Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs


Abstract
:1. Introduction
- Do uncoupled (LSMs) models correctly reproduce the spatial variability of LAI shown by satellite data over the Northern Hemisphere?
- How does the length of the growing season in the different models compare with the satellite data? And where are the main discrepancies (onset or dormancy)?
- What are the trends in LAI and the growing season over this period?
2. Materials and Methods
2.1. Model Data
- S1: real CO2 growth and climate kept constant, recycling the first 10 years of the century.
- S2: real CO2 and climate. In the present study we use the S2 simulations. All model outputs were regridded to a common 1 × 1 degree grid. Although satellite data are available before 1986, we focus on the last 20 years of the 20th century simulations (1986–2005) to be consistent with the analyses of the coupled models (Anav et al., this issue [22]).
2.2. LAI Parameterization and Calculation
- CLM4CN. The model has 16 PFTs. In this version the carbon-nitrogen cycling model simulates leaf carbon and specific leaf area to calculate the LAI for each PFT.
- LPJ. The leaf area index is updated daily and depends on temperature, soil water, and plant productivity for each PFT. The models have 3 different phenology types (evergreen, summergreen, raingreen) and 11 PFTs.
- LPJ-GUESS. The leaf area index is updated daily and depends on temperature, soil water, and plant productivity for each PFT. The models have 3 different phenology types (evergreen, summergreen, raingreen) and 11 PFTs.
- ORCHIDEE. LAI is estimated based on temperature. It also uses a maximum LAI threshold after which no more carbon is allocated to the leaves.
- OCN employs an approach based on the pipe-model for allocation, which results in much more rapid leaf development, and does not prescribe a maximum leaf area-rather, the maximal annual LAI is an emergent outcome of the NPP of the vegetation and the costs (roots, shoot) for maintaining the leaf area, which varies as a function of water and nitrogen stress.
- SDGVM. LAI is calculated to optimize stem & root NPP. This is achieved through consideration of the net carbon balance of the bottom layer of the canopy. The fraction of NPP available for leaf production is adjusted each year based on this carbon balance. The rate at which this fraction is adjusted is PFT-dependent.
- TRIFFID. LAI is calculated for each of the 5 PFTs, based on parameters describing the minimum, maximum and balanced LAI if full cover is reached. The actual LAI is then calculated as a function of the balanced LAI and the phonological status of the vegetation, which depends on temperature.
- VEGAS. The model has five PFTs: broadleaf tree, needleleaf tree, C3 grass, C4 grass, and crop. Whether a tree PFT is deciduous or evergreen is dynamically determined, so it has essentially 7 functional types. Phenology is calculated for each PFT as the balance between growth and respiration. The actual leaf mass is calculated based on photosynthesis allocation, and then converted to leaf area index.
2.3. Satellite Data
2.4. Study Region
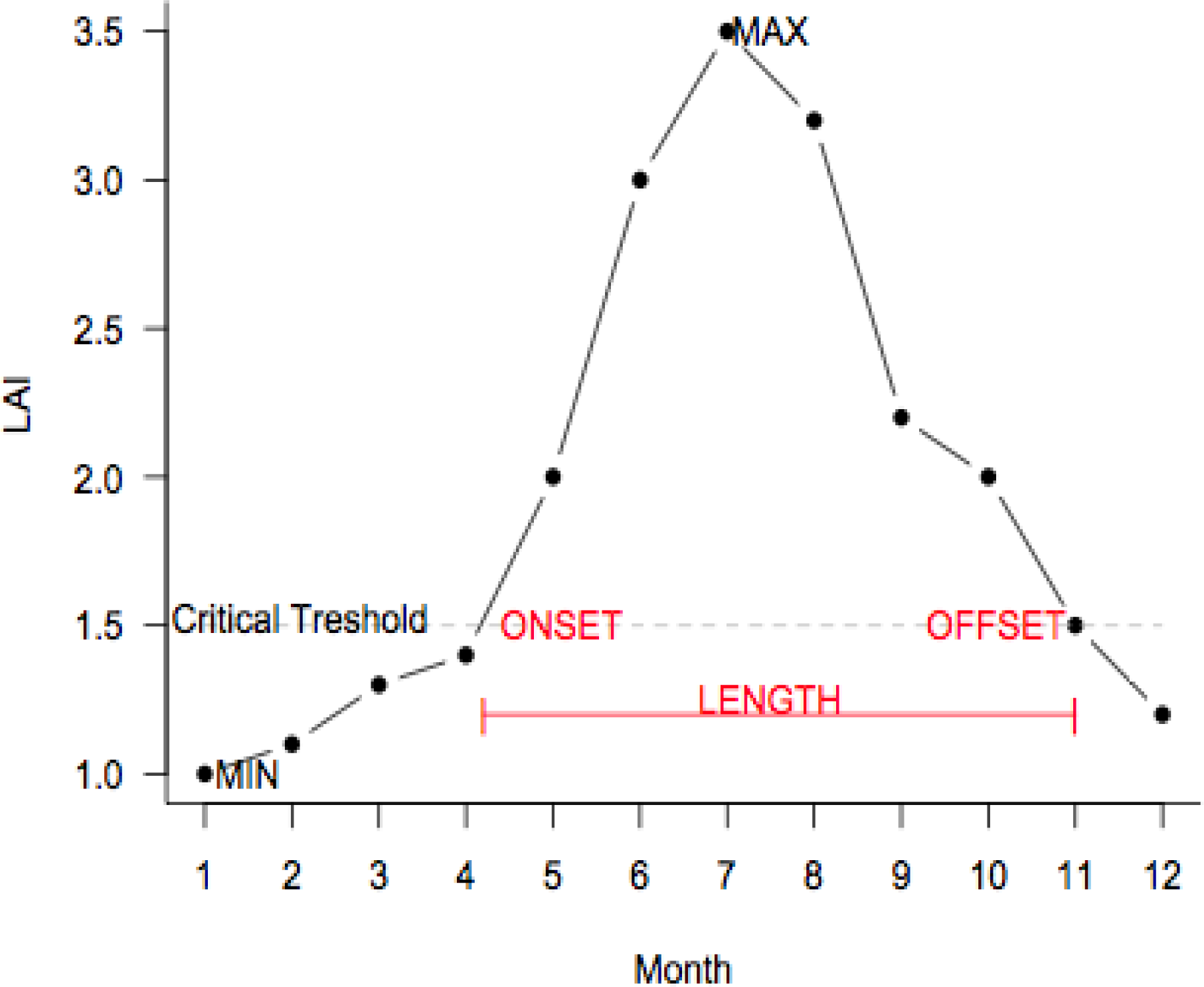
2.5. Leaf Phenology Analyses
2.6. Temporal Trends
3. Results
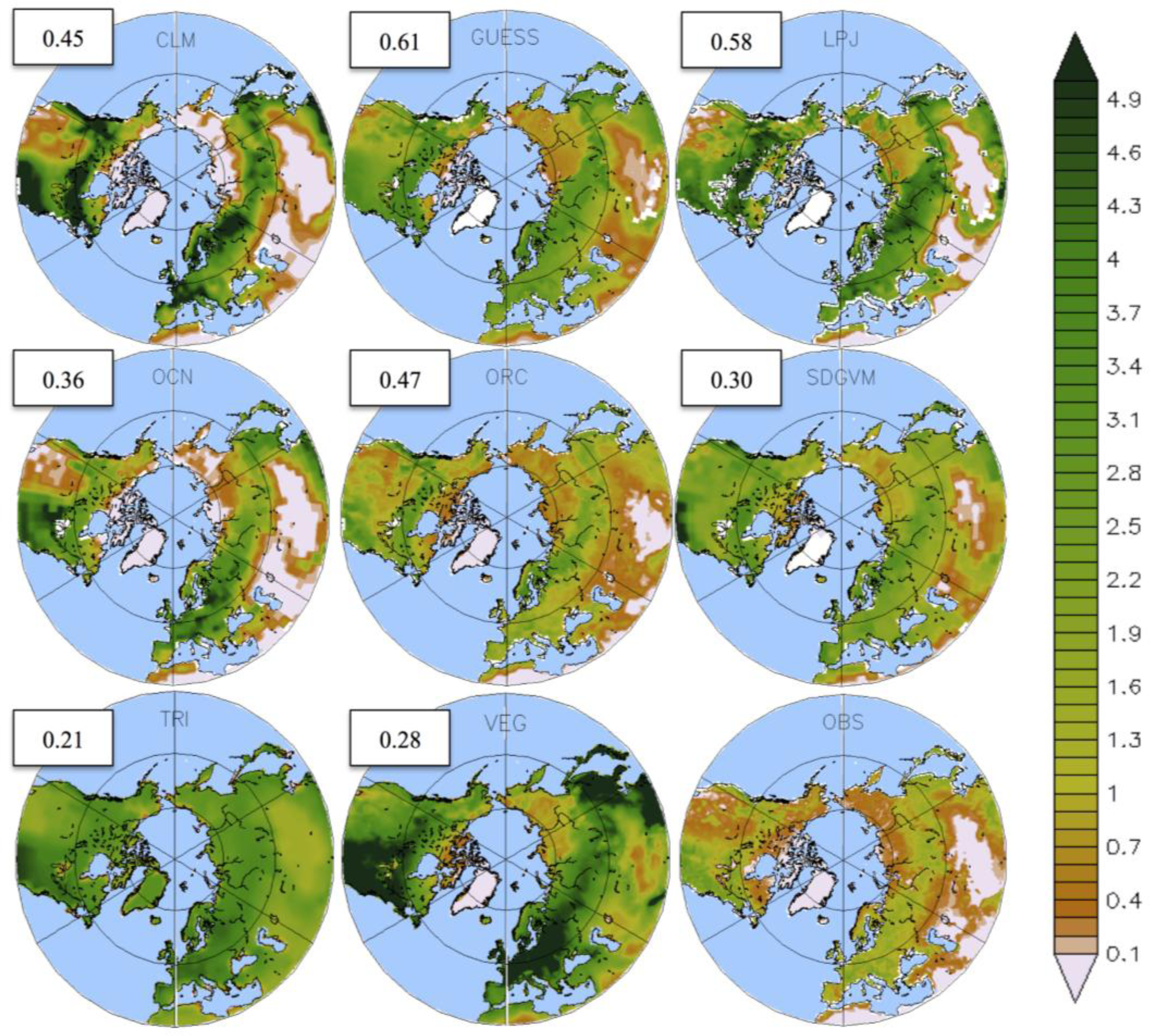
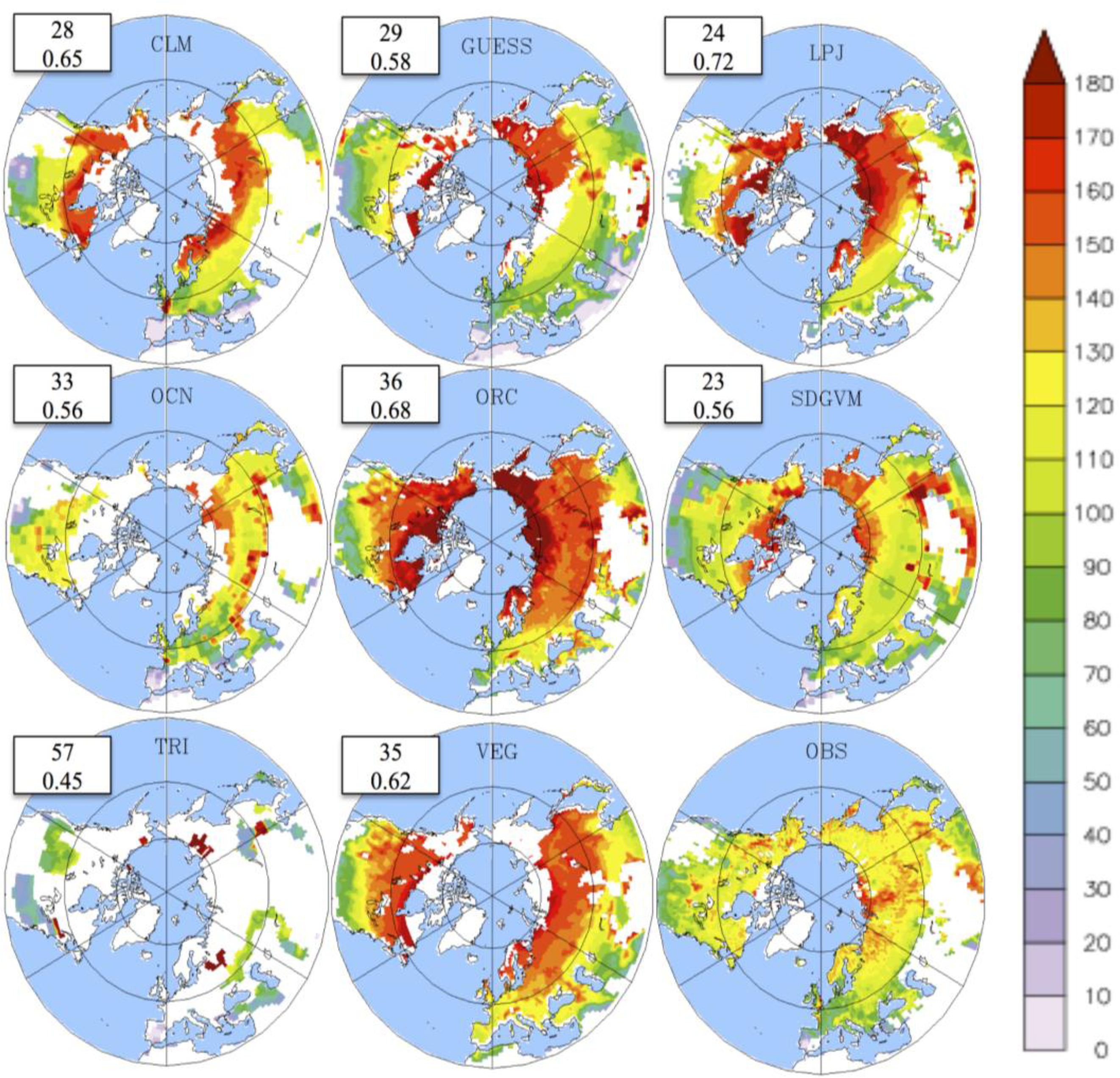
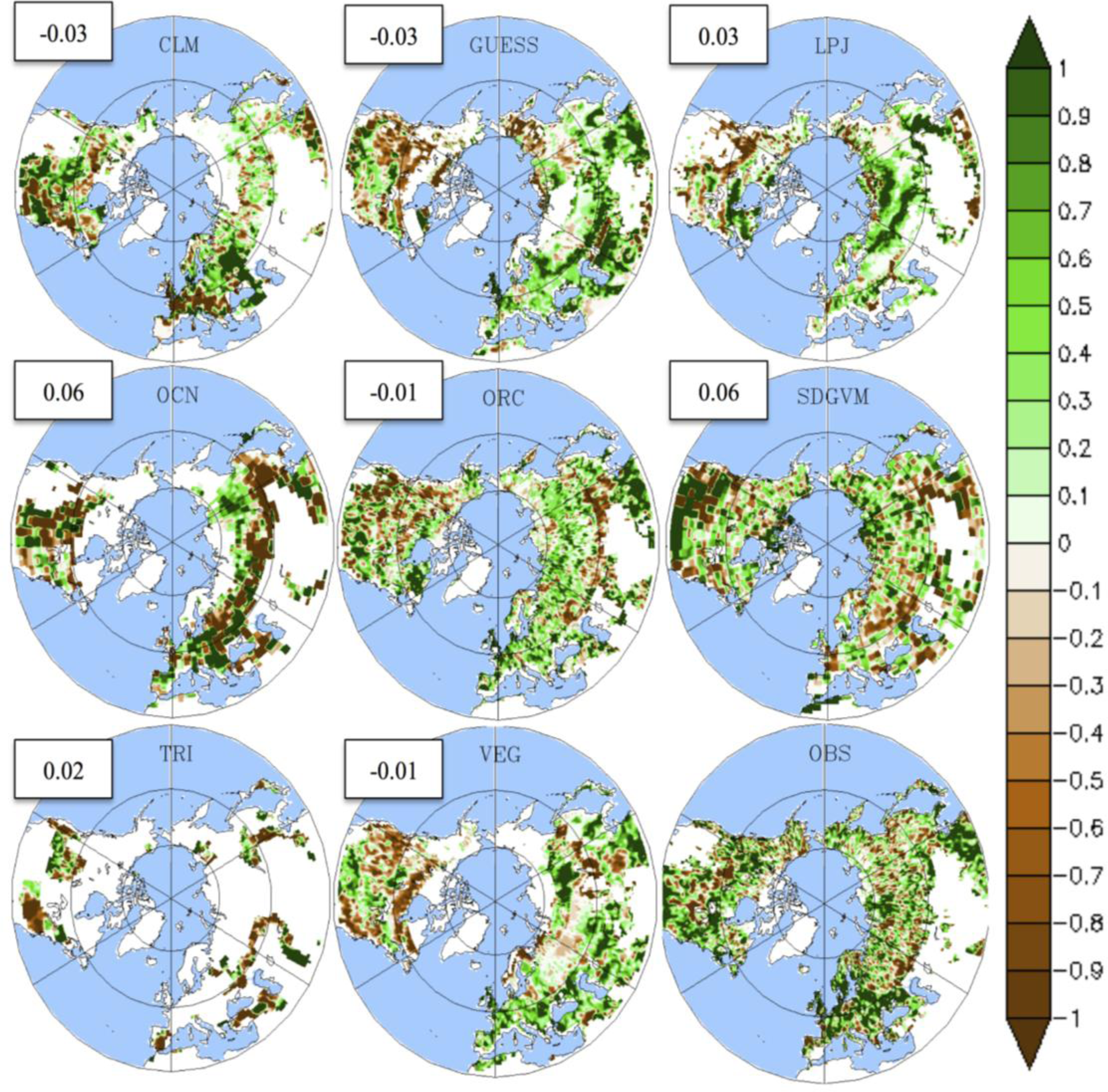
3.1. Mean LAI
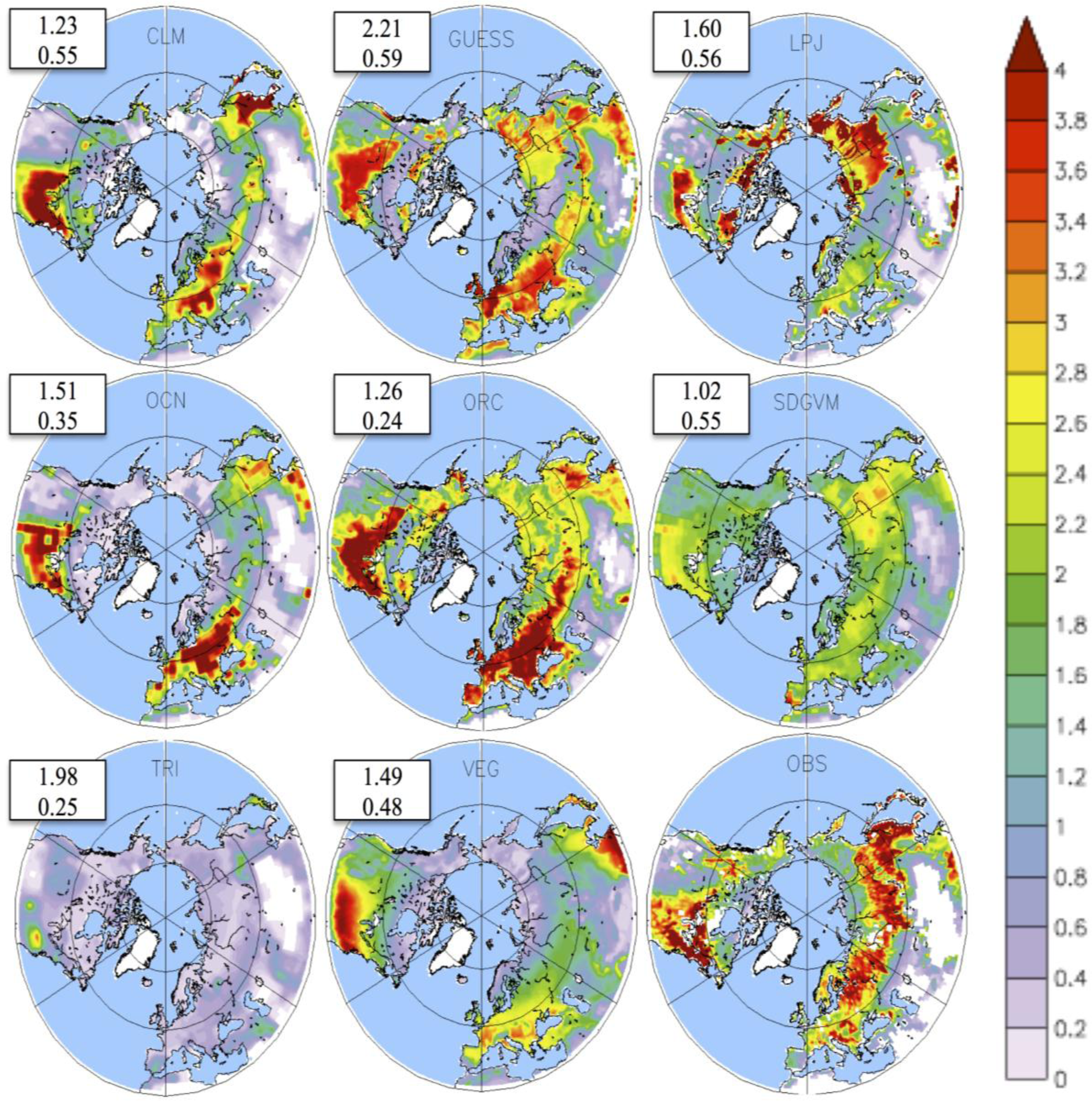
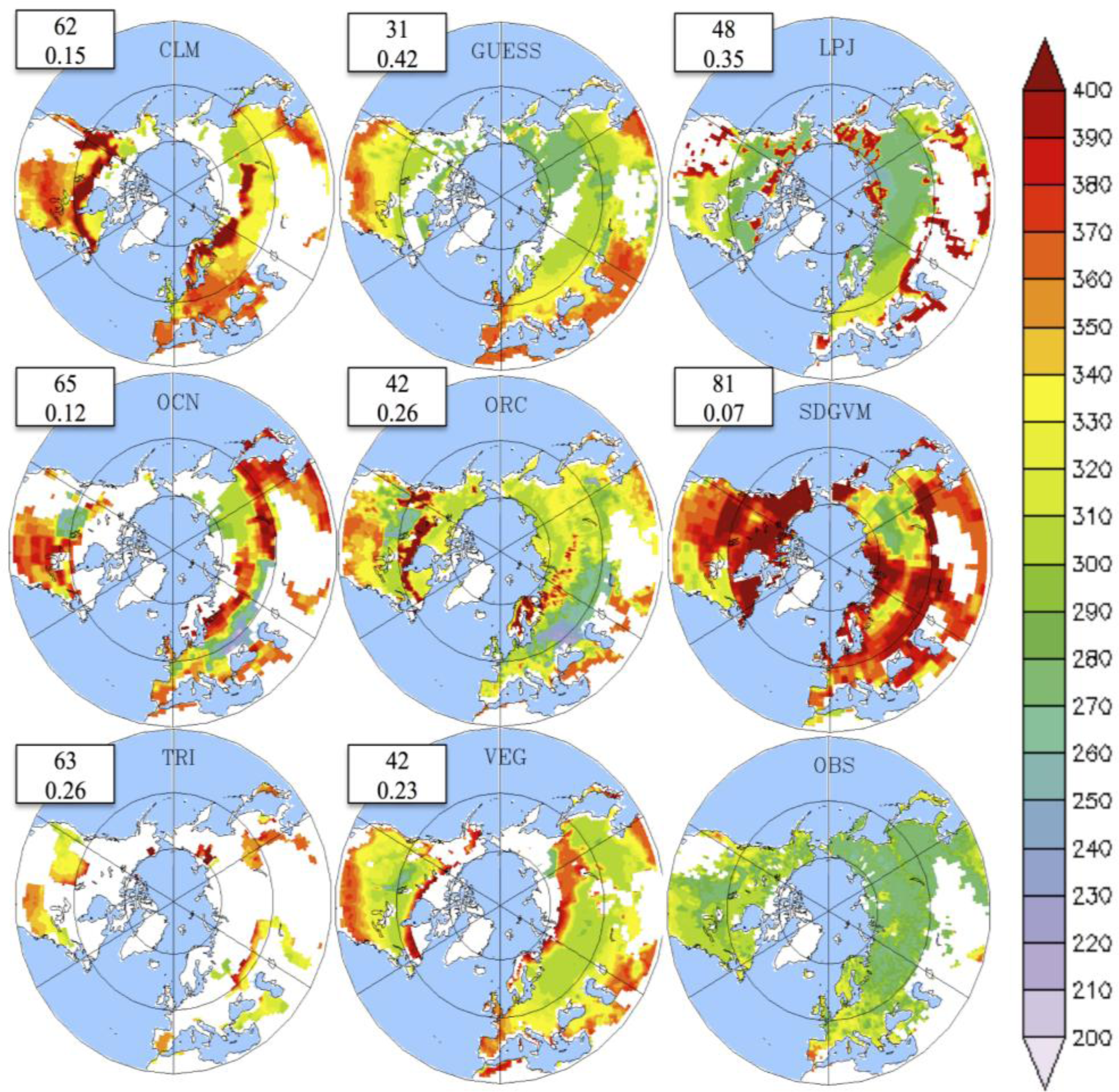
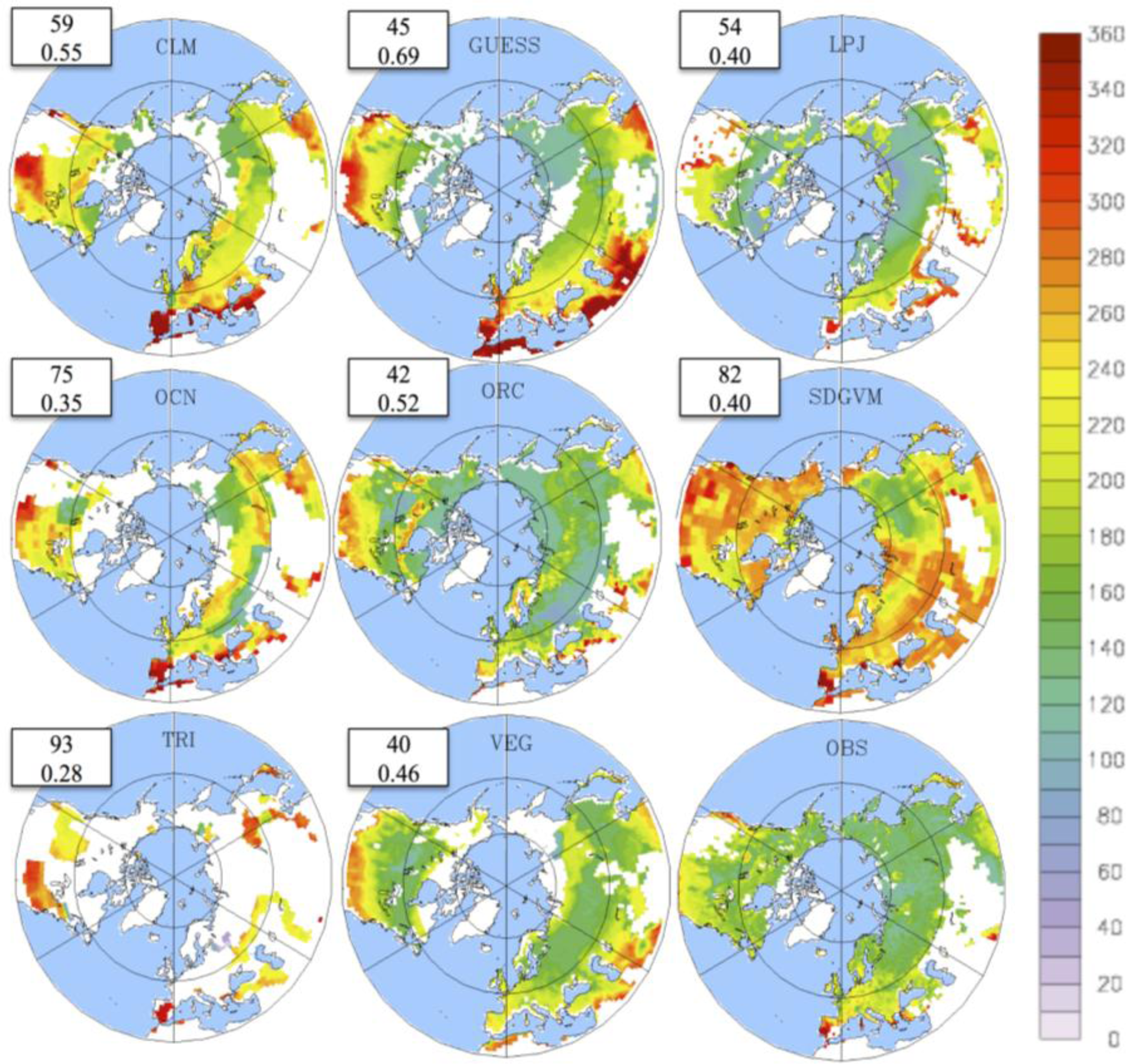
3.2. Growing Season
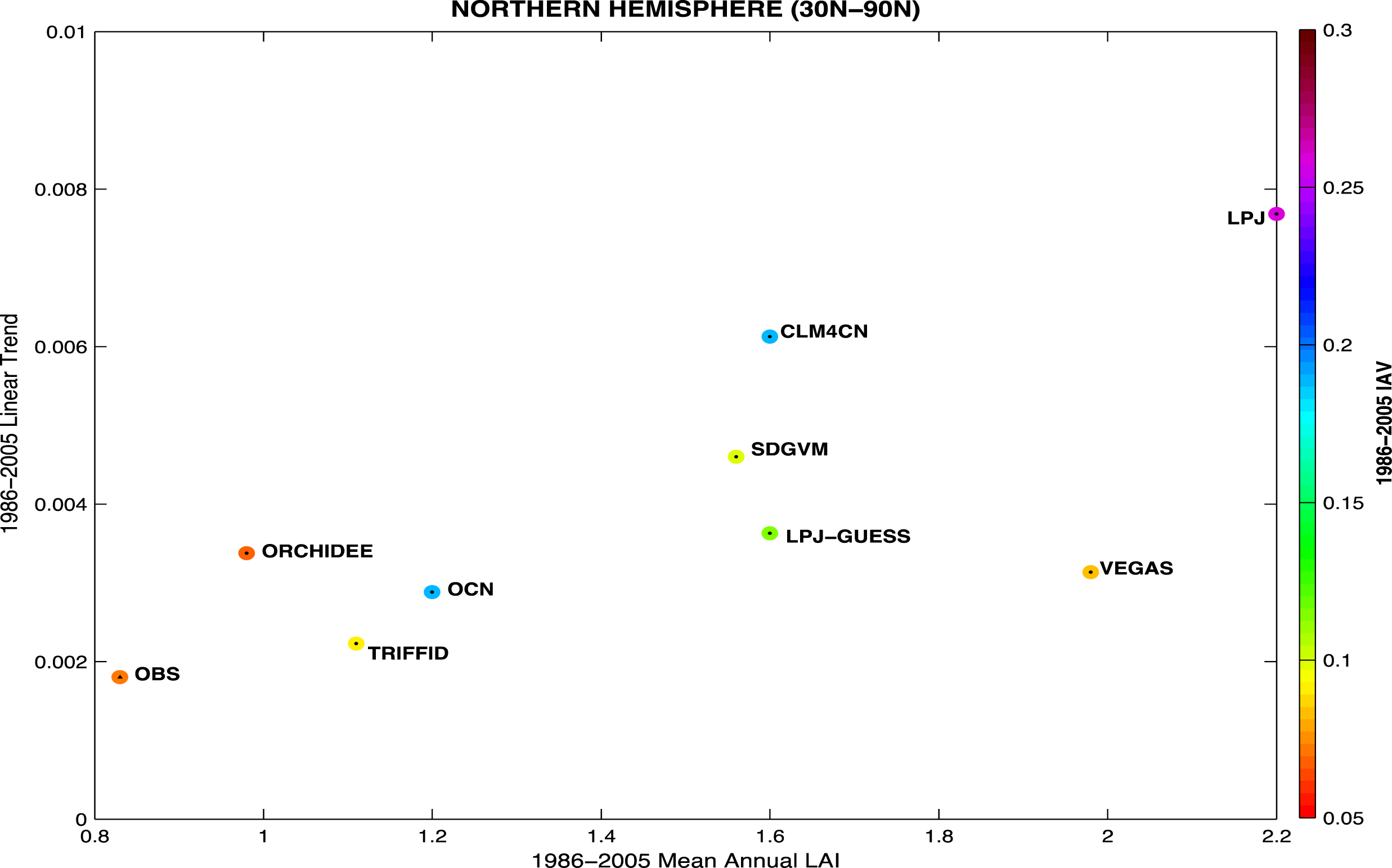
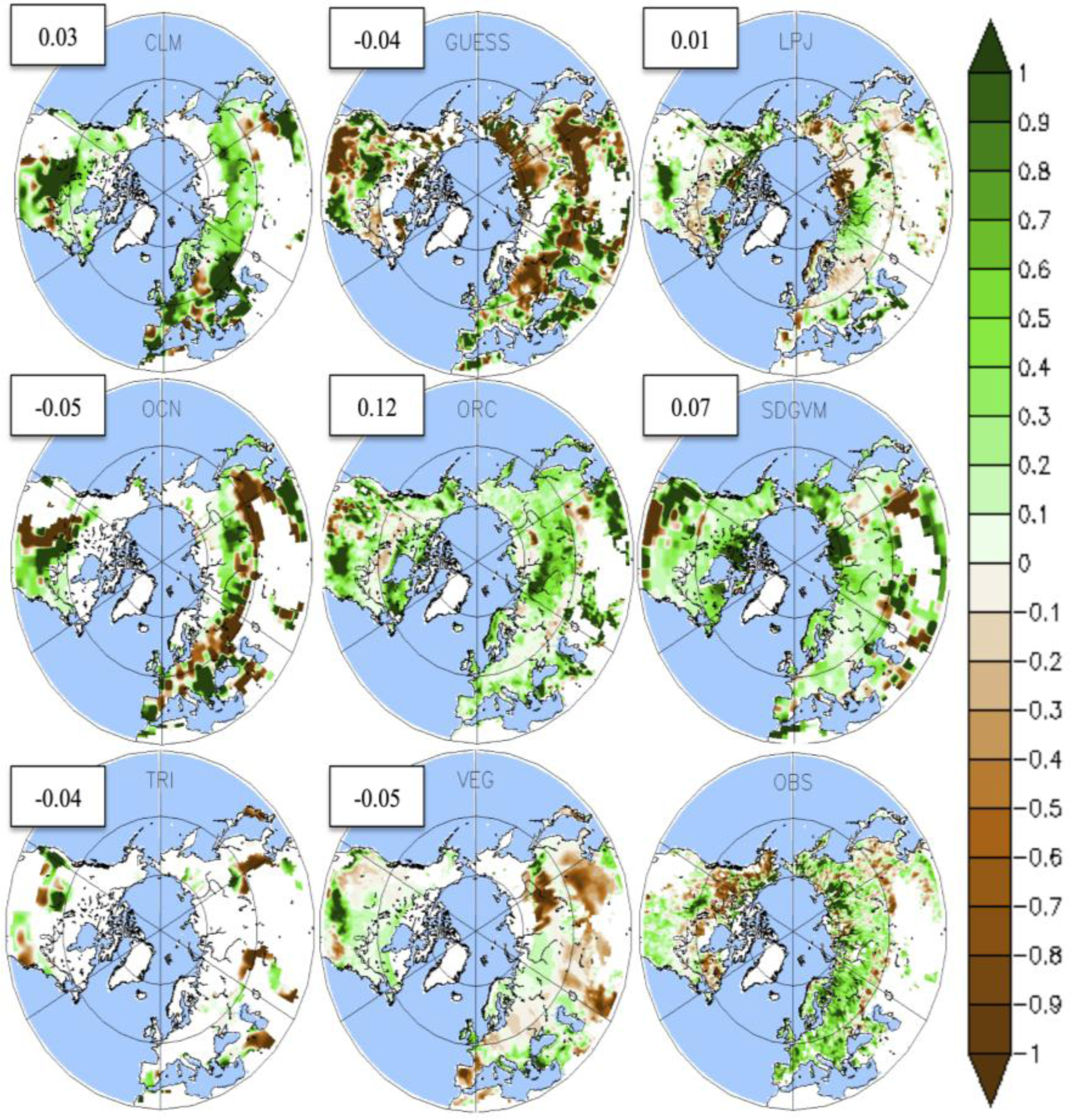
3.3. Temporal Trends
4. Discussion
5. Conclusion
Acknowledgments
Conflict of Interest
References
- Gomez-Peralta, D.; Oberbauer, S.F.; McClain, M.E.; Philippi, T.E. Rainfall and cloud-water interception in tropical montane forests in the eastern Andes of Central Peru. For. Ecol. Manag 2008, 255, 1315–1325. [Google Scholar]
- Running, S.W.; Coughlan, J.C. A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecol. Model 1988, 42, 125–154. [Google Scholar]
- Bréda, N.J.J. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. J. Exp. Bot 2003, 54, 2403–2417. [Google Scholar]
- Knyazikhin, Y.; Martonchik, J.V.; Diner, D.J.; Myneni, R.B.; Verstraete, M.; Pinty, B.; Gobron, N. Estimation of vegetation canopy leaf area index and fraction of absorbed photosynthetically active radiation from atmosphere-corrected MISR data. J. Geophys. Res 1998, 103, 32239–32256. [Google Scholar]
- Landsberg, J.J.; Waring, R.H. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For. Ecol. Manag 1997, 95, 209–228. [Google Scholar]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Change Biol 2003, 9, 161–185. [Google Scholar]
- Sitch, S.; Huntingford, C.; Gedney, N.; Levy, P.E.; Lomas, M.; Piao, S.L.; Betts, R.; Ciais, P.; Cox, P.; Friedlingstein, P.; et al. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Glob. Change Biol 2008, 14, 2015–2039. [Google Scholar]
- Sitch, S.; Friedlingstein, P.; Gruber, N.; Jones, S.; Murray-Tortarolo, G.; Ahlstrom, A.; Doney, S.C.; Graven, H.; Heinze, C.; Huntingford, C.; et al. Trends and drivers of the regional-scale sources and sinks of carbon dioxide over the past two decades. Biogeosci. Discuss. 2013, in press.. [Google Scholar]
- Diaz, S.; Cabido, M. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci 1997, 8, 463–474. [Google Scholar]
- Diaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci 1998, 9, 113–122. [Google Scholar]
- Badeck, F.-W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol 2004, 162, 295–309. [Google Scholar]
- Buermann, W.; Dong, J.; Zeng, X.; Myneni, R.B.; Dickinson, R.E. Evaluation of the utility of satellite-based vegetation leaf area index data for climate simulations. J. Clim 2001, 14, 3536–3550. [Google Scholar]
- Maignan, F.; Bréon, F.-M.; Chevallier, F.; Viovy, N.; Ciais, P.; Garrec, C.; Trules, J.; Mancip, M. Evaluation of a global vegetation model using time series of satellite vegetation indices. Geosci. Model Dev 2011, 4, 1103–1114. [Google Scholar]
- Richardson, A.D.; Anderson, R.S.; Arain, M.A.; Barr, A.G.; Bohrer, G.; Chen, G.; Chen, J.M.; Ciais, P.; Davis, K.J.; Desai, A.R.; et al. Terrestrial biosphere models need better representation of vegetation phenology: Results from the North American carbon program site synthesis. Glob. Change Biol 2011, 18, 566–584. [Google Scholar]
- Randerson, J.T.; Hoffman, F.M.; Thornton, P.E.; Mahowald, N.M.; Lindsay, K.; Lee, Y.-H.; Nevison, C.D.; Doney, S.C.; Bonan, G.; Stöckli, R.; et al. Systematic assessment of terrestrial biogeochemistry in coupled climate–carbon models. Glob. Chang. Biol 2009, 15, 2462–2484. [Google Scholar]
- Prince, S.D. A model of regional primary production for use with coarse resolution satellite data. Int. J. Remote Sens 1991, 12, 1313–1330. [Google Scholar]
- Myneni, R.B.; Ramakrishna, R.; Nemani, R.; Running, S.W. Estimation of global leaf area index and absorbed par using radiative transfer models. IEEE Trans. Geosci. Remote Sens 1997, 35, 1380–1393. [Google Scholar]
- Friedl, M.A.; Schimel, D.S.; Michaelsen, J.; Davis, F.W.; Walker, H. Estimating grassland biomass and leaf area index using ground and satellite data. Int. J. Remote Sens 1994, 15, 1401–1420. [Google Scholar]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using Landsat TM images. Remote Sens. Environ 1996, 55, 153–162. [Google Scholar]
- Green, E.P.; Mumby, P.J.; Edwards, A.J.; Clark, C.D.; Ellis, A.C. Estimating leaf area index of mangroves from satellite data. Aquat. Bot 1997, 58, 11–19. [Google Scholar]
- Prentice, I.C.; Bondeau, A.; Cramer, W.; Harrison, S.P.; Hickler, T.; Lucht, W.; Sitch, S.; Smith, B.; Sykes, M.T. Dynamic Global Vegetation Modeling: Quantifying Terrestrial Ecosystem Responses to Large-Scale Environmental Change. In Terrestrial Ecosystems in a Changing World; Canadell, J.G., Pataki, D.E., Pitelka, L.F., Eds.; Springer: New York City, NY, USA, 2007; pp. 175–192. [Google Scholar]
- Anav, A.; Murray-Tortarolo, G.; Friedlingstein, P.; Sitch, S.; Piao, S.; Zhu, Z. Evaluation of land surface models in reproducing satellite Derived leaf area index over the high-latitude northern hemisphere. Part II: Earth system models. Remote Sens 2013, 5, 3637–3661. [Google Scholar]
- Xu, L.; Myneni, R.B.; Chapin, F.S., III; Callaghan, T.V.; Pinzon, J.E.; Tucker, C.J.; Zhu, Z.; Bi, J.; Ciais, P.; Tømmervik, H.; et al. Temperature and vegetation seasonality diminishment over northern lands. Nat. Clim. Change 2013. [Google Scholar] [CrossRef]
- Anav, A.; Menut, L.; Khvorostyanov, D.; Viovy, N. Impact of tropospheric ozone on the Euro-Mediterranean vegetation. Glob. Chang. Biol 2011, 17, 2342–2359. [Google Scholar]
- Yang, W.; Tan, B.; Huang, D.; Rautiainen, M.; Shabanov, N.V.; Wang, Y.; Privette, J.L.; Huemmrich, K.F.; Fensholt, R.; Sandholt, I.; et al. MODIS leaf area index products: From validation to algorithm improvement. IEEE Trans. Geosci. Remote Sens 2006, 44, 1885–1898. [Google Scholar]
- Gao, F.; Morisette, J.T.; Wolfe, R.E.; Ederer, G.; Pedelty, J.; Masuoka, E.; Myneni, R.; Tan, B.; Nightingale, J. An algorithm to produce temporally and spatially continuous MODIS-LAI time series. IEEE Geosci. Remote Sens. Lett 2008, 5, 60–64. [Google Scholar]
- Ganguly, S.; Schull, M.A.; Samanta, A.; Shabanov, N.V.; Milesi, C.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Generating vegetation leaf area index earth system data record from multiple sensors. Part 1: Theory. Remote Sens. Environ 2008, 112, 4333–4343. [Google Scholar]
- Ganguly, S.; Nemani, R.R.; Zhang, G.; Hashimoto, H.; Milesi, C.; Michaelis, A.; Wang, W.; Votava, P.; Samanta, A.; Melton, F.; et al. Generating global Leaf Area Index from Landsat: Algorithm formulation and demonstration. Remote Sens. Environ 2012, 122, 185–202. [Google Scholar]
- Oleson, K.W.; Lawrence, G.B.; Flanner, M.G.; Kluzek, E.; Levis, P.J.S.; Swenson, S.C.; Thornton, E.; Feddema, J.; Heald, C.L.; Lamarque, J.; et al. Technical Description of Version 4.0 of the Community Land Model (CLM); NCAR: Boulder, CO, USA, 2010. [Google Scholar]
- Smith, B.; Prentice, I.C.; Sykes, M.T. Representation of vegetation dynamics in the modelling of terrestrial ecosystems: Comparing two contrasting approaches within European climate space. Glob. Ecol. Biogeogr 2001, 10, 621–637. [Google Scholar]
- Zaehle, S.; Friend, A.D. Carbon and nitrogen cycle dynamics in the O-CN land surface model: 1. Model description, site-scale evaluation, and sensitivity to parameter estimates. Glob. Biogeochem. Cy 2010, 24. [Google Scholar] [CrossRef]
- Krinner, G.; Viovy, N.; de Noblet-Ducoudré, N.; Ogée, J.; Polcher, J.; Friedlingstein, P.; Ciais, P.; Sitch, S.; Prentice, I.C. A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Glob. Biogeochem. Cy 2005, 19. [Google Scholar] [CrossRef]
- Woodward, F.I. Vegetation-climate feedbacks in a greenhouse world. Philos. Trans. R. Soc. Lond. B Biol. Sci 1998, 353, 29–39. [Google Scholar]
- Cox, P. Description of the TRIFFID Dynamic Global Vegetation Model. Techinal Note 24; Met Office: Berks, UK, 2001. Available online: http://climate.uvic.ca/model/common/HCTN_24.pdf (accessed on 23 April 2013).
- Zeng, N.; Qian, H.; Roedenbeck, C.; Heimann, M. Impact of 1998–2002 midlatitude drought and warming on terrestrial ecosystem and the global carbon cycle. Geophys. Res. Lett 2005, 32. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R. B. Global data sets of vegetation Leaf Area Index (LAI)3g and fraction of Photosynthetically Active Radiation (FPAR)3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens 2013, 5, 927–948. [Google Scholar]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cy 2007, 21. [Google Scholar] [CrossRef]
- Zhang, P.; Anderson, B.; Barlow, M.; Tan, B.; Myneni, R.B. Climate-related vegetation characteristics derived from Moderate Resolution Imaging Spectroradiometer (MODIS) leaf area index and normalized difference vegetation index. J. Geophys. Resea.-Atmos 2004, 109. [Google Scholar] [CrossRef]
- Asner, G.P.; Scurlock, J.M.O.; Hicke, J.A. Global synthesis of leaf area index observations: Implications for ecological and remote sensing studies. Glob. Ecol. Biogeogr 2003, 12, 191–205. [Google Scholar]
- Berterretche, M.; Hudak, A.T.; Cohen, W.B.; Maiersperger, T.K.; Gower, S.T.; Dungan, J. Comparison of regression and geostatistical methods for mapping Leaf Area Index (LAI) with Landsat ETM+ data over a boreal forest. Remote Sens. Environ 2005, 96, 49–61. [Google Scholar]
- Wang, Y.; Woodcock, C.E.; Buermann, W.; Stenberg, P.; Voipio, P.; Smolander, H.; Häme, T.; Tian, Y.; Hu, J.; Knyazikhin, Y.; et al. Evaluation of the MODIS LAI algorithm at a coniferous forest site in Finland. Remote Sens. Environ 2004, 91, 114–127. [Google Scholar]
- Bonan, G.B. Importance of Leaf Area Index and forest type when estimating photosynthesis in boreal forests. Remote Sens. Environ 1993, 43, 303–314. [Google Scholar]
- Jeong, S.-J.; Ho, C.-H.; Gim, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol 2011, 17, 2385–2399. [Google Scholar]
- Hickler, T.; Smith, B.; Prentice, I.C.; Mjöfors, K.; Miller, P.; Arneth, A.; Sykes, M.T. CO2 fertilization in temperate FACE experiments not representative of boreal and tropical forests. Glob. Change Biol 2008, 14, 1531–1542. [Google Scholar]
- Norby, R.J.; Sholtis, J.D.; Gunderson, C.A.; Jawdy, S.S. Leaf dynamics of a deciduous forest canopy: No response to elevated CO2. Oecologia 2003, 136, 574–584. [Google Scholar]
- Iivonen, S.; Kaakinen, S.; Jolkkonen, A.; Vapaavuori, E.; Linder, S. Influence of long-term nutrient optimization on biomass, carbon, and nitrogen acquisition and allocation in Norway spruce. Can. J. For. Res 2006, 36, 1563–1571. [Google Scholar]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic control of the high-latitude vegetation greening trend and pinatubo effect. Science 2002, 296, 1687–1689. [Google Scholar]
- De Jong, R.; Verbesselt, J.; Zeileis, A.; Schaepman, M. Shifts in global vegetation activity trends. Remote Sens 2013, 5, 1117–1133. [Google Scholar]
- Mao, J.; Shi, X.; Thornton, P.; Hoffman, F.; Zhu, Z.; Myneni, R. Global latitudinal-asymmetric vegetation growth trends and their driving mechanisms: 1982–2009. Remote Sens 2013, 5, 1484–1497. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Name | Abbreviation | Spatial Resolution | Number of PFTs | Vegetation | Fire dynamics | Full Nitrogen Cycle | References |
|---|---|---|---|---|---|---|---|
| Community Land Model 4CN | CLM | 0.5° × 0.5° | 16 | Imposed | Yes | Yes | [29] |
| Lund-Potsdam-Jena | LPJ | 0.5° × 0.5° | 11 | Dynamic | Yes | No | [6] |
| LPJ-GUESS | GUESS | 0.5° × 0.5° | 11 | Dynamic | Yes | No | [30] |
| ORCHIDEE-CN | OCN | 3.75° × 2.5° | 12 | Imposed | Yes | Yes | [31] |
| ORCHIDEE | ORC | 0.5° × 0.5° | 12 | Imposed | No | No | [32] |
| Sheffield-DGVM | SDGVM | 3.75° × 2.5° | 6 | Imposed | Yes | No | [33] |
| TRIFFID | TRI | 3.75° × 2.5° | 5 | Dynamic | No | No | [34] |
| VEGAS | VEG | 0.5° × 0.5° | 4 | Dynamic | No | No | [35] |
| Model | LAI | Onset (day) | Dormancy (day) | Length (days) |
|---|---|---|---|---|
| CLM | 1.6 | 131 | 351 (288) | 220 (164) |
| LPJ_GUESS | 1.6 | 125 | 314 (285) | 189 (151) |
| LPJ | 2.2 | 130 | 319 (278) | 189 (134) |
| OCN | 1.2 | 121 | 342 (268) | 221 (142) |
| ORCHIDEE | 0.98 | 151 | 323 (268) | 172 (134) |
| SDGVM | 1.56 | 122 | 374 (275) | 252(145) |
| TRIFFID | 1.11 | 133 | 355 (274) | 222(125) |
| VEGAS | 1.98 | 136 | 336 (277) | 200 (139) |
| LAI3g | 0.83 | 111 | 295 | 184 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Murray-Tortarolo, G.; Anav, A.; Friedlingstein, P.; Sitch, S.; Piao, S.; Zhu, Z.; Poulter, B.; Zaehle, S.; Ahlström, A.; Lomas, M.; et al. Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs. Remote Sens. 2013, 5, 4819-4838. https://doi.org/10.3390/rs5104819
Murray-Tortarolo G, Anav A, Friedlingstein P, Sitch S, Piao S, Zhu Z, Poulter B, Zaehle S, Ahlström A, Lomas M, et al. Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs. Remote Sensing. 2013; 5(10):4819-4838. https://doi.org/10.3390/rs5104819
Chicago/Turabian StyleMurray-Tortarolo, Guillermo, Alessandro Anav, Pierre Friedlingstein, Stephen Sitch, Shilong Piao, Zaichun Zhu, Benjamin Poulter, Soenke Zaehle, Anders Ahlström, Mark Lomas, and et al. 2013. "Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs" Remote Sensing 5, no. 10: 4819-4838. https://doi.org/10.3390/rs5104819
APA StyleMurray-Tortarolo, G., Anav, A., Friedlingstein, P., Sitch, S., Piao, S., Zhu, Z., Poulter, B., Zaehle, S., Ahlström, A., Lomas, M., Levis, S., Viovy, N., & Zeng, N. (2013). Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs. Remote Sensing, 5(10), 4819-4838. https://doi.org/10.3390/rs5104819