3.0–8.7%, and thick-walled exocarp cells 0.6%.
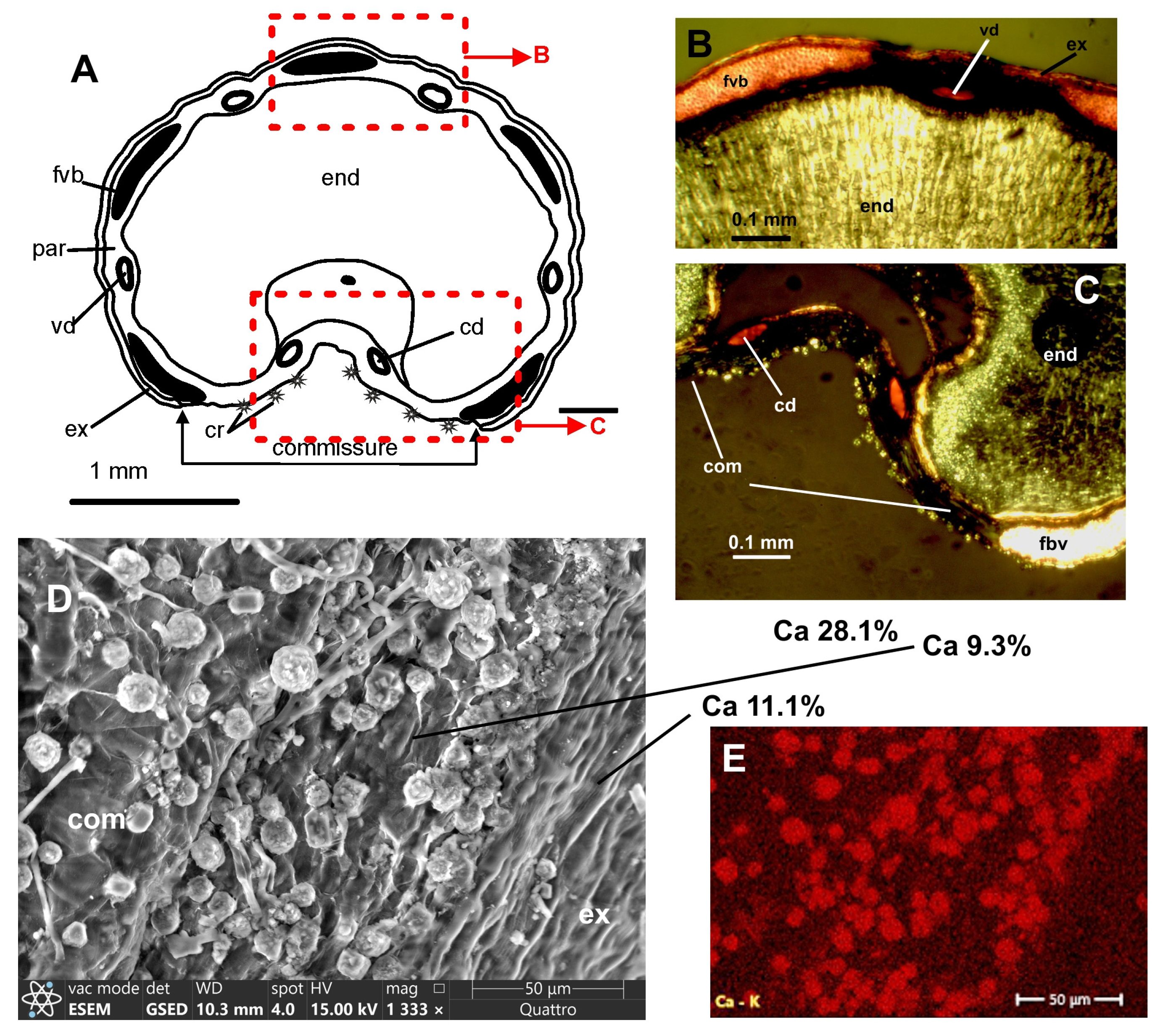
Clade Scandicinae:
Chaerophyllum bulbosum L. (
Figure 2).
Fruits with linear-lanceolate or linear mericarps; 4–6 mm long, 1–1.5 mm broad, primary ribs equal, broad, blunt; mericarps not compressed dorsally; exocarp cells small, commissure broad; mesocarp parenchyma not lignified; fibrovascular bundles large; vallecular secretory ducts solitary, two on the commissural side; rib secretory ducts small, present in some rib only.
In mature fruits, crystals densely cover the surface of the commissure, they are clearly visible in an optical stereomicroscope at a magnification of ×20–40 as a white powder. Spherical aggregates of crystals up to 15 µm in diameter are situated on the commissure surface, cell walls were destroyed during fruit dehiscence. Rompel [
8] observed that in young fruits the crystals are single prismatic, and in the process of ripening they are combined into groups. There is a lot of calcium oxalate in the endosperm, near the commissure, and the dorsal side of the endosperm is almost devoid of crystals. The exocarp and mesocarp cells do not contain crystals.
ESEM and EDS. Six maps and sixteen spectra were studied. The crystals are composed of carbon, oxygen, and calcium. Elemental maps of the commissural surface revealed from 15.8 to 16.9% of calcium. On the map of the transverse section near the dorsal side of the mericarp, (sclerenchyma of the vascular bundle, parenchyma of the mesocarp, and seed) calcium content is 1.0%. Spectra of globular clusters of crystals on the commissure show from 26.5 to 31.6% calcium; spectra of the commissure surface without visible crystals show 6.3% and 9.3%; the outer surface of the exocarp near the commissure 11.1%; endosperm on a transverse section near the dorsal side of the mericarp contains 0.3%; sclerenchyma of the vascular bundle 0.8% calcium.
The four species below belong to the clade Apieae, subclade B3.
The species studied have small fruits (1–4 × 0.6–1.6 mm) covered with hairs or papillae; pericarp non-lignified except for fibrovascular bundles; mesocarp cells above vallecular and commissural secretory ducts are relatively large, square, not compressed.
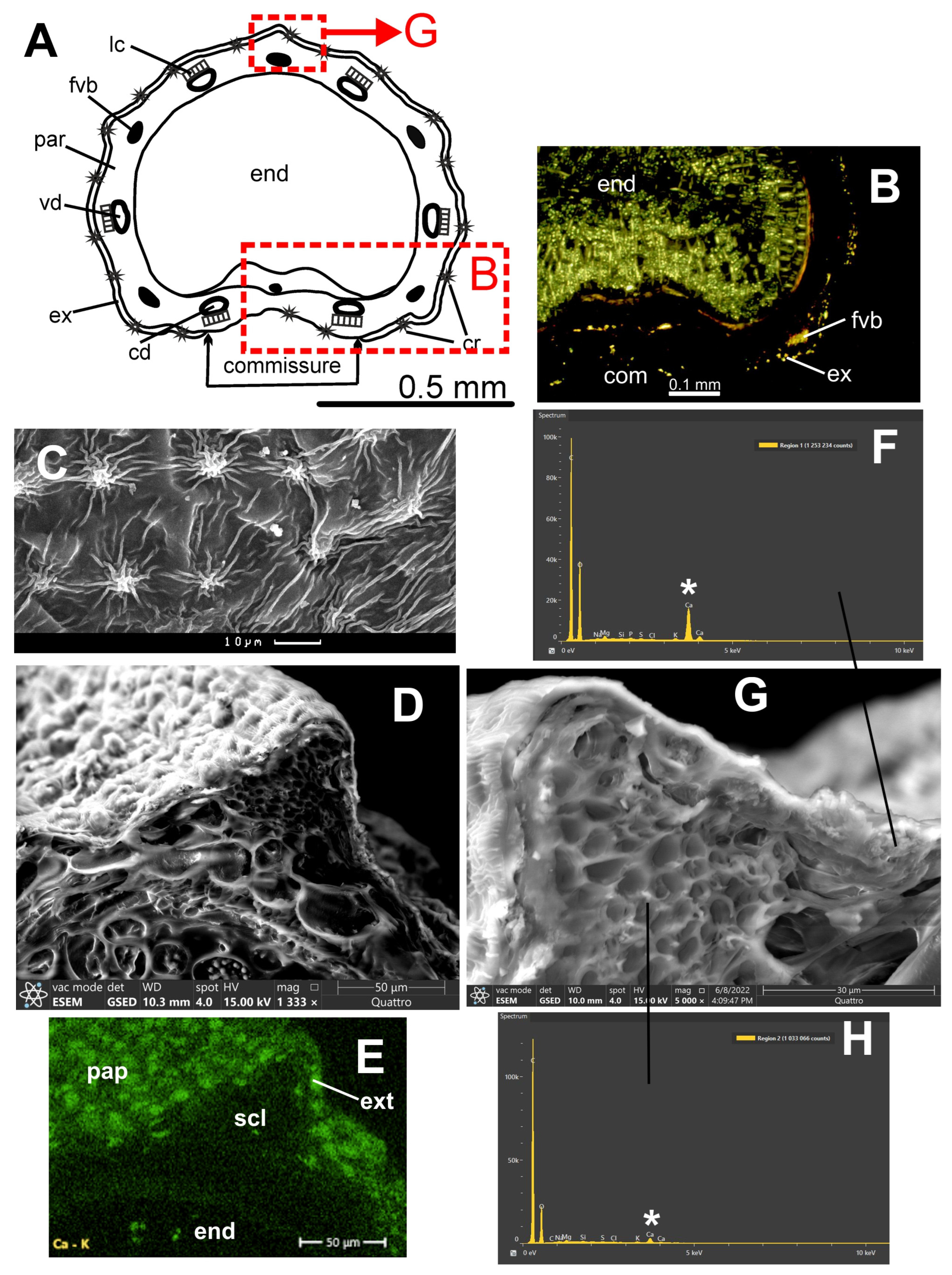
Fruits with ovate or oblong mericarps; 2–3.5 mm long, 0.8–1.5 mm broad, primary ribs equal, filiform. Mericarps not compressed dorsally; exocarp cells small, commissure of intermediate width; mesocarp parenchyma not lignified; mesocarp cells above vallecular and commissural secretory ducts are relatively large, square, not compressed; vallecular secretory ducts solitary, two on the commissural side; rib secretory ducts small, present in some rib only. Fruit surface covered with conical papillae 7–10 µm in diam. having striato-knotted cuticular foldings.
According to polarizing microscopy, crystals are located in the exocarp and on the commissural side. In a bright-field optical microscope, crystals ca. 3 μm are visible. Hydrochloric acid dissolves these crystals.
ESEM and EDS. Eight maps and seven spectra were studied. The exocarp contains small (3–5 μm) druses and crystals of irregular shape 1–5 μm in size. On the elemental maps, calcium compounds are marked in the exocarp, in some places in the mesocarp, and in the endosperm; apparently, calcium compounds do not form crystals in the mesocarp. The crystals are composed of carbon, oxygen, and calcium. The calcium content in the crystals is 16.2–21.0%. The sclerenchyma of the fibrovascular bundle contains 3.3–4.0% calcium; endosperm cells without obvious crystals contain 0.8%.
Figure 3E shows that calcium was determined in the short papillae of the exocarp, although the crystals are located under the cell wall.
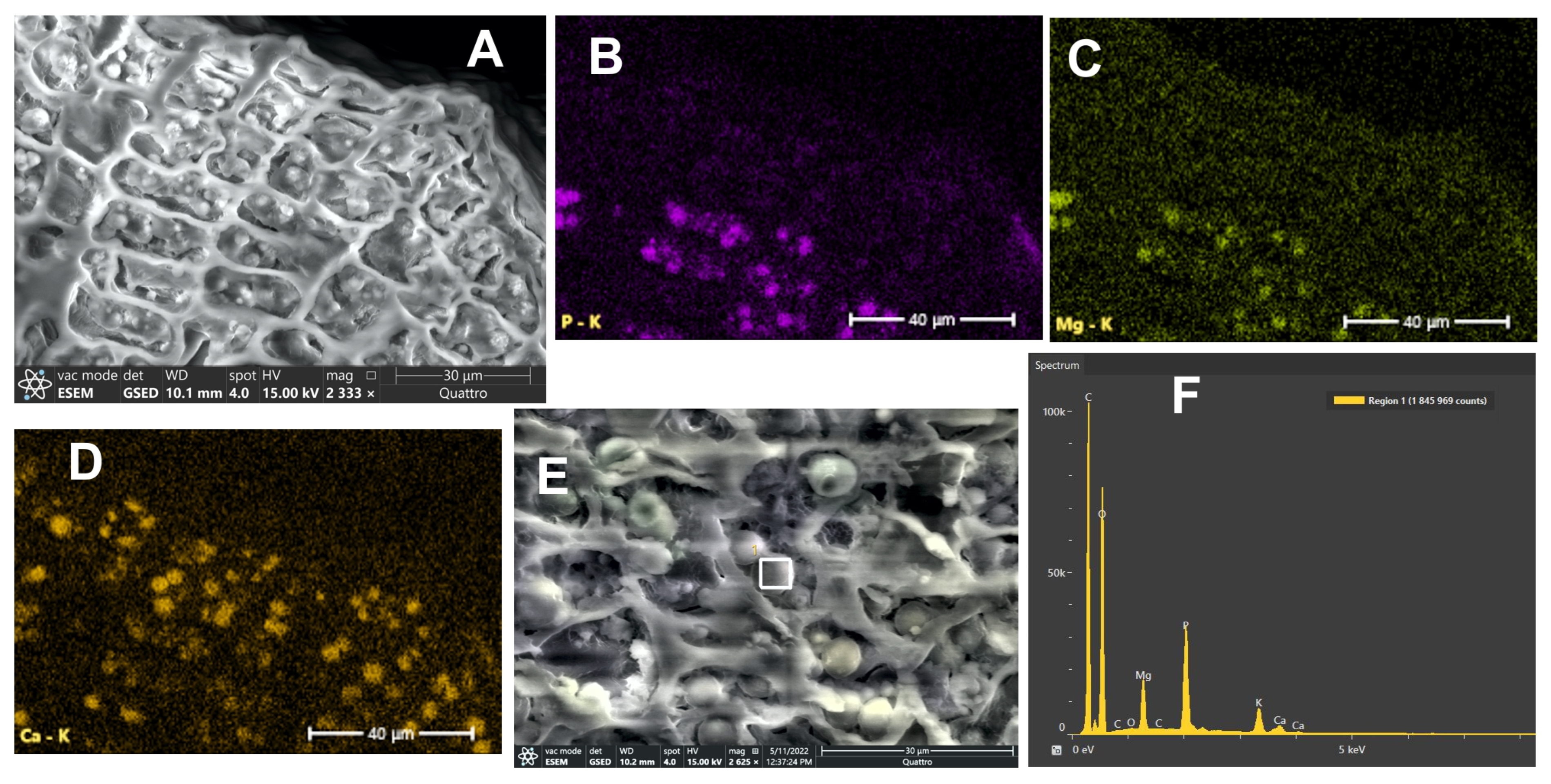
Fruits of two elliptical mericarps, pubescent; 2.5–4 mm long, 1.3–1.6 mm broad; primary ribs equal, keeled. The fruit surface is densely covered with short conical papillae and long hairs (150–400 μm). Mericarps not compressed dorsally; commissure of intermediate width; mesocarp parenchyma non-lignified; mesocarp cells above vallecular and commissural secretory ducts are relatively large, square, not compressed; secretory ducts solitary in each furrow, two on the commissural side; rib secretory ducts obsolete.
According to polarizing microscopy, crystals are located in the exocarp, mesocarp, and on commissural side.
ESEM and EDS. Eight maps and eight spectra were studied. On the elemental maps, calcium accumulations are shown in the exocarp, and parenchyma of the mesocarp; in the sclerenchyma of the vascular bundle, calcium inclusions were not detected. There is little calcium in the endosperm, it is noticeable in cells where spherical bodies with phosphorus and magnesium are not visible. Accumulations of calcium (3–10 μm in size) in the exocarp and mesocarp can fill almost the entire cell cavity, they are composed of carbon, oxygen, and calcium; the calcium content being from 15.4 to 19.6%. In the sclerenchyma of the vascular bundle, 4.2% calcium was determined, and in the top of the hair 3.9%.
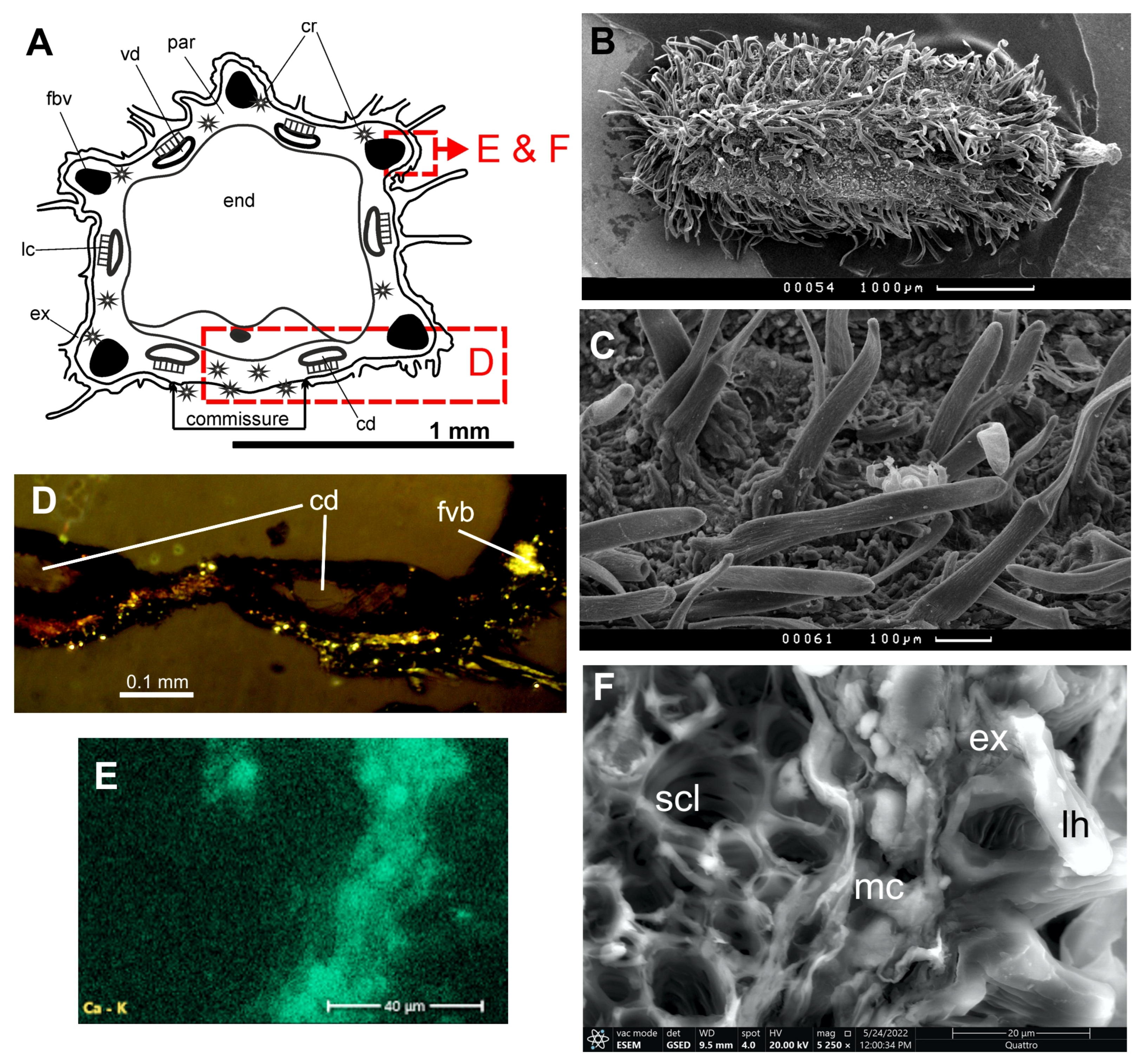
Modesciadium involucratum (Maire) P. Vargas and Jim. Mejías (
Figure 5).
Fruits of two ovate or almost spherical mericarps; 1–1.1 mm long, 0.6–0.8 mm broad; primary ribs equal, keeled; commissure of intermediate width. On the fruit surface, there are thick (20–25 μm diam.) short (20–35 μm) blunt hairs with rugulate cuticles. Mericarps not compressed dorsally, mesocarp parenchyma non-lignified; mesocarp cells above vallecular and commissural secretory ducts are relatively large, square, not compressed; vallecular ducts solitary in each furrow, two on the commissural side; rib secretory ducts obsolete.
The polarizing microscope shows crystals in exocarp, the outer layers of mesocarp, and a few crystals on the commissure.
ESEM and EDS: Nine maps and 14 spectra were studied. Small (3–5 µm) druses of crystals and irregularly shaped bodies 1–2 µm in size are located in the exocarp and mesocarp. In the endosperm, some cells are completely filled with calcium compounds up to 12 microns in size. The crystals are composed of carbon, oxygen, and calcium. The content of calcium on the elemental maps of the fruit transverse section was from 2.1% to 4.7%, and in the thick layer of non-cellular substance on the commissure from 13.3% to 15.9%. In the spectra of the elements, calcium content on the crystals was shown from 9.6% to 29.8%, in the sclerenchyma 4.9 to 7.7%, and in the parenchyma without obvious crystals, the calcium content was 5.4%.
Rutheopsis tortuosa (Webb and Berthel.) Frank., Reyes-Bet., Reduron and Spalik. (
Figure 6).
Fruits with ovate or oblong mericarps; 2–2.5 mm long, 1.3–1.4 mm broad, primary ribs equal, narrowly winged. Mericarps in the transverse section are not compressed dorsally; exocarp cells are small, commissure intermediate or broad; mesocarp consists mainly of non-lignified parenchyma cells and lignified parenchyma cells with pitted walls at the base of ribs; vallecular secretory ducts solitary, two on the commissural side; rib secretory ducts obsolete; crystals are present in exocarp and mesocarp, and few crystals on commissure. The fruit surface is densely covered with conical papillae, the cuticle being striato-knotted, similar to Ammi majus.
The polarized microscopy. On the transverse section, there are many crystals in the exocarp and outer layers of the mesocarp, and many in the endosperm. The exocarp contains crystals in almost every cell (
Figure 6C).
ESEM and EDS. Four maps and three spectra were studied. On the elemental maps, calcium is detected in the exocarp and mesocarp parenchyma. Some cells are almost completely filled with calcium compounds up to 15 µm in size, in other cells, small crystals of 1–2 µm are visible, which apparently appeared during the cutting of the material or lay in the cells initially. Small druses 2–3 μm in diam. are rare. The crystals are composed of carbon, oxygen, and calcium. The calcium content in crystal clusters is 17.6–30.0%. The sclerenchyma of the vascular bundle contains 4.1% calcium.
In all the studied species, the endosperm contains numerous spherical bodies 2–3 μm in size with a high content of phosphorus and magnesium and a low content of calcium (
Figure 7). On spectra and maps of all studied species, the silicon content is low (<0.3%), and it has no pronounced peaks on the diagrams (
Figure 3F,H and
Figure 7F).
4. Discussion
Angyalossy et al. (2016) mentioned that “It is often assumed that plant crystals are always composed of calcium oxalate, but … many plant crystals assumed to be Ca oxalate have never been positively identified as such” (p. 587). We have studied the elemental composition of crystals in the fruits of six species of the Apiaceae family (Sanicula rubriflora, Chaerophylum bulbosum, Ammi majus, Deverra aphylla, Modesciadium involucratum, and Rutheopis tortuosa) for the first time. The revealed crystals contain carbon, oxygen, and a large amount of calcium (10–30%) and are most likely oxalate. The crystals do not contain significant amounts of magnesium, phosphorus, sulfur, silicon, and other elements. The content of calcium in the surrounding parenchymal cells that did not contain crystals (5.4–9.2%) is lower than in the crystals, but nevertheless quite high. Sclerenchyma contains 0.8–6.7% calcium, and endosperm contains 0.3–8.7%. The calcium content in similar structures varies considerably even within the same fruit. The content of silicon in the fruits is low (<0.3%), this element does not form significant inclusions in the cells. The method of energy-dispersive analysis is useful at a qualitative level, and one should keep in mind the variability of indicators in biological objects.
We confirmed the presence of numerous calcium crystals in genera previously studied by other authors. The fruits of
Sanicula rubriflora contain large druses on the commissure and in the mesocarp. In the fruits of
Chaerophyllum bulbosum, large spherical clusters of crystals are located in the commissure region and are absent in other parts of the pericarp. SEM studies have shown that the crystals lie on the surface of the commissure, and the cell walls in this area are destroyed. Crystals in the endosperm of the seed are found in most Apiaceae and are of no importance for taxonomy [
8].
In all four studied species of the clade Apieae—
Ammi majus,
Deverra aphylla,
Modesciadium involucratum, and
Rutheopsis tortuosa—small crystals were found in the fruits in the exocarp, on the commissure, and often in the mesocarp parenchyma. Although the presence of crystals in
Ammi majus has been known for a long time [
12], no further searches for crystals were carried out, and their significance for taxonomy was not discussed. Pimenov and Pogorelova [
10] use the presence of crystals only as one of the characters distinguishing the genera
Ammi and
Visnaga. J.Rompel [
8] discovered that large druses and spheric crystal aggregates are of great taxonomic value at the level of subfamilies and tribes; probably, the taxonomic importance of the small crystals in the pericarp of Apiaceae was not so obvious (although he studied
Ammi majus) and therefore he did not discuss them in his publication. Thus, the character of the presence of small crystals in the exocarp of Apiaceae has not been sufficiently studied, and its taxonomic significance has yet to be determined.
Numerous hypotheses have been proposed regarding the function of oxalate crystals; strong evidence exists for the regulation of calcium metabolism, plant defense against herbivores, and metal detoxification [
26]. Crystals in the stems of Diospyros may support the electrolyte homeostasis in the plant, according to the authors [
27]. The association of crystals with cell death during tomato anther dehiscence was noted [
28] and in the process of tomato seed coat development [
29]. Crystals have also been found in anthers of
Cordia crenata and
C. sinensis (Boraginaceae) in the dehiscence zone [
30].
Calcium oxalate druses are involved in the defense response of the ornamental tree
Prunus serrulata to fungal infection [
31]. An abscission layer with a large number of crystals forms around the infected area of the leaf, the cells of this layer die and are destroyed, and the infected area falls out of the leaf, leaving a small hole (shot hole disease). Before the falling flowers of Hibiscus, a separating layer containing numerous druses of calcium oxalate is formed [
32]. Separation occurs due to the degradation of the middle lamella, the protoplast being intact. The authors suppose that calcium is released during the hydrolysis of pectin and accelerate the formation of druses.
In the carrot family, fruits split into two mercarps, which is important for seed dispersal in natural habitats. The carrot family contains many economically important species: vegetables, aromatic, and medicinal plants. Some wild species of the genera
Bunium L. and
Chaerophyllum L. produce edible tubers [
11]. Fruit dehiscence must be taken into account when agricultural plants breed. On the one hand, it is necessary to prevent crop loss due to seed fall, and on the other hand, individual mericarps containing single embryos are needed for sowing. The splitting of fruits into mericarps is associated with cell separation, and probably with cell death and the destruction of their walls. In fruits with
Sanicula-type and
Scandix-type crystals, a significant number of large calcium oxalate crystals are observed in the area of separation of the mericarps (commissure).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






