

Blue Carbon in Mangroves of the Arid Zones of San Ignacio and El Dátil Lagoons, El Vizcaino Biosphere Reserve, Baja California, Mexico

 and
and
Abstract
:1. Introduction
2. Materials and Methods
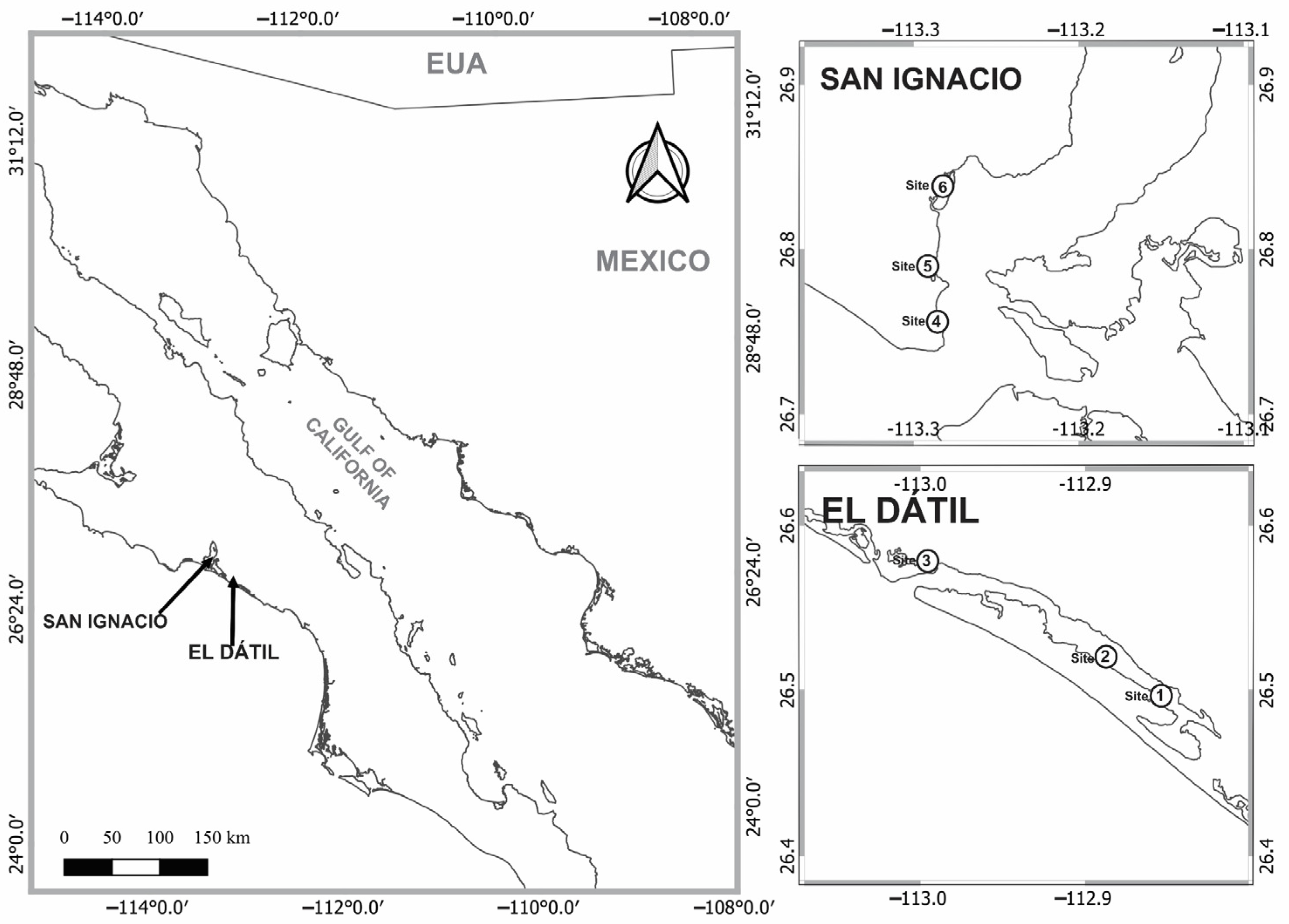
2.1. Study Site
2.2. Physico-Chemical Characteristics of Water and Sediment
2.3. Mangrove Forest Structure
2.4. Primary Productivity Based on Above-Ground Leaf Litter
2.5. Below-Ground Root Biomass (BRB)
2.6. Biomass-Related Carbon
2.7. Carbon in Soil
2.8. AGC/BGC Ratio
2.9. Statistical Analysis
3. Results
3.1. Water Physico-Chemical Characteristics
3.2. Sediment Physico-Chemical Characteristics
3.3. Mangrove Structural Attributes
3.4. Soil Leaf Litter
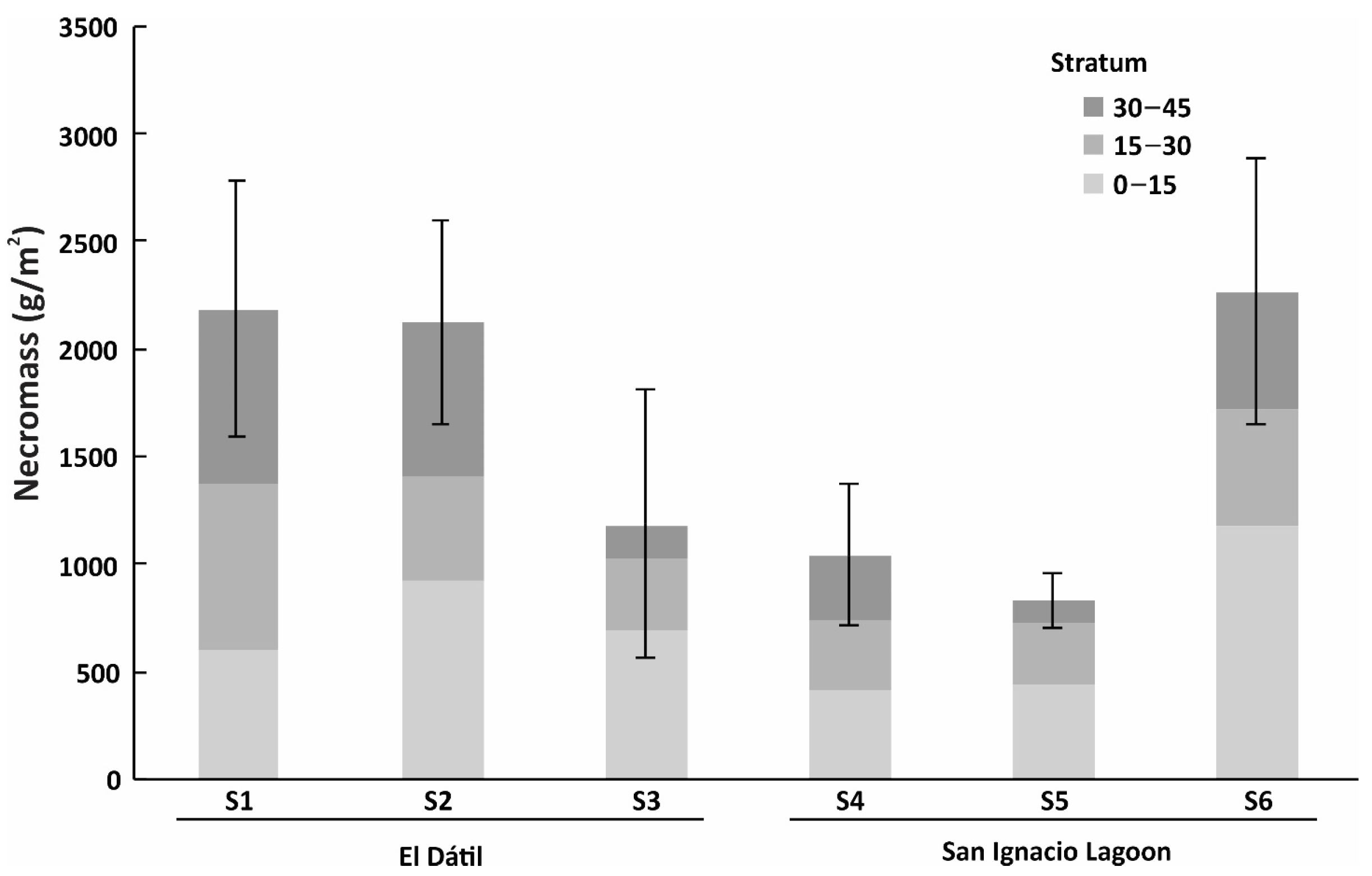
3.5. Below-Ground Root Biomass
3.6. Carbon Stock Related to Mangrove Biomass
3.7. Carbon in the Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; p. 1535. Available online: https://reliefweb.int/report/world/climate-change-2021-physical-science-basis?gclid=CjwKCAjw_aemBhBLEiwAT98FMjxUZr_pmpK8C-rVg3xi3xAj2FZa2Eeh6RsAnFdFezGlI_Om__tSxBoCK4AQAvD_BwE (accessed on 16 November 2023).
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef]
- Bulmer, R.H.; Stephenson, F.; Jones, H.F.E.; Townsend, M.; Hillman, J.R.; Schwendenmann, L.; Lundquist, C.J. Blue Carbon Stocks and Cross-Habitat Subsidies. Front. Mar. Sci. 2020, 7, 380. [Google Scholar] [CrossRef]
- McLeod, K.W.; Chmura, G.L.; Bouillon, S.; Salm, R.; Bjork, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal hábitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Fourqurean, J.W.; Duarte, C.M.; Kennedy, H.; Marbà, N.; Holmer, M.; Mateo, M.A.; Apostolaki, E.T.; Kendrick, G.A.; Krause-Jensen, D.; McGlathery, K.J. Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 2012, 5, 505–509. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marbà, N.; et al. Estimating global blue carbon emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef]
- Banerjee, K.; Sahoo, C.K.; Bal, G.; Mallik, K.; Paul, R.; Mitra, A. High blue carbon stock in mangrove forests of Eastern India. Trop. Ecol. 2019, 61, 150–167. [Google Scholar] [CrossRef]
- Adame, M.F.; Cherian, S.; Reef, R.; Stewart-Koster, B. Mangrove root biomass and the uncertainty of belowground carbon estimations. For. Ecol. Manag. 2017, 403, 52–60. [Google Scholar] [CrossRef]
- Torres, J.R.; Sanchez-Mejia, Z.M.; Arreola-Lizárraga, J.A.; Yépez, E.A.; Reynaga-Franco, F.D.; Choix, F.J. Root biomass and productivity in subtropical arid mangroves from the Gulf of California. Rhizosphere 2021, 18, 100356. [Google Scholar] [CrossRef]
- Osland, M.J.; Day, R.H.; Larriviere, J.C. Aboveground allometric models for freeze-affected black mangroves (Avicennia germinans): Equations for a climate sensitive mangrove-marsh ecotone. PLoS ONE 2014, 9, e99604. [Google Scholar] [CrossRef] [PubMed]
- Comley, B.W.T.; McGuinness, K.A. Above- and below-ground biomass, and allometry, of four common northern Australian mangroves. Aust. J. Bot. 2005, 53, 431–436. [Google Scholar] [CrossRef]
- Mitra, A.; Sengupta, K.; Banerjee, K. Standing biomass and carbon storage of above-ground structures in dominant mangrove trees in the Sundarbans. For. Ecol. Manag. 2011, 261, 1325–1335. [Google Scholar] [CrossRef]
- Njana, M.A.; Eid, T.; Zahabu, E.; Malimbwi, R. Procedures for quantification of belowground biomass of three mangrove tree species. Wetl. Ecol. Manag. 2015, 23, 749–764. [Google Scholar] [CrossRef]
- Virgulino-Júnior, P.C.C.; Carneiro, D.N.; Nascimento, W.R., Jr.; Cougo, M.F.; Fernandes, M.E.B. Biomass and carbon estimation for scrub mangrove forests and examination of their allometric associated uncertainties. PLoS ONE 2020, 15, e0230008. [Google Scholar] [CrossRef] [PubMed]
- Matsui, N. Estimated stocks of organic carbon in mangrove roots and sediments in Hinchinbrook Channel, Australia. Mangroves Salt Marshes 1998, 2, 199–204. [Google Scholar] [CrossRef]
- Torres, J.R.; Barba, E.; Choix, F.J. Production and biomass of mangrove roots in relation to hydroperiod and physico-chemical properties of sediment and water in the Mecoacan Lagoon, Gulf of Mexico. Wetl. Ecol. Manag. 2019, 27, 427–442. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Stanton, D.E.; Schmitz, N.; Farquhar, G.D.; Ball, M.C. Growth responses of the mangrove Avicennia marina to salinity: Development and function of shoot hydraulic systems require saline conditions. Ann. Bot. 2015, 115, 397–407. [Google Scholar] [CrossRef]
- Alongi, D.M. Global Significance of Mangrove Blue Carbon in Climate Change Mitigation. Sci 2020, 2, 67. [Google Scholar] [CrossRef]
- Herrera-Silveira, J.A.; Pech-Cardenas, M.A.; Morales-Ojeda, S.M.; Cinco-Castro, S.; Camacho-Rico, A.; Caamal Sosa, J.P.; Mendoza-Martinez, J.E.; Pech-Poot, E.Y.; Montero, J.; Teutli-Hernandez, C. Blue carbon of Mexico, carbon stocks and fluxes: A systematic review. PeerJ 2020, 8, e8790. [Google Scholar] [CrossRef]
- Félix-Pico, E.; Serviere-Zaragoza, E.; Riosmena-Rodriguez, R.; León-DeLaCruz, J. Los Manglares de la Península de Baja California. Interdisciplinary Center for Marine Sciences, Biological Research Center of the Northwest S.C. and the Autonomous University of Baja California. 2011. Available online: http://dspace.cibnor.mx:8080/handle/123456789/1400 (accessed on 16 November 2023).
- CONABIO. Sección Mexicana del Consejo Internacional para la Preservación de las Aves CIPAMEX- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Áreas de Importancia para la Conservación de las Aves. Financiado por CONABIO-FMCN-CCA. 1999. Available online: http://conabioweb.conabio.gob.mx/aicas/doctos/aicas.html (accessed on 16 November 2023).
- DOF. Decreto Por el Que se Declara la Reserva de la Biosfera “El Vizcaíno”, Ubicada en el Municipio de Mulegé, B.C.S. 1988. Available online: https://dof.gob.mx/nota_detalle.php?codigo=4794242&fecha=05/12/1988#gsc.tab=0 (accessed on 16 November 2023).
- Ruiz-Luna, A.; Acosta-Velázquez, J. Sitios de Manglar con Relevancia Biológica y Con Necesidades de Rehabilitación Ecológica, San Ignacio (Bocana-Dátil) PN07. CONABIO, México, D.F. 2009. Available online: https://bioteca.biodiversidad.gob.mx/janium/Documentos/15068.pdf (accessed on 16 November 2023).
- RAMSAR. Sistema Lagunar San Ignacio—Navachiste—Macapule, Fecha de Designación: 02-02-2004. 2008. Available online: https://rsis.ramsar.org/RISapp/files/RISrep/MX1341RIS.pdf (accessed on 16 November 2023).
- Moreno-Casasola, P.; Warner, B. Breviario Para Describir, Observar y Manejar Humedales. México: RAMSAR Instituto de Ecología A.C., CONANP, US Fish and Wildlife Service US State Department. 2009. Available online: http://www1.inecol.edu.mx/inecol/libros/Breviario_Humedales.pdf (accessed on 16 November 2023).
- Klute, A. Methods of soils analysis. Part 1. In Physical and Mineralogical Methods, 2nd ed.; EUA: Madison, WI, USA, 1986; p. 1188. [Google Scholar]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Infante, M.D. Estructura y Dinámica de las Selvas Inundables de la Planicie Costera Central del Golfo de México. Doctoral Thesis, INECOL. A. C., Jalapa, Mexico, 2011. [Google Scholar]
- Velázquez-Salazar, S.; Rodríguez-Zúñiga, M.T.; Alcántara-Maya, J.A.; Villeda-Chávez, E.; Valderrama-Landeros, L.; Troche-Souza, C.; Vázquez-Balderas, B.; Pérez-Espinosa, I.; Cruz-López, M.I.; Ressl, R.; et al. Mangroves of Mexico. Data Update and Analysis 2020. Comisión Nacional para el Uso y Conservación de la Biodiversidad. Mexico CDMX. p. 168. 2021. Available online: https://bioteca.biodiversidad.gob.mx/janium/Documentos/15638.pdf (accessed on 16 November 2023).
- Mostacedo, B.; Fredericksen, T. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal; BOLFOR: Santa Cruz, Bolivia, 2000; p. 87. Available online: https://pdf.usaid.gov/pdf_docs/PNACL893.pdf (accessed on 16 November 2023).
- Barrios-Calderón, R.J. Combustibles Forestales y su Relación con Incendios en Humedales de la Reserva de la Biósfera la Encrucijada, Chiapas. Master’s Thesis in Natural Resources and Rural Development. El Colegio de la Frontera Sur. Tapachula, Chiapas, Mex. p. 203. 2015. Available online: https://ecosur.repositorioinstitucional.mx/jspui/bitstream/1017/2003/1/100000056605_documento.pdf (accessed on 16 November 2023).
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado- Molina, C.; Ewe, S.M. Patterns of root dynamics in mangrove forests along environmental gradients in the Florida coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Smith, T.J., III; Whelan, K.R. Development of allometric relations for three mangrove species in South Florida for use in the Greater Everglades Ecosystem restoration. Wetl. Ecol. Manag. 2006, 14, 409–419. [Google Scholar] [CrossRef]
- Howard, J.; Hoyt, S.; Isensee, K.; Telszewski, M.; Pidgeon, E. (Eds.) Coastal Blue Carbon: Methods for Assessing Carbon Stocks and Emissions Factors in Mangroves, Tidal Salt Marshes, and Seagrasses; Conservation International; Intergovernmental Oceanographic Commission of UNESCO; International Union for Conservation of Nature: Arlington, VA, USA, 2014; Available online: https://www.cifor.org/knowledge/publication/5095/ (accessed on 16 November 2023).
- Kauffman, J.B.; Cummings, D.L.; Ward, D.E.; Babbitt, R. Fire in the Brazilian Amazon: 1. Biomass, nutrient pools, and losses in slashed primary forests. Oecologia 1995, 104, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; CIFOR: Bogor, Indonesia, 2012; Available online: https://www.cifor.org/publications/pdf_files/WPapers/WP86CIFOR.pdf (accessed on 16 November 2023).
- Jaramillo, V.J.; Kauffman, J.B.; Rentería-Rodríguez, L.; Cummings, D.L.; Ellingson, L.J. Biomass, carbon, and nitrogen pools in Mexican tropical dry forest landscapes. Ecosystems 2003, 6, 609–629. [Google Scholar] [CrossRef]
- Marín-Muñiz, J.L.; Hernández, M.E.; Moreno-Casasola, P. Comparing soil carbon sequestration in coastal freshwater wetlands with various geomorphic features and plant communities in Veracruz, Mexico. Plant Soil 2014, 378, 189–203. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Change Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Steel, A.; Torrie, M. Bioestadística: Principios y Procedimientos; México, D.F., Ed.; McGrawHill: New York, NY, USA, 1996. [Google Scholar]
- Lovelock, C.E.; Feller, I.C.; Ball, M.C.; Ellis, J.; Sorrell, B. Testing the Growth Rate vs. Geochemical Hypothesis for latitudinal variation in plant nutrients. Ecol. Lett. 2007, 10, 1154–1163. [Google Scholar] [CrossRef]
- Torres, J.R.; Barba, E.; Infante-Mata, D.; Sánchez, A.J. Primary productivity of the mangrove and its relationship with the population dynamics of shrimp (Decapoda: Penaeidae) in a coastal lagoon of the Gulf of Mexico. Áreas Nat. Protegidas Scr. 2022, 8, 91–107. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Adame, M.F.; Reef, R.; Santini, N.S.; Najera, E.; Turschuell, M.P.; Hayes, M.A.; Masque, P.; Lovelock, C.E. Mangroves in Arid Regions: Ecology, Threats, and Opportunities. Estuar. Coast. Shelf Sci. 2020, 248, 106796. [Google Scholar] [CrossRef]
- Ball, M.C. Patterns of secondary succession in a mangrove forest of southern Florida. Oecología 1980, 44, 226–235. [Google Scholar] [CrossRef]
- Almahasheer, H.; Serrano, O.; Duarte, C.M.; Arias-Ortiz, A.; Masque, P.; Irigoien, X. Low carbon sink capacity of Red Sea mangroves. Sci. Rep. 2017, 7, 9700. [Google Scholar] [CrossRef] [PubMed]
- Schile, L.M.; Kauffman, J.B.; Crooks, S.; Fourqurean, J.W.; Glavan, J.; Megonigal, J.P. Limits on carbon sequestration in arid blue carbon ecosystems. Ecol. Appl. 2017, 27, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, J.B.; Bhomia, R.K. Ecosystem carbon stocks of mangroves across broad environmental gradients in West-Central Africa: Global and regional comparisons. PLoS ONE 2017, 12, e0187749. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Silveira, J.A.; Camacho-Rico, A.; Pech, E.; Pech, M.; Ramírez-Ramírez, J.; Teutli-Hernández, C. Dinámica del carbono (almacenes y flujos) en manglares de México. Terra Latinoam. 2016, 34, 61–72. Available online: https://www.terralatinoamericana.org.mx/index.php/terra/article/view/76/82 (accessed on 16 November 2023).
- IPCC. IPCC Guidelines for National Greenhouse Gas Inventories—A Primer, Prepared by the National Greenhouse Gas Inventories Programme; Eggleston, H.S., Miwa, K., Srivastava, N., Tanabe, K., Eds.; IGES: Hayama, Japan, 2009. Available online: https://www.ipcc-nggip.iges.or.jp/support/Primer_2006GLs.pdf (accessed on 16 November 2023).
- Ouyang, X.; Lee, S.Y.; Connolly, R.M. The role of root decomposition in global mangrove and saltmarsh carbon budgets. Earth Sci. Rev. 2017, 166, 53–63. [Google Scholar] [CrossRef]
- Torres, J.R.; Infante-Mata, D.M.; Sánchez, A.J.; Espinoza-Tenorio, A.; Barba, E. Degradación de hojarasca y aporte de nutrientes del manglar en la Laguna Mecoacán, Golfo de México. Rev. Biol. Trop. 2018, 66, 892–907. [Google Scholar] [CrossRef]
- Wijaya, A.; Liesenberg, V.; Gloaguen, R. Retrieval of forest attributes in complex successional forests of Central Indonesia: Modeling and estimation of bitemporal data. For. Ecol. Manag. 2010, 259, 2315–2326. [Google Scholar] [CrossRef]
- Hamilton, S.; Friess, D.A. Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat. Clim. Chang. 2018, 8, 240–244. [Google Scholar] [CrossRef]
- Senger, D.F.; Saavedra-Hortua, D.A.; Engel, S.; Schnurawa, M.; Moosdorf, N.; Gillis, L.G. Impacts of wetland dieback on carbon dynamics: A comparison between intact and degraded mangroves. Sci. Total Env. 2021, 753, 141817. [Google Scholar] [CrossRef]
- Ahalya, A.; Park, J.S. Blue Carbon Stock of Mangrove Ecosystems. Int. J. Sci. Res. 2018, 8, 1371–1375. Available online: https://www.ijsr.net/getabstract.php?paperid=ART20203497 (accessed on 16 November 2023).
- Kauffman, J.B.; Adame, M.F.; Arifanti, V.D.; Schile-Beers, L.M.; Bernardino, A.F.; Bhomia, R.K.; Donato, D.C.; Feller, I.C.; Ferreira, T.O.; Garcia, M.C.; et al. Total ecosystem carbon stocks of mangroves across broad global environmental and physical gradients. Ecol. Monogr. 2020, 90, 1–18. [Google Scholar] [CrossRef]
- Chalermchatwilai, B.; Poungparn, S.; Patanaponpaiboon, P. Distribution of fine-root necromass in a secondary mangrove forest in Trat province, Eastern Thailand. ScienceAsia 2011, 37, 1–5. [Google Scholar] [CrossRef]
- Kairo, J.G.; Bosire, J.; Langat, J.; Kirui, B.; Koedam, N. Allometry and biomass distribution in replanted mangrove plantations at Gazi Bay, Kenya. Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, S63–S69. [Google Scholar] [CrossRef]
- Omar, R.; Garza-Caligaris, J.F.; Kanninen, M.; Karjalainen, T.; Liski, J.; Nabuurs, G.J.; Pussinen, A.; Jong, B.H.J.; Mohren, G.M.J. Modeling carbon sequestration in afforestation, agroforestry and forest management projects: The CO2 FIX vol 2 approach. Ecol. Model. 2003, 164, 177–199. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Component | Species | Equation | Reference |
|---|---|---|---|
| BA: Live trees | Rm | B = 0.722*ρ*(DR)1.731 | Smith and Whelan [33] |
| Lr | B = 0.362*ρ*(DS)1.92 |
| San Ignacio | El Dátil | |||||||
|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | F | p-Value | |
| pH | 7.5 ± 0.02 | 7.6 ± 0.02 | 7.1 ± 0.09 | 6.6 ± 0.01 | 6.9 ± 0.1 | 6.5 ± 0.11 | 9.8 | 0.24 |
| Temperature (°C) | 20.1 ± 0.08 | 20.1 ± 0.1 | 20.4 ± 0.1 | 22.3 ± 0.07 | 22.6 ± 0.27 | 22.3 ± 0.08 | 11.2 | 0.04 |
| Conductivity (mS/cm) | 57 ± 1.3 | 58 ± 0.1 | 67 ± 4.6 | 59 ± 0.2 | 55 ± 0.2 | 57 ± 0.3 | 5.1 | 0.55 |
| Salinity (PSU) | 38 ± 1 | 38 ± 0.2 | 46 ± 3.6 | 40 ± 0.1 | 36 ± 0.2 | 38 ± 0.2 | 6.4 | 0.65 |
| Redox potential (mV) | −133 ± 2 | −122 ± 3 | −121 ± 3 | −290 ± 1.4 | −263 ± 4.4 | −191 ± 4.2 | 7.4 | 0.62 |
| San Ignacio | El Dátil | |||||||
|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | F | p-Value | |
| pH | 6.5 ± 0.08 | 7 ± 0.08 | 6.9 ± 0.05 | 6.3.4 ± 0.1 | 6.5 ± 0.02 | 7.2 ± 0.14 | 14.1 | 0.43 |
| Temperature (°C) | 21 ± 0.1 | 22 ± 0.6 | 24 ± 1 | 25 ± 1 | 23 ± 0.3 | 21 ± 0.5 | 5.8 | 0.38 |
| Conductivity (mS/cm) | 58 ± 1.9 | 56 ± 1.3 | 54 ± 1.1 | 55 ± 0.1 | 56 ± 0.2 | 67 ± 2.5 | 3.8 | 0.08 |
| Salinity (PSU) | 38 ± 1.4 | 38 ± 1 | 36 ± 0.8 | 37 ± 0.1 | 37 ± 0.1 | 46 ± 2.3 | 4.2 | 0.06 |
| Redox potential (mV) | −134 ± 5.6 | −133 ± 2.2 | −130 ± 1.6 | −416 ± 9.7 | −338 ± 66 | −112 ± 4.1 | 6.5 | 0.49 |
| El Dátil | San Ignacio | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | Control San Ignacio | Control El Dátil | F | p-Value | ||
| pH | 7 ± 0.26 | 6 ± 0.24 | 7 ± 0.07 | 7 ± 0.06 | 7 ± 0.05 | 7 ± 0.11 | 9 ± 0.05 | 9 ± 0.17 | 24 | <0.05 | |
| Humidity | 0.37 ± 0.02 | 0.29 ± 0.01 | 0.29 ± 0.01 | 0.31 ± 0.03 | 0.35 ± 0.01 | 0.62 ± 0.01 | 0.16 ± 0.01 | 0.2 ± 0.02 | 22 | <0.05 | |
| BD | 0.9 ± 0.07 | 1.1 ± 0.06 | 1 ± 0.08 | 1.1 ± 0.07 | 1 ± 0.04 | 0.4 ± 0.02 | 1.1 ± 0.03 | 1.2 ± 0.01 | 16 | 0.06 | |
| OM | 2.7 ± 0.5 | 1.5 ± 0.5 | 2.1 ± 0.3 | 3.1 ± 0.4 | 2.2 ± 0.3 | 14.8 ± 1.34 | 0.08 | 0.05 | 61 | <0.05 | |
| Texture | Sand | 90 ± 1.1 | 88 ± 1.3 | 90 ± 0.5 | 82 ± 0.8 | 85 ± 0.8 | 85 ± 1.1 | 92 ± 0.9 | 93 ± 0.6 | 17 | 0.6 |
| Silt | 4 ± 0.8 | 4 ± 0.9 | 1 ± 0.5 | 6 ± 2.2 | 6 ± 0.5 | 3 ± 0.6 | 2 ± 0.4 | 1 ± 0.7 | 4.2 | 0.06 | |
| Clay | 6 ± 0.5 | 8 ± 1.3 | 8 ± 0.5 | 12 ± 1.5 | 9 ± 1.1 | 12 ± 0.7 | 7 ± 0.7 | 6 ± 0.6 | 7.1 | 0.16 |
| Site | Species | Density | Height | DS (cm) | Baseline Area (ha) | Cover (%/ha) | |
|---|---|---|---|---|---|---|---|
| (Ind/ha) | (m) | ||||||
| El Dátil | Site 1 | Rm | 4200 | 1.1 ± 0.04 | 4.4 ± 0.27 | 4.5 ± 0.7 | 55 ± 10.8 |
| Lr | 1600 | 1.4 ± 0.1 | 8.1 ± 0.7 | 14 ± 2.3 | 43 ± 8.1 | ||
| Site 2 | Rm | 2300 | 0.8 ± 0.05 | 4.1 ± 0.27 | 3.7 ± 0.4 | 15 ± 0.9 | |
| Lr | 1500 | 1.2 ± 0.1 | 4.3 ± 0.5 | 4 ± 0.9 | 10 ± 1 | ||
| Site 3 | Rm | 1700 | 0.6 ± 0.03 | 1.9 ± 0.2 | 0.8 ± 0.2 | 1 ± 0.4 | |
| Lr | 3300 | 1.9 ± 0.1 | 6.9 ± 0.6 | 11 ± 1.4 | 40 ± 5.4 | ||
| San Ignacio | Site 4 | Rm | 2200 | 0.7 ± 0.07 | 5.2 ± 0.37 | 5.9 ± 0.9 | 8 ± 1.5 |
| Lr | 1100 | 2.1 ± 0.1 | 8 ± 1.3 | 16 ± 2.7 | 76 ± 15.7 | ||
| Site 5 | Rm | 2800 | 1.2 ± 0.08 | 5.8 ± 0.5 | 7.8 ± 1.5 | 28 ± 4.8 | |
| Lr | 1200 | 2.3 ± 0.1 | 7.9 ± 0.6 | 13 ± 1.9 | 35 ± 5.4 | ||
| Site 6 | Rm | 1800 | 1.6 ± 0.14 | 7.7 ± 0.33 | 12 ± 1 | 36 ± 5.9 | |
| Lr | 700 | 2 ± 0.02 | 4.9 ± 0.3 | 5 ± 0.6 | 31 ± 1.1 |
| El Datil | San Ignacio | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Root Class | Stratum | Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | F | p-Value |
| Fine | 0–15 | 1297 ± 329 | 1257 ± 276 | 1073 ± 367 | 867 ± 173 | 535 ± 130 | 1988 ± 264 | 1.5 | 0.24 |
| 15–30 | 2169 ± 257 | 1133 ± 302 | 937 ± 174 | 301 ± 57 | 618 ± 92 | 1476 ± 380 | 1.9 | 0.15 | |
| 30–45 | 2708 ± 394 a | 1038 ± 240 b | 613 ± 83 b | 517 ± 301 b | 396 ± 75 b | 934 ± 279 b | 6.1 | 0.01 | |
| Sum | 6174 ± 980 | 3428 ± 818 | 2623 ± 624 | 1685 ± 531 | 1549 ± 297 | 4398 ± 923 | |||
| Medium | 0–15 | 266 ± 60 | 217 ± 105 | 1187 ± 107 | 275 ± 91 | 237 ± 28 | 459 ± 106 | 9.9 | 0.16 |
| 15–30 | 237 ± 44 a | 320 ± 265 ab | 538 ± 128 b | 432 ± 135 ab | 331 ± 36 ab | 293 ± 88 ab | 1.6 | 0.01 | |
| 30–45 | 293 ± 51 | 190 ± 52 | 356 ± 62 | 313 ± 76 | 147 ± 31 | 157 ± 42 | 1.1 | 0.46 | |
| sum | 796 ± 155 | 727 ± 422 | 2081 ± 297 | 1020 ± 302 | 715 ± 95 | 909 ± 236 | |||
| Large | 0–15 | 1902 ± 339 | 1247 ± 362 | 885 ± 191 | 536 ± 116 | 720 ± 136 | 1195 ± 294 | 3.5 | 0.11 |
| 15–30 | 1142 ± 307 | 938 ± 306 | 880 ± 123 | 1622 ± 341 | 365 ± 73 | 1140 ± 256 | 1.3 | 0.12 | |
| 30–45 | 1848 ± 243 | 738 ± 307 | 535 ± 204 | 1341 ± 325 | 300 ± 161 | 657 ± 196 | 1.9 | 0.13 | |
| Sum | 4892 ± 889 | 2923 ± 975 | 2300 ± 518 | 3499 ± 782 | 1385 ± 370 | 2992 ± 746 | |||
| TOTAL | 11,862 ± 2024 | 7078 ± 2215 | 7004 ± 1439 | 6204 ± 1615 | 3649 ± 762 | 8299 ± 1905 |
| Above-Ground | Below-Ground | |||||
|---|---|---|---|---|---|---|
| Site | Structure | Leaf Litter | Roots | Necromass | Total | |
| El Dátil | 1 | 29.8 ± 3.6 | 0.15 ± 0.04 | 46.3 ± 7.9 | 11 ± 3 | 87.3 ± 14.5 |
| 2 | 11.4 ± 1.1 | 0.29 ± 0.09 | 27.6 ± 8.6 | 10.6 ± 2.4 | 49.9 ± 12.12 | |
| 3 | 25.8 ± 3.7 | 0.45 ± 0.07 | 27.3 ± 5.6 | 5.9 ± 3.1 | 59.5 ± 12.5 | |
| San Ignacio | 4 | 21.7 ± 3.7 | 0.47 ± 0.1 | 24.2 ± 6.3 | 5.2 ± 1.6 | 51.6 ± 11.7 |
| 5 | 26.2 ± 3.1 | 0.1 ± 0.02 | 14.2 ± 3 | 4.2 ± 0.6 | 44.7 ± 6.7 | |
| 6 | 18.3 ± 1.6 | 0.56 ± 0.08 | 32.4 ± 7.4 | 11.4 ± 4.1 | 62.7 ± 13.2 | |
| El Dátil | San Ignacio | |||||||
|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Control El Dátil | Site 4 | Site 5 | Site 6 | Control San Ignacio | |
| Corg (%) | 1.2 | 0.76 | 0.09 | 0.07 | 0.28 | 0.35 | 1.7 | 0 |
| Corg (MgC ha−1) | 75.6 | 31.13 | 5.7 | 7.3 | 28.9 | 16.2 | 96.9 | 0 |
| Site | AGC | BGC | Total Corg | Root/Shoot 1 | Root/Shoot 2 | |
|---|---|---|---|---|---|---|
| El Dátil | Site 1 | 29.95 | 132.9 | 162.9 | 1.9 | 4.4 |
| Site 2 | 11.69 | 69.33 | 81.1 | 3.3 | 5.9 | |
| Site 3 | 26.25 | 38.9 | 65.2 | 1.3 | 1.5 | |
| San Ignacio | Site 4 | 22.17 | 58.3 | 80.5 | 1.3 | 2.6 |
| Site 5 | 26.3 | 34.6 | 60.9 | 0.7 | 1.3 | |
| Site 6 | 18.86 | 140.7 | 159.6 | 2.3 | 7.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, J.R.; Frausto-Illescas, T.; Ortega-Trasviña, C.; Barraza-Guardado, R.H.; Sanchez-Mejía, Z.M.; Choix-Ley, F. Blue Carbon in Mangroves of the Arid Zones of San Ignacio and El Dátil Lagoons, El Vizcaino Biosphere Reserve, Baja California, Mexico. Int. J. Plant Biol. 2023, 14, 1078-1091. https://doi.org/10.3390/ijpb14040078
Torres JR, Frausto-Illescas T, Ortega-Trasviña C, Barraza-Guardado RH, Sanchez-Mejía ZM, Choix-Ley F. Blue Carbon in Mangroves of the Arid Zones of San Ignacio and El Dátil Lagoons, El Vizcaino Biosphere Reserve, Baja California, Mexico. International Journal of Plant Biology. 2023; 14(4):1078-1091. https://doi.org/10.3390/ijpb14040078
Chicago/Turabian StyleTorres, Jony R., Tannia Frausto-Illescas, Celeste Ortega-Trasviña, Ramón H. Barraza-Guardado, Zulia M. Sanchez-Mejía, and Francisco Choix-Ley. 2023. "Blue Carbon in Mangroves of the Arid Zones of San Ignacio and El Dátil Lagoons, El Vizcaino Biosphere Reserve, Baja California, Mexico" International Journal of Plant Biology 14, no. 4: 1078-1091. https://doi.org/10.3390/ijpb14040078
APA StyleTorres, J. R., Frausto-Illescas, T., Ortega-Trasviña, C., Barraza-Guardado, R. H., Sanchez-Mejía, Z. M., & Choix-Ley, F. (2023). Blue Carbon in Mangroves of the Arid Zones of San Ignacio and El Dátil Lagoons, El Vizcaino Biosphere Reserve, Baja California, Mexico. International Journal of Plant Biology, 14(4), 1078-1091. https://doi.org/10.3390/ijpb14040078