
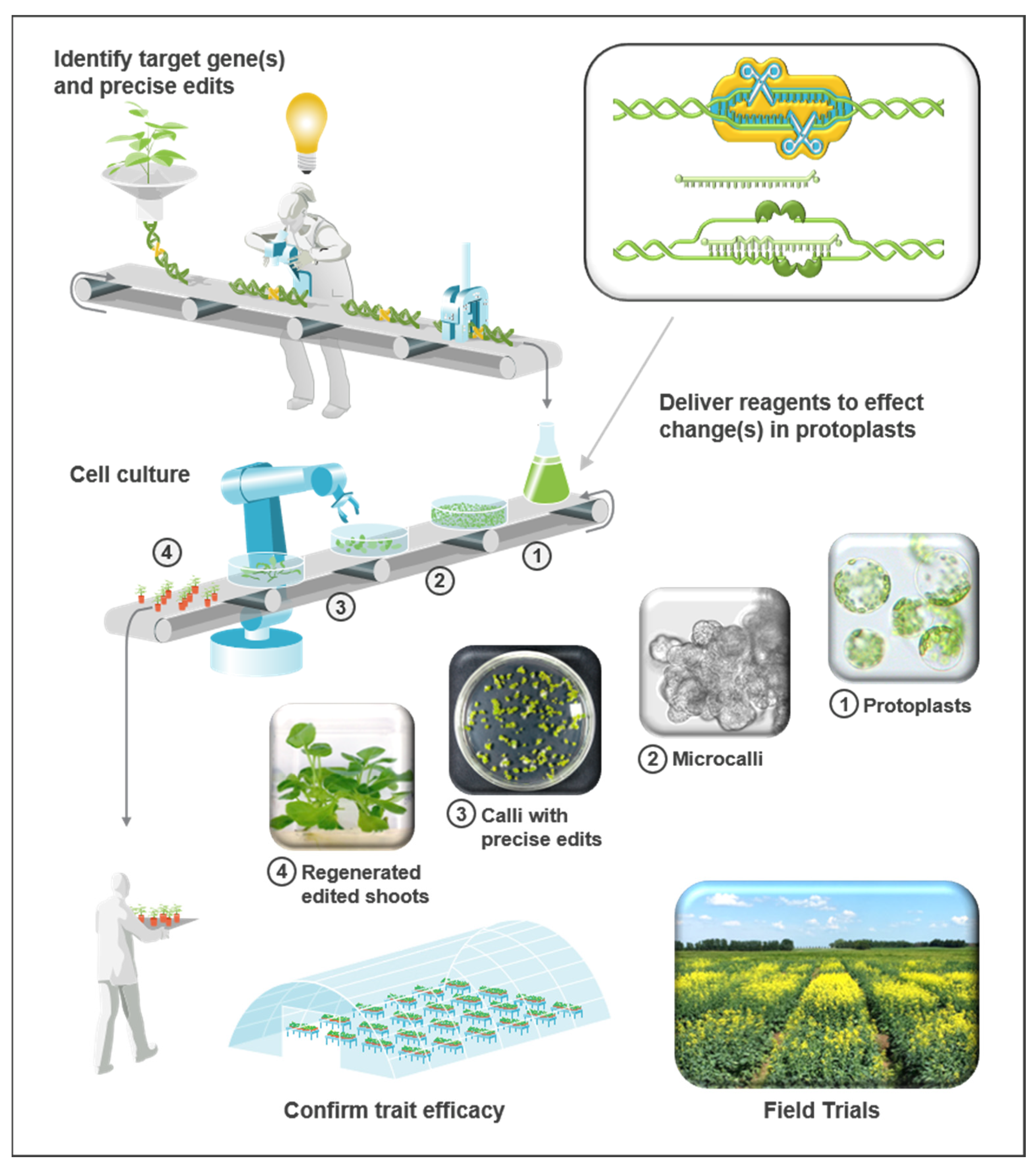
Industrial Scale Gene Editing in Brassica napus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Gene-Edited Plants
2.2. Genotype Identification by NGS
2.3. Off-Target Analysis
2.4. Characterization of Plants with BnaFAD2 LOF Mutations
2.5. Random Impact Test (RIT) for Pod Shatter
3. Results
3.1. Single Cell Approach—High Oleic Oil
3.2. Single Cell Approach—Pod Shatter Reduction
4. Discussion
4.1. Modified Oil Profiles Using Gene Editing
4.2. Canola with Pods Displaying Pod Shatter Reduction
5. Conclusions
5.1. Industrialized Gene Editing (IGE)
5.2. Multiplex/Multitrait Gene Editing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beetham, P.R.; Kipp, P.B.; Sawycky, X.L.; Arntzen, C.J.; May, G.D. A tool for functional plant genomics: Chimeric RNA/DNA oligonucleotides cause in vivo gene-specific mutations. Proc. Natl. Acad. Sci. USA 1999, 96, 8774–8778. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Gao, C. Targeted genome modification technologies and their applications in crop improvements. Plant Cell Rep. 2014, 33, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J. Applying gene editing to tailor precise genetic modifications in plants. J. Biol. Chem. 2020, 295, 13267–13276. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Saleem, F.; Kanwal, M.; Mustafa, G.; Yousaf, S.; Arshad, H.M.I.; Hameed, M.K.; Khan, M.S.; Joyia, F.A. Modern Trends in plant genome editing: An inclusive review of the CRISPR/Cas9 toolbox. Int. J. Mol. Sci. 2019, 20, 4045. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Hsu, C.T.; Yang, L.H.; Lee, L.Y.; Fu, J.Y.; Cheng, Q.W.; Wu, F.H.; Hsiao, H.C.; Zhang, Y.; Zhang, R.; et al. Application of protoplast technology to CRISPR/Cas9 mutagenesis: From single-cell mutation detection to mutant plant regeneration. Plant Biotechnol. J. 2018, 16, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef]
- Schiml, S.; Fauser, F.; Puchta, H. The CRISPR/Cas system can be used as nuclease for in planta gene targeting and as paired nickases for directed mutagenesis in Arabidopsis resulting in heritable progeny. Plant J. 2014, 80, 1139–1150. [Google Scholar] [CrossRef]
- Xu, R.; Li, H.; Qin, R.; Wang, L.; Li, L.; Wei, P.; Yang, J. Gene targeting using the Agrobacterium tumefaciens-mediated CRISPR-Cas system in rice. Rice 2014, 7, 5. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef]
- Mikami, M.; Toki, S.; Endo, M. Comparison of CRISPR/Cas9 expression constructs for efficient targeted mutagenesis in rice. Plant Mol. Biol. 2015, 88, 561–572. [Google Scholar] [CrossRef]
- Mikami, M.; Toki, S.; Endo, M. Parameters affecting frequency of CRISPR/Cas9 mediated targeted mutagenesis in rice. Plant Cell Rep. 2015, 34, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Tanaka, W.; Mikami, M.; Endo, M.; Hirano, H.Y. Generation of artificial drooping leaf mutants by CRISPR-Cas9 technology in rice. Genes Genet. Syst. 2016, 90, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhang, H.; Lou, D.; Yu, D. Selection of highly efficient sgRNAs for CRISPR/Cas9-based plant genome editing. Sci. Rep. 2016, 6, 21451. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef] [PubMed]
- Gocal, G.F.W. Gene editing in Brassica napus for basic research and trait development. Vitr. Cell. Dev. Biol.-Plant 2021, 9, 731–748. [Google Scholar] [CrossRef]
- Sauer, N.J.; Narváez-Vásquez, J.; Mozoruk, J.; Miller, R.B.; Warburg, Z.J.; Woodward, M.J.; Mihiret, Y.A.; Lincoln, T.A.; Segami, R.E.; Sanders, S.L.; et al. Oligonucleotide-mediated genome editing provides precision and function to engineered nucleases and antibiotics in plants. Plant Physiol. 2016, 170, 1917–1928. [Google Scholar] [CrossRef]
- Yang, H.; Wu, J.J.; Tang, T.; Liu, K.D.; Dai, C. CRISPR/Cas9-mediated genome editing efficiently creates specific mutations at multiple loci using one sgRNA in Brassica napus. Sci. Rep. 2017, 7, 7489. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, K.; Li, H.; Han, S.; Meng, Q.; Khan, S.U.; Fan, C.; Xie, K.; Zhou, Y. Precise editing of CLAVATA genes in Brassica napus L. regulates multilocular silique development. Plant Biotechnol. J. 2018, 16, 1322–1335. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N. DNA-free genome editing methods for targeted crop improvement. Plant Cell Rep. 2016, 35, 1469–1474. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.G.; Kim, S.T.; Choe, S.; Kim, J.S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, S.T.; Ryu, J.; Kang, B.C.; Kim, J.S.; Kim, S.G. CRISPR/Cpf1-mediated DNA-free plant genome editing. Nat. Commun. 2017, 8, 14406. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef] [PubMed]
- Haun, W.; Coffman, A.; Clasen, B.M.; Demorest, Z.L.; Lowy, A.; Ray, E.; Retterath, A.; Stoddard, T.; Juillerat, A.; Cedrone, F.; et al. Improved soybean oil quality by targeted mutagenesis of the fatty acid desaturase 2 gene family. Plant Biotechnol. J. 2014, 12, 934–940. [Google Scholar] [CrossRef]
- Clasen, B.M.; Stoddard, T.J.; Luo, S.; Demorest, Z.L.; Li, J.; Cedrone, F.; Tibebu, R.; Davison, S.; Ray, E.E.; Daulhac, A.; et al. Improving cold storage and processing traits in potato through targeted gene knockout. Plant Biotechnol. J. 2016, 14, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhu, J.; Liu, H.L. Protoplasts: A useful research system for plant cell biology, especially dedifferentiation. Protoplasma 2013, 250, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Dlugosz, E.M.; Lenaghan, S.C.; Stewart, C.N., Jr. A robotic platform for high-throughput protoplast isolation and transformation. J. Vis. Exp. 2016, 115, e54300. [Google Scholar] [CrossRef]
- Sauer, N.J.; Mozoruk, J.; Miller, R.B.; Warburg, Z.J.; Walker, K.A.; Beetham, P.R.; Schöpke, C.R.; Gocal, G.F.W. Oligonucleotide-directed mutagenesis for precision gene editing. Plant Biotechnol. J. 2016, 14, 496–502. [Google Scholar] [CrossRef]
- Baltes, N.J.; Gil-Humanes, J.; Voytas, D.F. Genome engineering and agriculture: Opportunities and challenges. In Gene Editing in Plants; Weeks, D.P., Yang, B., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 149, pp. 1–26. [Google Scholar] [CrossRef]
- Liang, Z.; Zhang, K.; Chen, K.; Gao, C. Targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. J. Genet. Genom. 2014, 41, 63–68. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted mutagenesis; precise gene editing; and site-specific gene insertion in maize using Cas9 and guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.L.; Wang, D.; Gao, C. Precise base editing in rice; wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.S.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiol. Plant. 2018, 164, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Kartha, K.K.; Michayluk, M.R.; Kao, K.N.; Gamborg, O.L.; Constabel, F. Callus formation and plant regeneration from mesophyll protoplasts of rape plants (Brassica napus L. cv. Zephyr). Plant Sci. 1974, 3, 265–271. [Google Scholar] [CrossRef]
- Gocal, G.F.W.; Schöpke, C.; Beetham, P.R. Oligo-mediated targeted gene editing. In Advances in New Technology for Targeted Modification of Plant Genomes; Zhang, F., Puchta, H., Thomson, J.G., Eds.; Springer: New York, NY, USA, 2015; pp. 73–90. [Google Scholar] [CrossRef]
- Wang, B.; Guo, X.; Wang, C.; Ma, J.; Niu, F.; Zhang, H.; Yang, B.; Liang, W.; Han, F.; Jiang, Y.Q. Identification and characterization of plant-specific NAC gene family in canola (Brassica napus L.) reveal novel members involved in cell death. Plant Mol. Biol. 2015, 87, 395–411. [Google Scholar] [CrossRef]
- Pelletier, G.; Primard, C.; Vedel, F.; Chetrit, P.; Remy, R.; Rouselle, P.; Renard, M. Intergeneric cytoplasm hybridization in Cruciferae by protoplast fusion. Mol. Gen. Genet. 1983, 191, 244–250. [Google Scholar] [CrossRef]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef]
- Hu, X.; Sullivan-Gilbert, M.; Gupta, M.; Thompson, S.A. Mapping of the loci controlling oleic and linolenic acid contents and development of fad2 and fad3 allele-specific markers in canola (Brassica napus L.). Theor. Appl. Genet. 2006, 113, 497–507. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, C.; Gua, Z.; Qin, J.; Wu, J.; Li, Q.; Fu, T.; Zhou, Y. Identification of FAD2 and FAD3 genes in Brassica napus genome and development of allele-specific markers for high oleic and low linoleic acid contents. Theor. Appl. Genet. 2012, 125, 715–729. [Google Scholar] [CrossRef]
- Lee, K.; Sohn, I.S.; Jung, J.H.; Kim, S.H.; Roh, K.H.; Kim, J.; Suh, M.C.; Kim, H.U. Functional analysis and tissue-differential expression of four FAD2 genes in amphidiploid Brassica napus derived from Brassica rapa and Brassica oleracea. Gene 2013, 531, 253–262. [Google Scholar] [CrossRef]
- Well, R.; Trick, M.; Soumpourou, E.; Clissold, L.; Morgan, C.; Bancroft, I. The control of seed oil polyunsaturate content in the polyploid crop species Brassica napus. Mol. Breed. 2014, 33, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Engelen, S.; Denolf, P.; Wallis, J.G.; Lynch, K.; Bengtsson, J.D.; Van Thournout, M.; Haesendonckx, B.; Browse, J. Identification, characterization and field testing of Brassica napus mutants producing high-oleic oils. Plant J. 2019, 98, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Cui, T.; Zhang, L.; Yang, Q.; Yang, Y.; Xie, K.; Fan, C.; Zhou, Y. Modifications of fatty acid profile through targeted mutation at BnaFAD2 gene with CRISPR/Cas9-mediated gene editing in Brassica napus. Theor. Appl. Genet. 2020, 133, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Gocal, G.F.W.; Knuth, M. Fad2 Genes and Mutations. PCT International Publication Number WO 2019/195611A1, 4 April 2019. [Google Scholar]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Kadkol, G.P.; Beliharz, V.C.; Halloran, G.M.; MacMillan, R.H. Anatomical basis of shatter-resistance in the oilseed Brassica. Aust. J. Bot. 1986, 34, 595–601. [Google Scholar] [CrossRef]
- Wang, R.; Ripley, V.L.; Rakow, G. Pod shatter resistance evaluation in cultivars and breeding lines of Brassica napus, B. juncea and Sinapis alba. Plant Breed. 2007, 126, 588–595. [Google Scholar] [CrossRef]
- Gocal, G.F.W. Shatterproof Genes and Mutations. U.S. Patent 11,359,208B2, 9 January 2019. [Google Scholar]
- Zaman, Q.U.; Wen, C.; Yuqin, S.; Mengyu, H.; Desheng, M.; Jacqueline, B.; Baohong, Z.; Chao, L.; Qiong, H. Characterization of SHATTERPROOF Homoeologs and CRISPR-Cas9-Mediated Genome Editing Enhances Pod-Shattering Resistance in Brassica napus L. CRISPR J. 2021, 4, 360–370. [Google Scholar] [CrossRef]
- Laga, B.; den Boer, B.; Lambert, B. 2008 Brassica Plant Comprising a Mutant Indehiscent Allele. U.S. Patent 9,475,849B2, 25 November 2008. [Google Scholar]
- Helm, J. Morphologisch-taxonomische Gliederung der Kultursippen von Brassica oleracea. Kulturpflanze 1963, 11, 92–210. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Number of LOF Genes | LOF Genotype Number | Targeted Genes * | Number of Shoots with Precise Edits | |
|---|---|---|---|---|
| Line A | Line B | |||
| 1 | 1 | 1 | 130 | 71 |
| 2 | 2 | 106 | 67 | |
| 3 | 3 | 161 | 174 | |
| 4 | 4 | 118 | 96 | |
| 2 | 5 | 1 + 2 | 51 | 6 |
| 6 | 1 + 3 | 62 | 19 | |
| 7 | 1 + 4 | 34 | 8 | |
| 8 | 2 + 3 | 58 | 21 | |
| 9 | 2 + 4 | 37 | 8 | |
| 10 | 3 + 4 | 121 | 41 | |
| 3 | 11 | 1 + 2 + 3 | 29 | 2 |
| 12 | 1 + 2 + 4 | 12 | 0 | |
| 13 | 1 + 3 + 4 | 91 | 13 | |
| 14 | 2 + 3 + 4 | 68 | 1 | |
| 4 | 15 | 1 + 2 + 3 + 4 | 51 | 1 |
| Genotype | Number of Edited FAD2 Genes | Oleic (18:1) | Linoleic (18:2) | Linolenic (18:3) |
|---|---|---|---|---|
| Mole% | Mole% | Mole% | ||
| Wildtype (n = 1) | 0 | 61.13 ± 0.00 a | 20.18 ± 0.00 | 12.34 ± 0.00 a |
| BnaA05.FAD2a; BnaC05.FAD2a (n = 3) | 2 | 82.73 ± 3.92 b | 3.45 ± 0.79 | 7.35 ± 2.36 b |
| BnaC05.FAD2a; BnaC01.FAD2b (n = 1) | 2 | 69.51 ± 0.00 a | 13.64 ± 0.00 | 10.52 ± 0.00 ab |
| BnaA05.FAD2a; BnaC05.FAD2a; BnaC01.FAD2b(n = 3) | 3 | 88.42 ± 0.50 c | 1.89 ± 0.05 a | 3.59 ± 0.32 c |
| BnaA05.FAD2a; BnaC05.FAD2a; BnaA01.FAD2b BnaC01.FAD2b (n = 2) | 4 | 88.52 ± 0.22 bc | 1.89 ± 0.02 a | 3.47 ± 0.01 c |
| Number of LOF Genes | Number of Targeted Genotypes | Number of Shoots with Precise Edits * | |||
|---|---|---|---|---|---|
| Line A | Line B | Line C | Line D | ||
| 1 | 8 | 296 | 8 | 145 | 153 |
| 2 | 28 | 234 | 16 | 71 | 87 |
| 3 | 56 | 203 | 23 | 50 | 73 |
| 4 | 70 | 154 | 11 | 15 | 26 |
| 5 | 56 | 114 | 12 | 13 | 25 |
| 6 | 28 | 81 | 10 | 8 | 18 |
| 7 | 8 | 39 | 2 | 10 | 12 |
| 8 | 1 | 7 | 1 | 1 | 2 |
| Number of shoots screened: | 5395 | 1490 | 2620 | 4620 | |
| Line | Greenhouse | Field | ||
|---|---|---|---|---|
| N | % Additional BF | N | % Additional BF | |
| A_01 | 3 | 44.44 a | 4 | 48.43 a |
| A_02 | 1 | 42.85 ab | 4 | 51.09 a |
| A_03 | 2 | 35.71 abc | 4 | 37.11 a |
| A_04 | 2 | 30.95 abcd | 4 | 38.80 a |
| A_05 | 3 | 25.55 bcd | 4 | 43.76 a |
| A_06 | 3 | 23.65 cd | 4 | 26.67 a |
| A_07 | 3 | 18.88 cd | 4 | 33.98 a |
| A_08 | 3 | 16.34 de | 4 | 38.63 a |
| A_WT | 3 | 0.00 e | 4 | 0.00 b |
| LSD (0.05) | (3,3) | 17.4 | (4,4) | 25.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, A.; Narváez-Vásquez, J.; Mozoruk, J.; Niu, Z.; Luginbühl, P.; Sanders, S.; Schöpke, C.; Sauer, N.; Radtke, J.; Gocal, G.; et al. Industrial Scale Gene Editing in Brassica napus. Int. J. Plant Biol. 2023, 14, 1064-1077. https://doi.org/10.3390/ijpb14040077
Walker A, Narváez-Vásquez J, Mozoruk J, Niu Z, Luginbühl P, Sanders S, Schöpke C, Sauer N, Radtke J, Gocal G, et al. Industrial Scale Gene Editing in Brassica napus. International Journal of Plant Biology. 2023; 14(4):1064-1077. https://doi.org/10.3390/ijpb14040077
Chicago/Turabian StyleWalker, Andrew, Javier Narváez-Vásquez, Jerry Mozoruk, Zhixia Niu, Peter Luginbühl, Steve Sanders, Christian Schöpke, Noel Sauer, Jim Radtke, Greg Gocal, and et al. 2023. "Industrial Scale Gene Editing in Brassica napus" International Journal of Plant Biology 14, no. 4: 1064-1077. https://doi.org/10.3390/ijpb14040077
APA StyleWalker, A., Narváez-Vásquez, J., Mozoruk, J., Niu, Z., Luginbühl, P., Sanders, S., Schöpke, C., Sauer, N., Radtke, J., Gocal, G., & Beetham, P. (2023). Industrial Scale Gene Editing in Brassica napus. International Journal of Plant Biology, 14(4), 1064-1077. https://doi.org/10.3390/ijpb14040077