
Exploring the Relationship between Forest Canopy Height and Canopy Density from Spaceborne LiDAR Observations




Abstract
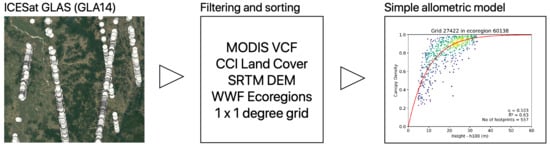
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Datasets
2.1.1. ICESat GLAS
2.1.2. Additional Datasets
2.1.3. World Wildlife Federation Ecoregions
2.2. Pre-Processing
2.3. Methods
3. Results
4. Discussion
4.1. Biomes
4.2. Maximum Canopy Density and Canopy Height Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Means, J.E.; Acker, S.A.; Harding, D.J.; Blair, J.B.; Lefsky, M.A.; Cohen, W.B.; Harmon, M.E.; Mckee, W.A. Use of Large-Footprint Scanning Airborne Lidar To Estimate Forest Stand Characteristics in the Western Cascades of Oregon. Remote Sens. Environ. 1999, 308, 298–308. [Google Scholar] [CrossRef]
- Hansen, A.; Barnett, K.; Jantz, P.; Phillips, L.; Goetz, S.J.; Hansen, M.; Venter, O.; Watson, J.E.; Burns, P.; Atkinson, S.; et al. Global humid tropics forest structural condition and forest structural integrity maps. Sci. Data 2019, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.B.; Clark, D.A. Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. Manag. 2000, 137, 185–198. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933. [Google Scholar] [CrossRef]
- Nelson, R.; Ranson, K.J.; Sun, G.; Kimes, D.S.; Kharuk, V.; Montesano, P. Estimating Siberian timber volume using MODIS and ICESat/GLAS. Remote Sens. Environ. 2009, 113, 691–701. [Google Scholar] [CrossRef]
- Scarth, P.; Armston, J.; Lucas, R.; Bunting, P. A structural classification of Australian vegetation using ICESat/GLAS, ALOS PALSAR, and Landsat sensor data. Remote Sens. 2019, 11, 147. [Google Scholar] [CrossRef] [Green Version]
- Rosette, J.A.; North, P.R.; Suárez, J.C. Vegetation height estimates for a mixed temperate forest using satellite laser altimetry. Int. J. Remote Sens. 2008, 29, 1475–1493. [Google Scholar] [CrossRef]
- Los, S.O.; Rosette, J.A.; Kljun, N.; North, P.R.; Chasmer, L.; Suárez, J.C.; Hopkinson, C.; Hill, R.A.; Van Gorsel, E.; Mahoney, C.; et al. Vegetation height and cover fraction between 60° S and 60° N from ICESat GLAS data. Geosci. Model Dev. 2012, 5, 413–432. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Lefsky, M.; Sun, G.; Miller, M.; Li, Z. Temperate forest height estimation performance using icesat glas data from different observation periods. Int. Arch. Photogramm. Remote. Sens. Spat. Inf.-Sci.-Isprs Arch. 2008, 37, 777–782. [Google Scholar]
- Pang, Y.; Lefsky, M.; Sun, G.; Ranson, J. Impact of footprint diameter and off-nadir pointing on the precision of canopy height estimates from spaceborne lidar. Remote Sens. Environ. 2011, 115, 2798–2809. [Google Scholar] [CrossRef]
- Mahoney, C.; Hopkinson, C.; Held, A.; Kljun, N.; Van Gorsel, E. ICESat/GLAS canopy height sensitivity inferred from airborne Lidar. Photogramm. Eng. Remote Sens. 2016, 82, 351–363. [Google Scholar] [CrossRef]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping forest canopy height globally with spaceborne lidar. J. Geophys. Res. Biogeosciences 2011, 116, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Khalefa, E.; Smit, I.P.; Nickless, A.; Archibald, S.; Comber, A.; Balzter, H. Retrieval of savanna vegetation canopy height from ICESat-GLAS spaceborne LiDAR with terrain correction. IEEE Geosci. Remote. Sens. Lett. 2013, 10, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Armston, J.; Hancock, S.; Marselis, S.; Goetz, S.; Dubayah, R. Characterizing global forest canopy cover distribution using spaceborne lidar. Remote Sens. Environ. 2019, 231, 111262. [Google Scholar] [CrossRef]
- García, M.; Popescu, S.; Riaño, D.; Zhao, K.; Neuenschwander, A.; Agca, M.; Chuvieco, E. Characterization of canopy fuels using ICESat/GLAS data. Remote Sens. Environ. 2012, 123, 81–89. [Google Scholar] [CrossRef]
- Tao, S.; Guo, Q.; Li, C.; Wang, Z.; Fang, J. Global patterns and determinants of forest canopy height. Ecology 2016, 97, 3265–3270. [Google Scholar] [CrossRef]
- Joshi, C.; Leeuw, J.D.; Skidmore, A.K.; Duren, I.C.; van Oosten, H. Remotely sensed estimation of forest canopy density: A comparison of the performance of four methods. Int. J. Appl. Earth Obs. Geoinf. 2006, 8, 84–95. [Google Scholar] [CrossRef]
- Watt, P.J.; Donoghue, D.N. Measuring forest structure with terrestrial laser scanning. Int. J. Remote Sens. 2005, 26, 1437–1446. [Google Scholar] [CrossRef]
- Montesano, P.M.; Nelson, R.; Sun, G.; Margolis, H.; Kerber, A.; Ranson, K.J. MODIS tree cover validation for the circumpolar taiga-tundra transition zone. Remote Sens. Environ. 2009, 113, 2130–2141. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Su, Y.; Guo, Q. Comparison of canopy cover estimations from airborne LiDAR, aerial imagery, and satellite imagery. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 4225–4236. [Google Scholar] [CrossRef]
- Santoro, M.; Cartus, O.; Fransson, J.E. Integration of allometric equations in the water cloud model towards an improved retrieval of forest stem volume with L-band SAR data in Sweden. Remote Sens. Environ. 2021, 253, 112235. [Google Scholar] [CrossRef]
- Hofton, M.A.; Minster, J.B.; Blair, J.B. Decomposition of laser altimeter waveforms. IEEE Trans. Geosci. Remote Sens. 2000, 38, 1989–1996. [Google Scholar] [CrossRef]
- ESA2015. 2015. Available online: www.esa-landcover-cci.org/ (accessed on 25 November 2021).
- DiMiceli, C.; Carroll, M.; Sohlberg, R.; Kim, D.H.; Kelly, M.; Townshend, J.R.G. MOD44B MODIS/Terra Vegetation Continuous Fields Yearly L3 Global 250 m SIN Grid V006. 2015. Available online: https://lpdaac.usgs.gov/products/mod44bv006/ (accessed on 10 September 2020).
- Staver, A.C.; Hansen, M.C. CORRESPON D E N C E Analysis of stable states in global savannas: Is the CART pulling the horse ?—A comment However, the MODIS VCF—which has facilitated major steps in our ability to examine ecological phenomena at global scales—Remains a useful t. Glob. Ecol. Biogeogr. 2015, 24, 985–987. [Google Scholar] [CrossRef]
- Huang, S.; Siegert, F. Land cover classification optimized to detect areas at risk of desertification in North China based on SPOT VEGETATION imagery. J. Arid. Environ. 2006, 67, 308–327. [Google Scholar] [CrossRef]
- Sexton, J.O.; Song, X.P.; Feng, M.; Noojipady, P.; Anand, A.; Huang, C.; Kim, D.H.; Collins, K.M.; Channan, S.; DiMiceli, C.; et al. Global, 30-m resolution continuous fields of tree cover: Landsat-based rescaling of MODIS vegetation continuous fields with lidar-based estimates of error. Int. J. Digit. Earth 2013, 6, 427–448. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Ghilardi, A.; Paneque-Galvez, J.; Skutsch, M.; Mas, J.F. Validation of MODIS Vegetation Continuous Fields for monitoring deforestation and forest degradation: Two cases in Mexico. Geocarto Int. 2015, 31, 1019–1031. [Google Scholar] [CrossRef]
- de Ferranti. 2009. Available online: http://www.viewfinderpanoramas.org (accessed on 15 September 2020).
- Rizzoli, P.; Martone, M.; Gonzalez, C.; Wecklich, C.; Borla Tridon, D.; Bräutigam, B.; Bachmann, M.; Schulze, D.; Fritz, T.; Huber, M.; et al. Generation and performance assessment of the global TanDEM-X digital elevation model. ISPRS J. Photogramm. Remote Sens. 2017, 132, 119–139. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Fashing, P.J.; Forrestel, A.; Scully, C.; Cords, M. Long-term tree population dynamics and their implications for the conservation of the Kakamega Forest, Kenya. Biodivers. Conserv. 2004, 13, 753–771. [Google Scholar] [CrossRef]
- Zhan, X.; DeFries, R.; Hansen, M.; Townshend, J.; DiMiceli, C.; Sohlberg, R.; Huang, C. MODIS Enhanced Land Cover and Land Cover Change Product Algorithm Theoretical Basis Documents (ATBD). 1999. Available online: https://modis.gsfc.nasa.gov/data/atbd/atbd_mod29.pdf (accessed on 12 August 2020).
- NSIDC.org. 2020. Available online: https://nsidc.org/ (accessed on 10 October 2020).
- Scipy.org. 2019. Available online: https://scipy.org/ (accessed on 22 September 2020).
- Sayre, R.; Karagulle, D.; Frye, C.; Boucher, T.; Wolff, N.H.; Breyer, S.; Wright, D.; Martin, M.; Butler, K.; Van Graafeiland, K.; et al. An assessment of the representation of ecosystems in global protected areas using new maps of World Climate Regions and World Ecosystems. Glob. Ecol. Conserv. 2020, 21, e00860. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 134, 850–853. [Google Scholar] [CrossRef] [Green Version]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Silva, C.E.; et al. Mapping global forest canopy height through integration of GEDI and Landsat data. Remote Sens. Environ. 2021, 253, 112165. [Google Scholar] [CrossRef]
- Kier, G.; Mutke, J.; Dinerstein, E.; Ricketts, T.H. Global patterns of plant diversity and floristic knowledge. J. Biogeogr. 2005, 32, 1107–1116. [Google Scholar] [CrossRef]
- Sánchez-Azofeifa, G.A.; Quesada, M.; Rodríguez, J.P.; Nassar, J.M.; Stoner, K.E.; Castillo, A.; Garvin, T.; Zent, E.L.; Calvo-Alvarado, J.C.; Kalacska, M.E.; et al. Research priorities for neotropical dry forests. Biotropica 2005, 37, 477–485. [Google Scholar] [CrossRef]
- Flint, E.P.; Richards, J.F. Trends in Carbon Content of Vegetation in South and Southeast Asia Associated with Changes in Land Use. In Effects of Land-Use Change on Atmospheric CO2 Concentrations; Dale, V.H., Ed.; Springer: New York, NY, USA, 1994; pp. 201–299. [Google Scholar] [CrossRef]
- Worldwildlife. Available online: https://www.worldwildlife.org/publications/terrestrial-ecoregions-of-the-world (accessed on 12 September 2020).
- Murphy, S.J.; Audino, L.D.; Whitacre, J.; Eck, J.L.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Species associations structured by environment and land-use history promote beta-diversity in a temperate forest. Ecology 2015, 96, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runkle, J.R. Gap dynamics in an Ohio Acer–Fagus forest and speculations on the geography of disturbance. Can. J. For. Res. 1990, 20, 632–641. [Google Scholar] [CrossRef]
- Zackrisson, O. Nordic Society Oikos Influence of Forest Fires on the North Swedish Boreal Forest. Oikos 1977, 29, 22–32. [Google Scholar] [CrossRef]
- Fuentes, D.A.; Gamon, J.A.; Qiu, H.L.; Sims, D.A.; Roberts, D.A. Mapping Canadian boreal forest vegetation using pigment and water absorption features derived from the AVIRIS sensor. J. Geophys. Res. Atmos. 2001, 106, 33565–33577. [Google Scholar] [CrossRef] [Green Version]
- Viers, J.; Prokushkin, A.S.; Pokrovsky, O.S.; Auda, Y.; Kirdyanov, A.V.; Beaulieu, E.; Zouiten, C.; Oliva, P.; Dupré, B. Seasonal and spatial variability of elemental concentrations in boreal forest larch foliage of Central Siberia on continuous permafrost. Biogeochemistry 2013, 113, 435–449. [Google Scholar] [CrossRef]
- Hirota, M.; Holmgren, M.; van Nes, E.H.; Scheffer, M. Global Resilience of Tropical Forest. Science 2011, 334, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Sturtevant, B.R.; Hanberry, B.B. Processes underlying restoration of temperate savanna and woodland ecosystems: Emerging themes and challenges. For. Ecol. Manag. 2021, 481, 2019–2022. [Google Scholar] [CrossRef]
- Wellington, A.B. Leaf water potentials, fire and the regeneration of mallee eucalypts in semi-arid, south-eastern Australia. Oecologia 1984, 64, 360–362. [Google Scholar] [CrossRef] [PubMed]
- Hopper, S. Biodiversity in managed landscapes: Theory and practice. In The Use of Genetic Information in Establishing Reserves for Nature Conservation; Szaro, R., Johnston, D.W., Eds.; Oxford University Press: Oxford, UK, 1996; pp. 253–260. [Google Scholar]
- White, F. The Vegetation of Africa; Natural Resources Research 20; UNESCO: Paris, France, 1983; ISBN 92-3-101955-4. [Google Scholar]
- Ecological Stratification Working Group. A National Framework for Canada; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 1996; Available online: https://sis.agr.gc.ca/cansis/publications/manuals/1996/A42-65-1996-national-ecological-framework.pdf (accessed on 12 August 2020).
- Lee, S.; Ni-Meister, W.; Yang, W.; Chen, Q. Physically based vertical vegetation structure retrieval from ICESat data: Validation using LVIS in White Mountain National Forest, New Hampshire, USA. Remote Sens. Environ. 2011, 115, 2776–2785. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kay, H.; Santoro, M.; Cartus, O.; Bunting, P.; Lucas, R. Exploring the Relationship between Forest Canopy Height and Canopy Density from Spaceborne LiDAR Observations. Remote Sens. 2021, 13, 4961. https://doi.org/10.3390/rs13244961
Kay H, Santoro M, Cartus O, Bunting P, Lucas R. Exploring the Relationship between Forest Canopy Height and Canopy Density from Spaceborne LiDAR Observations. Remote Sensing. 2021; 13(24):4961. https://doi.org/10.3390/rs13244961
Chicago/Turabian StyleKay, Heather, Maurizio Santoro, Oliver Cartus, Pete Bunting, and Richard Lucas. 2021. "Exploring the Relationship between Forest Canopy Height and Canopy Density from Spaceborne LiDAR Observations" Remote Sensing 13, no. 24: 4961. https://doi.org/10.3390/rs13244961
APA StyleKay, H., Santoro, M., Cartus, O., Bunting, P., & Lucas, R. (2021). Exploring the Relationship between Forest Canopy Height and Canopy Density from Spaceborne LiDAR Observations. Remote Sensing, 13(24), 4961. https://doi.org/10.3390/rs13244961