The Fast Cooking and Enhanced Iron Bioavailability Properties of the Manteca Yellow Bean (Phaseolus vulgaris L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Yellow Bean Panel
2.2. Field Design and Storage Conditions
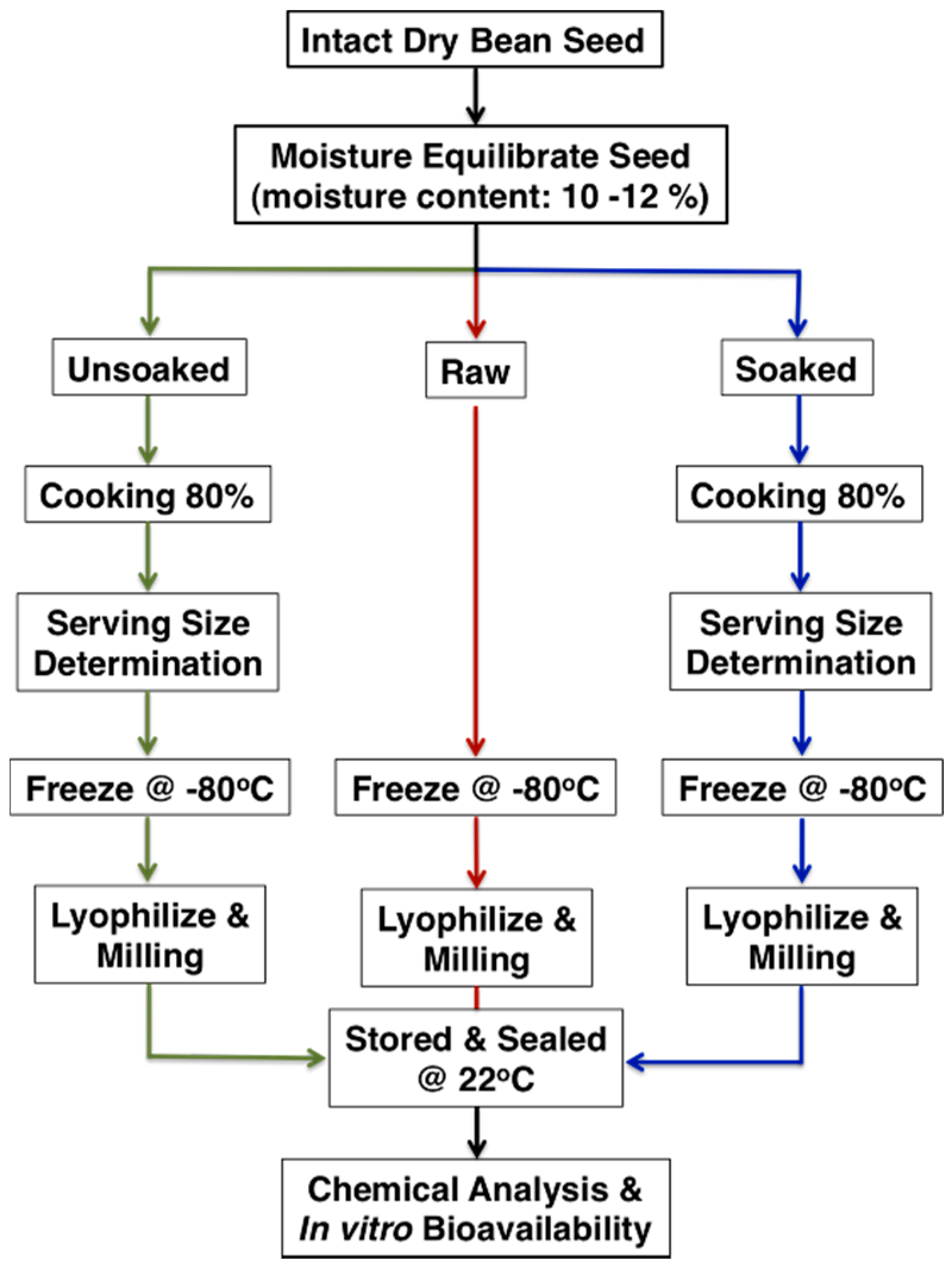
2.3. Moisture Equilibration, Cooking Time Determination and Sample Preparation
2.4. Iron Analysis
2.5. Iron Content, Serving-Size, Dietary Reference Intake and Retention Values
2.6. Iron Bioavailability: In Vitro Digestion/Caco-2 Cell Bioassay
2.7. Statistical Analysis
3. Results
3.1. Cooking Times and Cooking Classifications of the YBP
3.2. Iron Density of the YBP
3.3. Iron Retention Values of the YBP
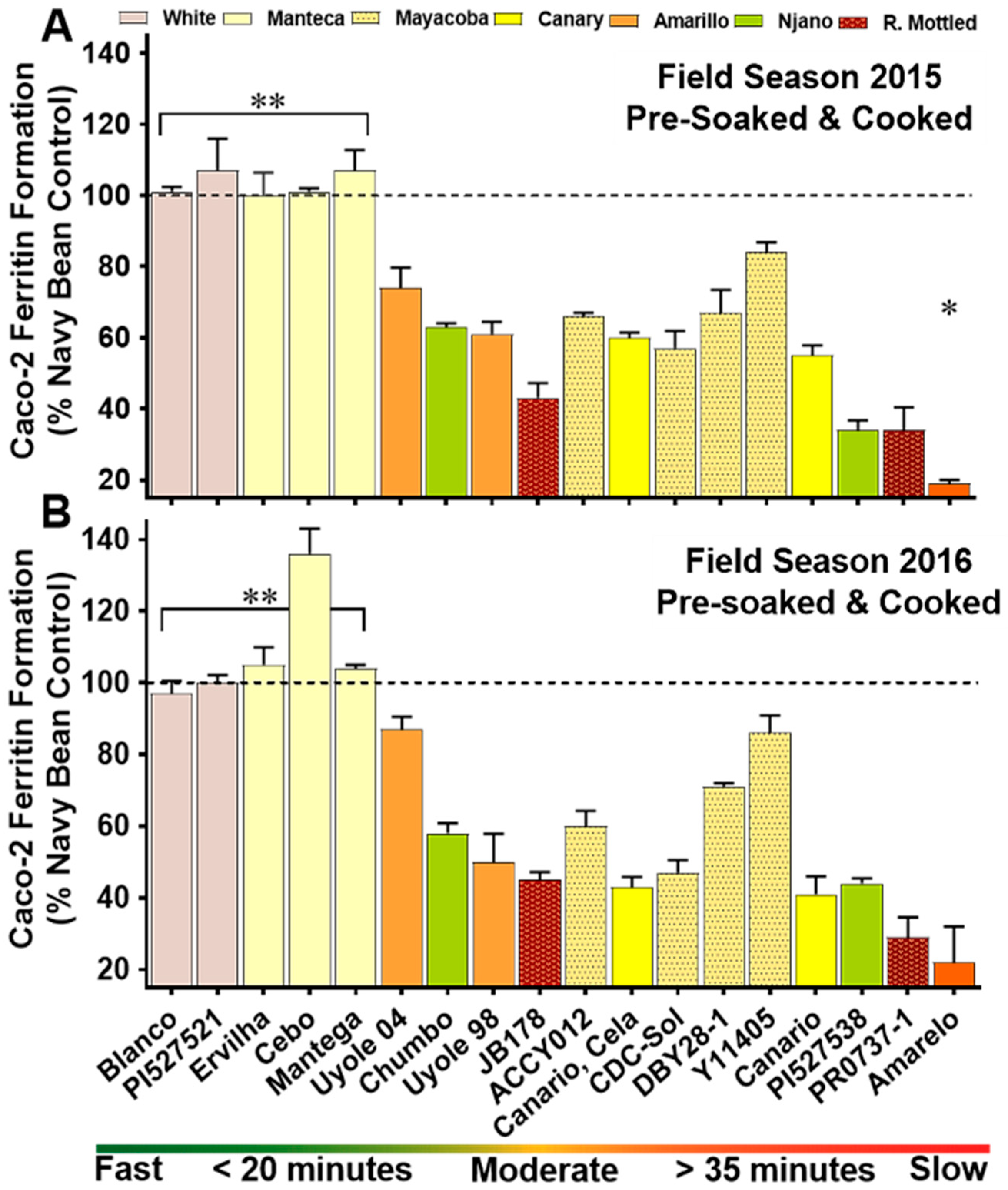
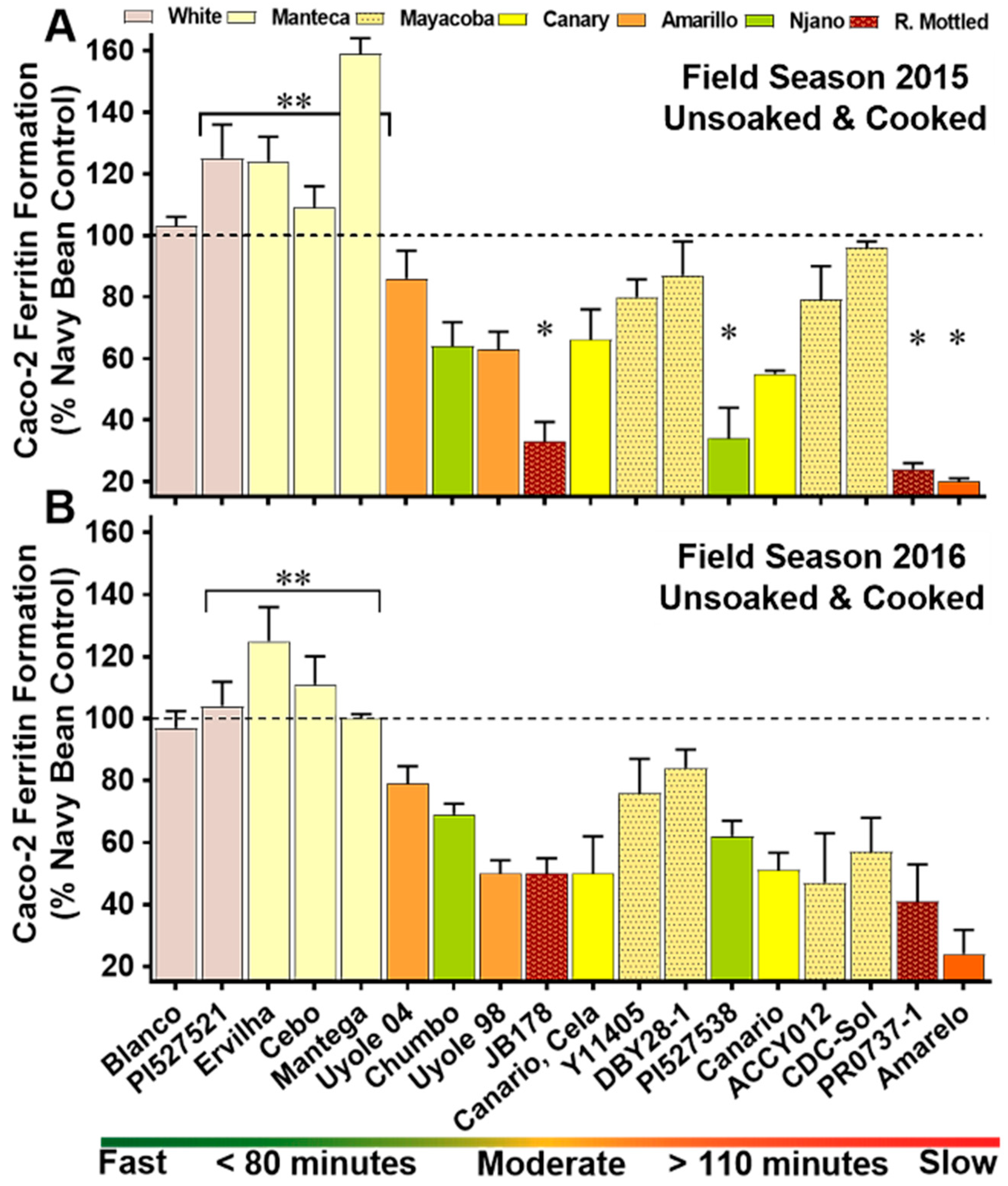
3.4. Iron Bioavailability of the YBP
4. Discussion
4.1. The YBP Is a Model to Explore the Health Benefits Yellow Beans
4.2. The Manteca Yellow Bean: A Genetic Resource for the New Generation of Fast Cooking Andean Beans
4.3. Iron Nutrition Benefits of the Fast Cooking Manteca Yellow Bean
4.4. Profile of the Manteca
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bouis, H.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Secur. 2017, 12, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Boy, E.; Wirth, J.; Hurrell, R. Review: The potential of the common bean (Phaseolus vulgaris) as a vehicle for iron biofortification. Nutrients 2015, 7, 1144–1173. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.; Tako, E.; Kochian, L.; Glahn, R. Indentification of black bean (Phaseolus vulgaris L.) polyphenols that inhibit and promote iron uptake by caco-2 cells. J. Agric. Food Chem. 2015, 63, 5950–5956. [Google Scholar] [CrossRef] [PubMed]
- Glahn, R.; Tako, E.; Cichy, K.; Wiesinger, J. The cotyledon cell wall and intracellular matrix are factors that limit iron bioavailability of the common bean (Phaseolus vulgaris). Food Funct. 2016, 7, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, J.; Guzmán, M.; Jiménez, A.; Mejia, C.; Ramos Muñoz, J.; Gallegos Acosta, J.; Hoyos, G. Preferential habits of consumers of common bean (Phaseolus vulgaris L.) in Mexico. Arch. Latinoam. Nutr. 1997, 47, 163–167. [Google Scholar] [PubMed]
- Palmer, S.; Winham, D.; Oberhauser, A.; Litchfield, R. Socio-ecological barriers to dry grain pulse consumption among low-income women: A mixed methods approach. Nutrients 2018, 10, 1108. [Google Scholar] [CrossRef] [PubMed]
- International Energy Agency. World Energy Outlook 2006; International Energy Agency: Paris, France, 2006. [Google Scholar]
- WEC. World Energy Insight: Powering up the South through Energy Poverty Alleviation; World Energy Council: London, UK, 2011. [Google Scholar]
- De Benoist, B.; Mclean, E.; Egli, I.; Cogswell, M. Worldwide Prevalence of Anaemia 1993–2005: WHO Global Data on Anaemia; WHO Press: Geneva, Switzerland, 2008. [Google Scholar]
- WHO Global Health Atlas. Map Library, World: Population Using Solid Fuels (%). 2013. Available online: http://apps.who.int/globalatlas/ (accessed on 24 June 2017).
- Adkins, E.; Oppelstrup, K.; Modi, V. Rural household energy consumption in the millennium villages in Sub-Saharan Africa. Energy Sustain Dev. 2012, 16, 249–259. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Shyamsundar, P.; Baccini, A. Forests biomass use and poverty in Malawi. Ecol. Econ. 2011, 70, 2461–2471. [Google Scholar] [CrossRef]
- FAO. Forests for Improved Nutrition and Food Security; Food and Agriculture Organization: Rome, Italy, 2011. [Google Scholar]
- Makungwa, S.; Epulani, F.; Woodhouse, I. Fuelwood supply: A missed essential component in a food security equation. J. Food Secur. 2013, 1, 49–51. [Google Scholar]
- Brouwer, I.; den Hartog, A.; Kamwendo, M.; Heldens, M. Wood quality and wood preferences in relation to food preparation and diet composition in Central Malawi. Ecol. Food Nutr. 1996, 35, 1–13. [Google Scholar] [CrossRef]
- Brouwer, I.; Hoorweg, J.; VanLiere, M. When households run out of fuel: Responses of rural households to decreasing fuelwood availability, Ntcheu District, Malawi. World Dev. 1997, 25, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Buruchara, R.; Chirwa, R.; Sperling, L.; Mukankusi, C.; Rubyogo, J.C.; Mutonhi, R.; Abang, M. Development and delivery of bean varieties in Africa: The Pan-Africa Bean Research Alliance (PABRA) model. Afr. Crop Sci. J. 2011, 19, 227–245. [Google Scholar]
- USAID Feed the Future. The U.S. Government’s Global Food Security Research Strategy. Available online: http://www.feedthefuture.gov (accessed on 8 April 2017).
- PABRA. Transforming Agriculture for Better Incomes and Diets in Africa; Pan-Africa Bean Research Alliance Press: Nairobi, Kenya, 2017; Available online: http://www.pabra-africa.org (accessed on 23 March 2018).
- Wiesinger, J.; Cichy, K.; Glahn, R.; Grusak, M.; Brick, M.; Thompson, H.; Tako, E. Demonstrating A Nutritional Advantage to the Fast Cooking Dry Bean (Phaseolus vulgaris L.). J. Agric. Food Chem. 2016, 64, 8592–8603. [Google Scholar] [CrossRef] [PubMed]
- Cichy, K.; Wiesinger, J.; Mendoza, F. Genetic diversity and genome wide association analysis of cooking time in dry bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2015, 128, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Voysest, O. Yellow Beans in Latin America; Report 0084-7747; Centro Internacional de Agricultura Tropical (CIAT): Cali, Colombia, 2012. [Google Scholar]
- Sichilima, T.; Mapemba, L.; Tembo, G. Drivers of dry common beans trade in Lusaka, Zambia: A trader’s perspective. Sustain. Agric. Res. 2016, 5, 15–25. [Google Scholar] [CrossRef]
- United Nations. A Value Chain Analysis of the Dry Bean Sub-Sector in Uganda : Development of Inclusive Markets in Agriculture and Trade (DIMAT) Project; United Nations Development Programme Uganda Issuing Body: Kampala, Uganda, 2012. [Google Scholar]
- Sones, D. Soya Njano is the bean for home consumption. In Our Blog: The Inside Story; Africa Soil Health Consortium: Nairobi, Kenya, 2015; Available online: http://africasoilhealth.cabi.org/2015/09/29/soya-njano-is-the-bean-for-home-consumption/ (accessed on 28 July 2017).
- Hillocks, R.; Madata, C.; Chirwa, R.; Minja, E.; Nchimbi-Msolla, S. Phaseolus bean improvement in Tanzania, 1959–2005. Euphytica 2006, 150, 215–231. [Google Scholar] [CrossRef]
- Canadian Food Insepection Agency (CFIA). CDC Sol. Available online: http://www.inspection.gc.ca/english/plaveg/pbrpov/cropreport/bea/app00007688e.shtml (accessed on 15 May 2018).
- Balasubramanian, P.; Chatterton, S.; Conner, R. AAC Y012 and AAC Y015 yellow dry bean. Can. J. Plant Sci. 2017, 97, 340–343. [Google Scholar] [CrossRef]
- Arnaud-Santana, E.; Nin, J.; Saladin, F.; Bodoy-Lutz, G.; Beaver, J.; Coyne, D.; Steadman, J. Registration of “JB-178” red mottled bean. Crop Sci. 2000, 40, 857–858. [Google Scholar]
- Prophete, E.; Demosthenes, G.; Godoy-Lutz, G.; Porch, T.; Beaver, J. Registration of PR0633-10 and PR0737-1 red mottled dry bean germplasm lines with resistance to BGYMV, BCMV, BCMNV and common bacterial blight. J. Plant Regist. 2014, 8, 49–52. [Google Scholar] [CrossRef]
- Hooper, S.; Wiesinger, J.; Echeverria, D.; Thompson, H.; Brick, M.; Nchimbi-Msolla, S.; Cichy, K. The carbohydrate profile of a dry bean (Phaseolus vulgaris L.) panel encompassing broad genetic variability for cooking time. Cereal Chem. 2017, 94, 135–141. [Google Scholar] [CrossRef]
- Morris, H.; Wood, E. Influence of moisture content on keeping quality of dry beans. Food Technol. 1956, 10, 225–229. [Google Scholar]
- Wang, N.; Daun, J. Determination of cooking times of pulses using an automated Mattson cooker apparatus. J. Sci. Food Agric. 2005, 85, 1631–1635. [Google Scholar] [CrossRef]
- Wang, N.; Hatcher, D.; Tyler, R.; Toews, R.; Gawalko, E. Effect of cooking on the composition of beans (Phaseolus vulgaris L.) and chickpeas (Cicer arietinum L.). Food Res. Int. 2010, 43, 589–594. [Google Scholar] [CrossRef]
- Marinangeli, C.; Curran, J.; Barr, S.; Slavin, J.; Puri, S.; Seaminathan, S.; Tapsell, L.; Patterson, C. Enhancing nutrition with pulses: Defining a recommended serving size for adults. Nutr. Rev. 2017, 75, 990–1006. [Google Scholar] [CrossRef] [PubMed]
- NAS. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; National Academy of Sciences National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Glahn, R.; Lee, O.; Yeung, A.; Goldman, M. Caco-2 Cell ferritin formation predicts nonradiolabeled food iron availability in an in vitro digestion/Caco-2 Cell culture model. J. Nutr. 1998, 128, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Glahn, R.; Tako, E.; Hart, J.; Haas, J.; Lung’aho, M.; Beebe, S. Iron bioavailability studies of the first generation of iron-biofortified beans released in Rwanda. Nutrients 2017, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Borchgrevink, C. Culinary perspective of dry beans and pulses. In Dry Beans and Pulses Production, Processing and Nutrition; Uebersax, M.S., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2013; pp. 313–334. [Google Scholar]
- Tako, E.; Glahn, R. White beans provide more bioavailable iron than red beans: Studies in poultry (Gallus gallus) and an in vitro digestion/Caco-2 model. Int. J. Vitam. Nutr. Res. 2010, 80, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Ariza-Nieto, M.; Blair, M.; Welch, R.; Glahn, R. Screening of iron bioavailability patterns in eight bean (Phaseolus vulgaris L.) genotypes using the Caco-2 cell in vitro model. J. Agric. Food Chem. 2007, 55, 7950–7956. [Google Scholar] [CrossRef] [PubMed]
- Beebe, S.; Viviana Gonzalez, A.; Rengifo, J. Research on trace minerals in the common bean. Food Nutr. Bull. 2000, 21, 387–391. [Google Scholar] [CrossRef]
- McClean, P.; Moghaddam, S.; Lopez-Millan, A.; Brick, M.; Kelly, J.; Miklas, P.; Osorno, J.; Porch, T.; Urrea, C.; Soltani, A.; et al. Phenotypic diversity for seed mineral concentration in North American dry bean germplasm of Middle American ancestry. Crop Sci. 2017, 57, 3129–3144. [Google Scholar] [CrossRef]
- Barampama, Z.; Simard, R. Oligosaccharides, antinutritional factors and protein disgestibility of dry beans as affected by processing. J. Food Sci. 1994, 4, 833–838. [Google Scholar] [CrossRef]
- Pujola, M.; Farreras, A.; Casanas, F. Protein and starch content of raw, soaked and cooked beans (Phaseolus vulgaris L.). Food Chem. 2007, 102, 1034–1041. [Google Scholar] [CrossRef]
- Saha, S.; Singh, G.; Mahajan, V.; Gupta, H. Variability of nutritional and cooking quality in bean (Phaseolus vulgaris L) as a function of genotype. Plant Foods Hum. Nutr. 2009, 64, 174–180. [Google Scholar] [CrossRef] [PubMed]
- McCouch, S.; Baute, J.G.; Bradeen, J.; Bramel, P.; Bretting, P.K.; Buckler, E.; Burke, M.J.; Charest, D.; Cloutier, S.; Cole, G.; et al. Feeding the future. Nature 2013, 499, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Kolkman, J.; Schneider, K. Breeding for yield in dry bean (Phaseolus vulgaris L.). Euphytica 1998, 102, 343–356. [Google Scholar] [CrossRef]
- Bassett, M.J. The Seedcoat Color of Genotypes Prim, the Manteca and Coscorrón Market Classes of Common Bean. HortScience. 1999, 34, 336–337. [Google Scholar]
- Leakey, C. Breeding on the C and J and B loci for modification of bean seedcoat flavonoids with the objective of improving food acceptability. Annu. Rep. Bean Improv. Coop. 1992, 35, 13–17. [Google Scholar]
- Leakey, C.; Hosfield, G.; Dubois, A. Mantecas, a new class of beans (Phaseolus vulgaris) of enhanced digestibility. In Proceedings of the 3rd European Conference on Grain Legumes, Valladolid, Spain, 14–19 November 1998; pp. 336–337. [Google Scholar]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. J. Mol. Evol. 1983, 19, 150–170. [Google Scholar] [CrossRef]
- Deshpande, S.; Cheryan, M. Water uptake during cooking of dry beans (Phaseolus vulgaris L.). Plant Foods Hum. Nutr. 1986, 36, 157–165. [Google Scholar] [CrossRef]
- Santos, G.; Ribeiro, N.; Maziero, S. Evaluation of common bean morphological traits identifies grain thickness directly correlated with cooking time. Pesqui. Agropecu. Trop. 2016, 46, 35–42. [Google Scholar] [CrossRef]
- Agbo, G.; Hosfield, G.; Uebersax, M.; Klomparens, K. Seed microstructure and its relationship to water uptake in isogenic lines and a cultivar of dry beans (Phaseolus vulgaris L.). Food Struct. 1987, 6, 12. [Google Scholar]
- Beninger, C.; Hosfield, G. Antioxidant activity of extracts, condensed tannin fractions, and pure flavonoids from Phaseolus vulgaris L. seed coat color genotypes. J. Agric. Food Chem. 2003, 51, 7879–7883. [Google Scholar] [CrossRef] [PubMed]
- Elia, F.; Hosfield, G.; Kelly, J.; Uebersax, M. Genetic analysis and interrelationships between traits for cooking time, water absorption, and protein and tannin content of Andean dry beans. J. Am. Soc. Hortic. Sci. 1997, 122, 512–518. [Google Scholar]
- Pereira, H.; Del Peloso, M.; Bassinello, P.; Guimaraes, C. Gentic variability for iron and zinc content in common bean lines and interaction with water availability. Genet. Mol. Res. J. 2014, 13, 6773–6785. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, N. Hybrid the History and Science of Plant Breeding; The University of Chicago Press: Chicago, IL, USA, 2009. [Google Scholar]
- Leakey, C. Progress in developing tannin-free dry phaseolus beans. Annu. Rep. Bean Improv. Coop. 2000, 43, 18–20. [Google Scholar]
- Hosfield, G.; Bennink, M.; Beninger, C.; Engleright, R.; Ospina, M. Variability for starch digestibility in dry bean (Phaseolus vulgaris L.). HortScience 1998, 33, 472. [Google Scholar]
- Engleright, R.; Beimiriki, M.; Hosfield, G. Determination of total dietary fiber, indigestible starch, and indigestible protein in dry bean (Phaseolus vulgaris L.). Annu. Rep. Bean Improv. Coop. 1999, 42, 123–124. [Google Scholar]
- Beninger, C.; Hosfield, G.; Nair, M. Flavonol glycosides from the seed coat of a new Manteca-type dry bean (Phaseolus v ulgaris L.). J. Agric. Food Chem. 1998, 46, 2906–2910. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Hart, J.; Tako, E.; Glahn, R. Characterization of polyphenol effects on inhibition and promotion of iron uptake by caco-2 cells. J. Agric. Food Chem. 2017, 65, 3285–3294. [Google Scholar] [CrossRef] [PubMed]
- Bassett, M.; Lee, R.; Otto, C.; McClean, P. Classical and molecular genetic studies of the strong greenish yellow seedcoat color in “Wagenaar” and “Enola” common bean. J. Am. Soc. Hortic. Sci. 2002, 127, 50–55. [Google Scholar]
- Beninger, C.; Hosfield, G.; Bassett, M. Flavonoid composition of three genotypes of dry bean (Phaseolus vulgaris) differing in seedcoat color. J. Am. Soc. Hortic. Sci. 1999, 124, 514–518. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Type | Genotype | Collection Site | Source | Cultivation | COD |
|---|---|---|---|---|---|
| Manteca | Ervilha | IIA | Huambo, Angola | Landrace * | Andean |
| Manteca | Cebo | marketplace | Cela, Angola | Landrace * | Andean |
| Manteca | Mantega Blanca | marketplace | Kibala, Angola | Landrace * | Andean |
| Mayocoba | CDC-Sol | Canada | Unv. of Saskatchewan | Variety | Andean |
| Mayocoba | ACC Y012 | Canada | Alberta | Variety | Andean |
| Mayocoba | Y11405 | United States | Michigan State Unv. | Breeding Line | Andean |
| Mayocoba | DBY28-1 | United States | Oregon State Unv. | Breeding Line | Andean |
| Canary | Canario | IIA | Huambo, Angola | Landrace * | Andean |
| Canary | Canario, Cela | marketplace | Cela, Angola | Landrace * | Andean |
| Amarillo (lt.) | Uyole 04 | Tanzania | Tanzania Breeding | Variety | Andean |
| Amarillo (dk.) | Uyole 98 | Tanzania | Tanzania Breeding | Variety | Andean |
| Amarillo (dk.) | Amarelo | marketplace | Cela, Angola | Landrace * | MA |
| Njano | Chumbo | marketplace | Cela, Angola | Landrace * | Andean |
| Njano | PI527538 | Burundi | US GRIN | Landrace | Andean |
| White | PI527521 | Burundi | US GRIN | Landrace | Andean |
| White | Blanco Fanesquero | Ecuador | INIAP | Variety | Andean |
| Red Mottled | JB178 | Dominican Rep. | CIAS | Variety | Andean |
| Red Mottled | PR0737-1 | Puerto Rico | Unv. of Puerto Rico | Variety | Andean |
| Genotype (Seed Type) | Cooking Time (min) 2 | Cooking Class |
|---|---|---|
| Blanco (white) | 16 k | fast |
| PI527521 (white) | 18 k | fast |
| Ervilha (Manteca) | 18 j,k | fast |
| Cebo (Manteca) | 19 j,k | fast |
| Mantega (Manteca) | 19 j,k | fast |
| Uyole 04 (lt. Amarillo) | 22 i,j | moderate |
| Chumbo (Njano) | 24 h,i | moderate |
| Uyole 98 (dk. Amarillo) | 26 f,g,h | moderate |
| JB178 (Red Mottled) | 26 g,h | moderate |
| ACC Y012 (Mayocoba) | 28 e,f,g | moderate |
| Canario, Cela (Canary) | 29 e,f,g | moderate |
| CDC-Sol (Mayocoba) | 30 d,e,f | moderate |
| DBY28-1 (Mayocoba) | 31 d,e | moderate |
| Y11405 (Mayocoba) | 33 d | moderate |
| Canario (Canary) | 38 c | slow |
| PI527538 (Njano) | 39 c | slow |
| PR0737-1 (Red Mottled) | 59 b | slow |
| Amarelo (dk. Amarillo) | 69 a | slow |
| Genotype (Seed Type) | Cooking Time (min) 2 | Cooking Class |
|---|---|---|
| Blanco (white) | 76 k,l | fast |
| PI527521 (white) | 76 j,k,l | fast |
| Ervilha (Manteca) | 76 l | fast |
| Cebo (Manteca) | 76 l | fast |
| Mantega (Manteca) | 79 i,j,k | fast |
| Uyole 04 (lt. Amarillo) | 82 h,i,j | moderate |
| Chumbo (Njano) | 83 h | moderate |
| Uyole 98 (dk. Amarillo) | 83 h,i | moderate |
| JB178 (Red Mottled) | 95 g | moderate |
| Canario, Cela (Canary) | 101 f | moderate |
| Y11405 (Mayocoba) | 101 f | moderate |
| DBY28-1 (Mayocoba) | 108 d,e | moderate |
| PI527538 (Njano) | 108 e | moderate |
| Canario (Canary) | 112 c,d | slow |
| ACC Y012 (Mayocoba) | 113 b,c | slow |
| CDC-Sol (Mayocoba) | 116 b | slow |
| PR0737-1 (Red Mottled) | 124 a | slow |
| Amarelo (dk. Amarillo) | 126 a | slow |
| One Serving Size (Half Cup) | |||||
|---|---|---|---|---|---|
| 2015 | 2016 | ||||
| Genotype (Seed Type) | Cooking Class | Iron (mg) 2 | % DRI 3 | Iron (mg) | % DRI |
| Blanco (white) | fast | 1.95 d,e,f | 11 | 2.28 b,c,d,e | 13 |
| PI527521 (white) | fast | 2.13 c,d | 12 | 2.32 b,c,d | 13 |
| Ervilha (Manteca) | fast | 2.02 d,e | 11 | 2.30 b,c,d,e | 13 |
| Cebo (Manteca) | fast | 1.75 f,g | 10 | 2.02 g,h | 11 |
| Mantega (Manteca) | fast | 2.06 c,d | 11 | 2.29 b,c,d,e | 13 |
| Uyole 04 (lt. Amarillo) | moderate | 1.84 e,f,g | 10 | 2.16 d,e,f,g | 12 |
| Chumbo (Njano) | moderate | 1.98 d,e | 11 | 2.25 c,d,e,f | 12 |
| Uyole 98 (dk. Amarillo) | moderate | 1.85 e,f,g | 10 | 2.06 f,h,g | 11 |
| JB178 (Red Mottled) | moderate | 2.71 a | 15 | 2.89 a | 16 |
| ACC Y012 (Mayocoba) | moderate | 1.84 e,f,g | 10 | 2.10 e,f,g | 12 |
| Canario, Cela (Canary) | moderate | 2.24 c | 12 | 2.30 b,c,d,e | 13 |
| CDC-Sol (Mayocoba) | moderate | 1.82 e,f,g | 10 | 1.95 g,h | 11 |
| DBY28-1 (Mayocoba) | moderate | 1.73 g | 10 | 2.02 g,h | 11 |
| Y11405 (Mayocoba) | moderate | 2.63 a,b | 15 | 2.49 b | 14 |
| Canario (Canary) | slow | 1.98 d,e | 11 | 2.14 d,e,f,g | 12 |
| PI527538 (Njano) | slow | 1.71 g | 10 | 1.87 h | 10 |
| PR0737-1 (Red Mottled) | slow | 2.49 b | 14 | 2.45 b,c | 14 |
| Amarelo (dk. Amarillo) | slow | 1.70 g | 9 | 2.02 g,h | 11 |
| One Serving Size (Half Cup) | |||||
|---|---|---|---|---|---|
| 2015 | 2016 | ||||
| Genotype (Seed Type) | Cooking Class | Iron (mg) 2 | % DRI 3 | Iron (mg) | % DRI |
| Blanco (white) | fast | 2.07 c,d | 11 | 2.24 b,c,d | 12 |
| PI527521 (white) | fast | 1.98 d,e | 11 | 2.17 b,c,d,e,f | 12 |
| Ervilha (Manteca) | fast | 2.19 b,c | 12 | 2.20 b,c,d,e | 12 |
| Cebo (Manteca) | fast | 1.62 i,j | 9 | 2.00 e,f,g,h | 11 |
| Mantega (Manteca) | fast | 1.85 e,f | 10 | 2.01 e,f,g,h | 11 |
| Uyole 04 (lt. Amarillo) | moderate | 1.68 f,g,h,i,j | 9 | 2.12 d,e,f,g | 12 |
| Chumbo (Njano) | moderate | 1.83 e,f,g | 10 | 1.95 g,h,i | 11 |
| Uyole 98 (dk. Amarillo) | moderate | 1.79 f,g,h | 10 | 1.98 f,g,h,i | 11 |
| JB178 (Red Mottled) | moderate | 2.43 a | 13 | 2.49 a | 14 |
| Canario, Cela (Canary) | moderate | 2.25 b | 12 | 2.14 d,e,f,g | 12 |
| Y11405 (Mayocoba) | moderate | 2.50 a | 14 | 2.35 a,b | 13 |
| DBY28-1 (Mayocoba) | moderate | 1.65 h,i,j | 9 | 1.90 h,i | 11 |
| PI527538 (Njano) | moderate | 1.60 j | 9 | 1.79 i | 10 |
| Canario (Canary) | slow | 2.01 d | 11 | 2.04 e,f,g,h | 11 |
| ACC Y012 (Mayocoba) | slow | 1.68 g,h,i,j | 9 | 1.89 h,i | 10 |
| CDC-Sol (Mayocoba) | slow | 1.77 f,g,h,i | 10 | 1.83 h,i | 10 |
| PR0737-1 (Red Mottled) | slow | 2.11 b,c,d | 12 | 2.28 a,b,c | 13 |
| Amarelo (dk. Amarillo) | slow | 1.56 j | 9 | 1.39 j | 8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiesinger, J.A.; Cichy, K.A.; Tako, E.; Glahn, R.P. The Fast Cooking and Enhanced Iron Bioavailability Properties of the Manteca Yellow Bean (Phaseolus vulgaris L.). Nutrients 2018, 10, 1609. https://doi.org/10.3390/nu10111609
Wiesinger JA, Cichy KA, Tako E, Glahn RP. The Fast Cooking and Enhanced Iron Bioavailability Properties of the Manteca Yellow Bean (Phaseolus vulgaris L.). Nutrients. 2018; 10(11):1609. https://doi.org/10.3390/nu10111609
Chicago/Turabian StyleWiesinger, Jason A., Karen A. Cichy, Elad Tako, and Raymond P. Glahn. 2018. "The Fast Cooking and Enhanced Iron Bioavailability Properties of the Manteca Yellow Bean (Phaseolus vulgaris L.)" Nutrients 10, no. 11: 1609. https://doi.org/10.3390/nu10111609
APA StyleWiesinger, J. A., Cichy, K. A., Tako, E., & Glahn, R. P. (2018). The Fast Cooking and Enhanced Iron Bioavailability Properties of the Manteca Yellow Bean (Phaseolus vulgaris L.). Nutrients, 10(11), 1609. https://doi.org/10.3390/nu10111609