Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Liver Samples, Total Liver TAG Measurement, and PNPLA3 Genotyping
2.2. Lipidyzer Platform
2.3. Data Evaluation
2.4. Statistics
3. Results
3.1. Characteristics of Study Groups
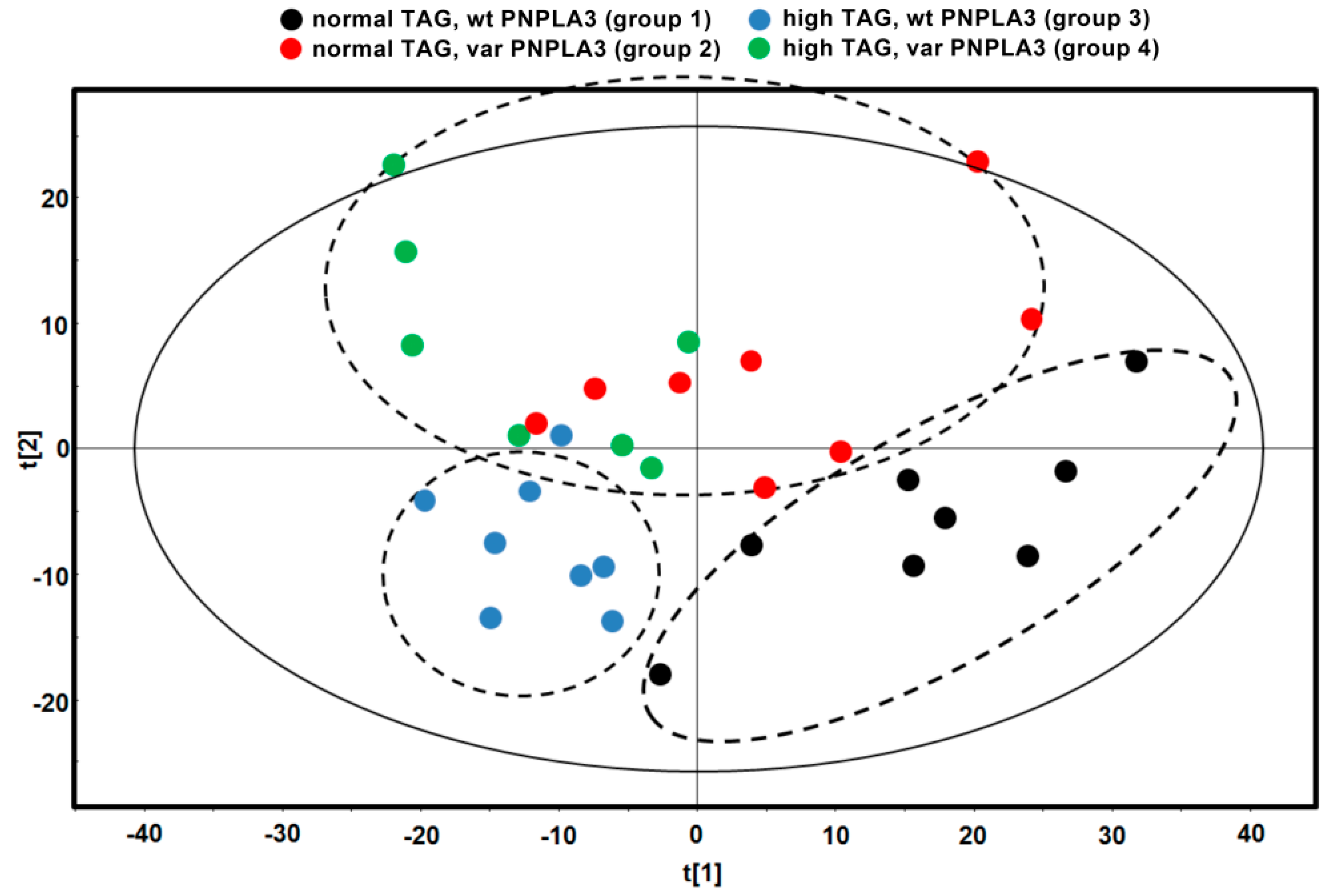
3.2. I148M PNPLA3 Variant Does Not Change Relative Total Lipid Contents
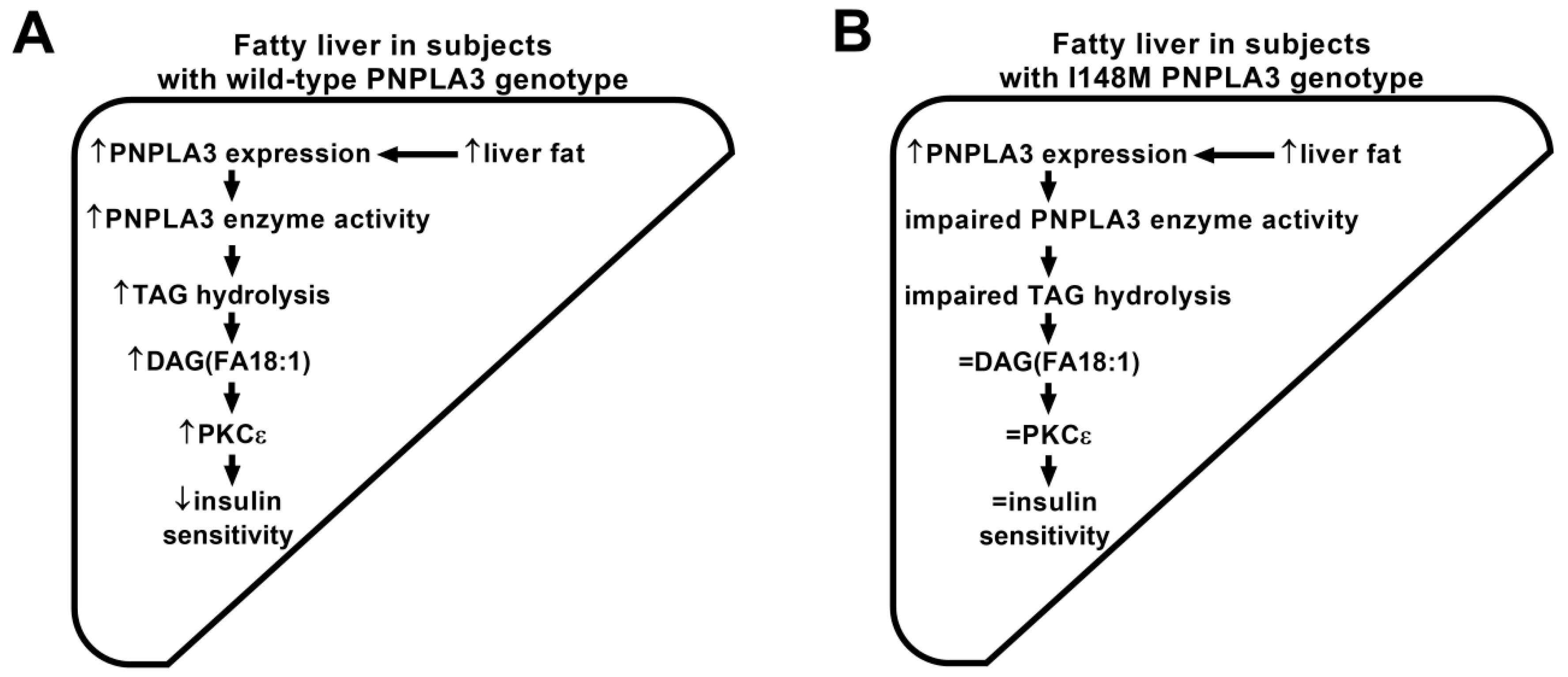
3.3. Increased Liver Fat Content Is Associated with High Proportion of DAG (FA18:1) Species in Subjects with Wild-Type PNPLA3, However, DAG (FA18:1) Remains Unchanged in I148M PNPLA3 Carriers
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DAG | Diacylglycerol |
| FA | Fatty acid |
| group 1 | Subjects with normal liver fat and wild-type PNPLA3 |
| group 2 | Subjects with normal liver fat and I148M/M148M PNPLA3 variants |
| group 3 | Subjects with high TAG and wild-type PNPLA3 |
| group 4 | Subjects with high TAG and I148M/M148M PNPLA3 variants |
| MAG | Monoacylglycerol |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| PKC | Protein kinase C |
| PNPLA3 | Patatin-like phospholipase domain-containing protein 3 |
| TAG | Triacylglycerol |
| var | I148M/M148M PNPLA3 variants |
| wt | Wild-type PNPLA3 allele |
References
- Stefan, N.; Schick, F.; Häring, H.U. Causes, characteristics, and consequences of metabolically unhealthy normal weight in humans. Cell Metab. 2017, 26, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Häring, H.U. The role of hepatokines in metabolism. Nat. Rev. Endocrinol. 2013, 9, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Loomba, R.; Rinella, M.E.; Bugianesi, E.; Marchesini, G.; Neuschwander-Tetri, B.A.; Serfaty, L.; Negro, F.; Caldwell, S.H.; Ratziu, V.; et al. Current and future therapeutic regimens for nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Hepatology 2018, 68, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Böhm, A.; Hoffmann, C.; Irmler, M.; Schneeweiss, P.; Schnauder, G.; Sailer, C.; Schmid, V.; Hudemann, J.; Machann, J.; Schick, F.; et al. TGF-beta contributes to impaired exercise response by suppression of mitochondrial key regulators in skeletal muscle. Diabetes 2016, 65, 2849–2861. [Google Scholar] [CrossRef] [PubMed]
- Franko, A.; Neschen, S.; Rozman, J.; Rathkolb, B.; Aichler, M.; Feuchtinger, A.; Brachthäuser, L.; Neff, F.; Kovarova, M.; Wolf, E.; et al. Bezafibrate ameliorates diabetes via reduced steatosis and improved hepatic insulin sensitivity in diabetic TallyHo mice. Mol. Metab. 2017, 6, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Itoh, M.; Suganami, T.; Kanai, S.; Shirakawa, I.; Sakai, T.; Asakawa, M.; Yoneyama, T.; Kai, T.; Ogawa, Y. Obeticholic acid protects against hepatocyte death and liver fibrosis in a murine model of nonalcoholic steatohepatitis. Sci. Rep. 2018, 8, 8157. [Google Scholar] [CrossRef] [PubMed]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2008, 40, 1461–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baclig, M.O.; Lozano-Kuhne, J.P.; Mapua, C.A.; Gopez-Cervantes, J.; Natividad, F.F. Genetic variation I148M in patatin-like phospholipase 3 gene and risk of non-alcoholic fatty liver disease among Filipinos. Int. J. Clin. Exp. Med. 2014, 7, 2129–2136. [Google Scholar] [PubMed]
- Kantartzis, K.; Peter, A.; Machicao, F.; Machann, J.; Wagner, S.; Konigsrainer, I.; Konigsrainer, A.; Schick, F.; Fritsche, A.; Häring, H.U.; et al. Dissociation between fatty liver and insulin resistance in humans carrying a variant of the patatin-like phospholipase 3 gene. Diabetes 2009, 58, 2616–2623. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, I.A.; Ericson, U.; Rukh, G.; Riddestrale, M.; Romeo, S.; Orho-Melander, M. The PNPLA3 Ile148Met interacts with overweight and dietary intakes on fasting triglyceride levels. Genes Nutr. 2014, 9, 388. [Google Scholar] [CrossRef] [PubMed]
- Sevastianova, K.; Santos, A.; Kotronen, A.; Hakkarainen, A.; Makkonen, J.; Silander, K.; Peltonen, M.; Romeo, S.; Lundbom, J.; Lundbom, N.; et al. Effect of short-term carbohydrate overfeeding and long-term weight loss on liver fat in overweight humans. Am. J. Clin. Nutr. 2012, 96, 727–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scorletti, E.; West, A.L.; Bhatia, L.; Hoile, S.P.; McCormick, K.G.; Burdge, G.C.; Lillycrop, K.A.; Clough, G.F.; Calder, P.C.; Byrne, C.D. Treating liver fat and serum triglyceride levels in NAFLD, effects of PNPLA3 and TM6SF2 genotypes: Results from the WELCOME trial. J. Hepatol. 2015, 63, 1476–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, A.; Kovarova, M.; Nadalin, S.; Cermak, T.; Konigsrainer, A.; Machicao, F.; Stefan, N.; Häring, H.U.; Schleicher, E. PNPLA3 variant I148M is associated with altered hepatic lipid composition in humans. Diabetologia 2014, 57, 2103–2107. [Google Scholar] [CrossRef] [PubMed]
- Luukkonen, P.K.; Zhou, Y.; Sadevirta, S.; Leivonen, M.; Arola, J.; Oresic, M.; Hyotylainen, T.; Yki-Jarvinen, H. Hepatic ceramides dissociate steatosis and insulin resistance in patients with non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Cohen, J.C.; Hobbs, H.H. Expression and characterization of a PNPLA3 protein isoform (I148M) associated with nonalcoholic fatty liver disease. J. Biol. Chem. 2011, 286, 37085–37093. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Schoiswohl, G.; Chitraju, C.; Paar, M.; Cornaciu, I.; Rangrez, A.Y.; Wongsiriroj, N.; Nagy, H.M.; Ivanova, P.T.; Scott, S.A.; et al. Adiponutrin functions as a nutritionally regulated lysophosphatidic acid acyltransferase. Cell Metab. 2012, 15, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Mancina, R.M.; Matikainen, N.; Maglio, C.; Soderlund, S.; Lundbom, N.; Hakkarainen, A.; Rametta, R.; Mozzi, E.; Fargion, S.; Valenti, L.; et al. Paradoxical dissociation between hepatic fat content and de novo lipogenesis due to PNPLA3 sequence variant. J. Clin. Endocrinol. Metab. 2015, 100, 821–825. [Google Scholar]
- Petaja, E.M.; Yki-Jarvinen, H. Definitions of normal liver fat and the association of insulin sensitivity with acquired and genetic NAFLD-A systematic review. Int. J. Mol. Sci. 2016, 17, 633. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Huang, Y.; Karaman, R.; Ivanova, P.T.; Brown, H.A.; Roddy, T.; Castro-Perez, J.; Cohen, J.C.; Hobbs, H.H. Chronic overexpression of PNPLA3I148M in mouse liver causes hepatic steatosis. J. Clin. Investig. 2012, 122, 4130–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, V.T.; Shulman, G.I. Nonalcoholic fatty liver disease as a nexus of metabolic and hepatic diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef] [PubMed]
- Jelenik, T.; Kaul, K.; Sequaris, G.; Flogel, U.; Phielix, E.; Kotzka, J.; Knebel, B.; Fahlbusch, P.; Horbelt, T.; Lehr, S.; et al. Mechanisms of insulin resistance in primary and secondary nonalcoholic fatty liver. Diabetes 2017, 66, 2241–2253. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, K.W.; Gilijamse, P.W.; Versteeg, R.I.; Ackermans, M.T.; Nederveen, A.J.; la Fleur, S.E.; Romijn, J.A.; Nieuwdorp, M.; Zhang, D.; Samuel, V.T.; et al. Hepatic diacylglycerol-associated protein kinase cepsilon translocation links hepatic steatosis to hepatic insulin resistance in humans. Cell Rep. 2017, 19, 1997–2004. [Google Scholar] [CrossRef] [PubMed]
- Ollero, M. Methods for the study of lipid metabolites in cystic fibrosis. J. Cyst. Fibros. 2004, 3, 97–98. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Franken, H.; Dammeier, S.; Rosenbaum, L.; Kantartzis, K.; Peter, A.; Zell, A.; Adam, P.; Li, J.; Xu, G.; et al. Circulating lysophosphatidylcholines are markers of a metabolically benign nonalcoholic fatty liver. Diabetes Care 2013, 36, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Franko, A.; Huypens, P.; Neschen, S.; Irmler, M.; Rozman, J.; Rathkolb, B.; Neff, F.; Prehn, C.; Dubois, G.; Baumann, M.; et al. Bezafibrate improves insulin sensitivity and metabolic flexibility in STZ-induced diabetic mice. Diabetes 2016, 65, 2540–2552. [Google Scholar] [CrossRef] [PubMed]
- Markgraf, D.F.; Al-Hasani, H.; Lehr, S. Lipidomics-reshaping the analysis and perception of type 2 diabetes. Int. J. Mol. Sci. 2016, 17, 1841. [Google Scholar] [CrossRef] [PubMed]
- Ubhi, B.K. Direct Infusion-Tandem Mass Spectrometry (DI-MS/MS) analysis of complex lipids in human plasma and serum using the lipidyzer platform. Methods Mol. Biol. 2018, 1730, 227–236. [Google Scholar] [PubMed]
- Peter, A.; Kantartzis, K.; Machicao, F.; Machann, J.; Wagner, S.; Templin, S.; Königsrainer, I.; Königsrainer, A.; Schick, F.; Fritsche, A.; et al. Visceral obesity modulates the impact of apolipoprotein C3 gene variants on liver fat content. Int. J. Obes. 2012, 36, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Franko, A.; Kovarova, M.; Feil, S.; Feil, R.; Wagner, R.; Heni, M.; Köngisrainer, A.; Ruoß, M.; Nüssler, A.K.; Weigert, C.; et al. cGMP-dependent protein kinase I (cGKI) modulates human hepatic stellate cell activation. Metabolism 2018, 88, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hoene, M.; Li, J.; Li, Y.; Zhao, X.; Häring, H.U.; Schleicher, E.D.; Weigert, C.; Xu, G.; Lehmann, R. Simultaneous extraction of metabolome and lipidome with methyl tert-butyl ether from a single small tissue sample for ultra-high performance liquid chromatography/mass spectrometry. J. Chromatogr. A 2013, 1298, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumashiro, N.; Yoshimura, T.; Cantley, J.L.; Majumdar, S.K.; Guebre-Egziabher, F.; Kursawe, R.; Vatner, D.F.; Fat, I.; Kahn, M.; Erion, D.M.; et al. Role of patatin-like phospholipase domain-containing 3 on lipid-induced hepatic steatosis and insulin resistance in rats. Hepatology 2013, 57, 1763–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amati, F. Revisiting the diacylglycerol-induced insulin resistance hypothesis. Obes. Rev. 2012, 13, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Moro, C.; Galgani, J.E.; Luu, L.; Pasarica, M.; Mairal, A.; Bajpeyi, S.; Schmitz, G.; Langin, D.; Liebisch, G.; Smith, S.R. Influence of gender, obesity, and muscle lipase activity on intramyocellular lipids in sedentary individuals. J. Clin. Endocrinol. Metab. 2009, 94, 3440–3447. [Google Scholar] [CrossRef] [PubMed]
- Szendroedi, J.; Yoshimura, T.; Phielix, E.; Koliaki, C.; Marcucci, M.; Zhang, D.; Jelenik, T.; Muller, J.; Herder, C.; Nowotny, P.; et al. Role of diacylglycerol activation of PKCtheta in lipid-induced muscle insulin resistance in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 9597–9602. [Google Scholar] [CrossRef] [PubMed]
- Franko, A.; von Kleist-Retzow, J.C.; Neschen, S.; Wu, M.; Schommers, P.; Böse, M.; Kunze, A.; Hartmann, U.; Sanchez-Lasheras, C.; Stoehr, O.; et al. Liver adapts mitochondrial function to insulin resistant and diabetic states in mice. J. Hepatol. 2014, 60, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Ayala, J.E.; Bracy, D.P.; McGuinness, O.P.; Wasserman, D.H. Considerations in the design of hyperinsulinemic-euglycemic clamps in the conscious mouse. Diabetes 2006, 55, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Dziewulska, A.; Dobrzyn, P.; Jazurek, M.; Pyrkowska, A.; Ntambi, J.M.; Dobrzyn, A. Monounsaturated fatty acids are required for membrane translocation of protein kinase C-theta induced by lipid overload in skeletal muscle. Mol. Membr. Biol. 2012, 29, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6, a009191. [Google Scholar] [CrossRef] [PubMed]
- Hyysalo, J.; Gopalacharyulu, P.; Bian, H.; Hyotylainen, T.; Leivonen, M.; Jaser, N.; Juuti, A.; Honka, M.J.; Nuutila, P.; Olkkonen, V.M.; et al. Circulating triacylglycerol signatures in nonalcoholic fatty liver disease associated with the I148M variant in PNPLA3 and with obesity. Diabetes 2014, 63, 312–322. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Group 1 Normal TAG, wt PNPLA3 | Group 2 Normal TAG, var PNPLA3 | Group 3 High TAG, wt PNPLA3 | Group 4 High TAG, var PNPLA3 |
|---|---|---|---|---|
| Age (years) | 59.3 ± 12.6 | 60.6 ± 16.4 | 64.0 ± 11.8 | 65.1 ± 14.5 |
| Body weight (kg) | 79.3 ± 9.9 | 80.3 ± 13.5 | 86.2 ± 11.1 | 87.1 ± 13.1 |
| BMI (kg/m2) | 26.1 ± 3.2 | 28.0 ± 5.3 | 31.0 ± 3.4 | 28.6 ± 4.0 |
| ALT (U/L) | 24.5 ± 6.5 | 22.0 ± 5.2 | 31.1 ± 9.3 | 35.7 ± 18.6 |
| Liver fat (%) | 1.1 ± 0.8 | 1.5 ± 0.7 | 5.9 ± 2.0 *** | 7.6 ± 2.9 *** |
| PNPLA3 148 (II/IM/MM) (n) | 8/0/0 | 0/7/1 | 8/0/0 | 0/6/1 |
| Sex (m/f) (n) | 6/2 | 4/4 | 5/3 | 5/2 |
| Number of subjects (n) | 8 | 8 | 8 | 7 |
| Lipid Class % | Group 1 Normal TAG, wt PNPLA3 | Group 2 Normal TAG, var PNPLA3 | Group 3 High TAG, wt PNPLA3 | Group 4 High TAG, var PNPLA3 |
|---|---|---|---|---|
| TAG | 25.63 ± 12.49 | 35.22 ± 15.30 | 68.64 ± 11.26 *** | 69.82 ± 11.86 *** |
| DAG | 0.60 ± 0.30 | 0.57 ± 0.17 | 0.92 ± 0.12 * | 1.02 ± 0.20 ** |
| FFA | 7.23 ± 3.57 | 6.07 ± 1.71 | 2.60 ± 1.12 *** | 2.37 ± 1.01 ** |
| CER | 0.26 ± 0.04 | 0.25 ± 0.08 | 0.10 ± 0.03 *** | 0.10 ± 0.05 *** |
| DCER | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.01 ± 0.00 *** | 0.01 ± 0.01 *** |
| HCER | 0.07 ± 0.02 | 0.05 ± 0.02 | 0.03 ± 0.01 *** | 0.02 ± 0.01 ** |
| LCER | 0.06 ± 0.01 | 0.06 ± 0.03 | 0.04 ± 0.02 | 0.03 ± 0.01 |
| PC | 38.51 ± 9.74 | 33.06 ± 8.63 | 14.86 ± 6.41 *** | 14.75 ± 7.83 *** |
| LPC | 0.66 ± 0.22 | 0.60 ± 0.14 | 0.24 ± 0.14 *** | 0.28 ± 0.17 ** |
| PE | 20.43 ± 5.80 | 17.84 ± 5.82 | 8.19 ± 3.24 *** | 7.29 ± 3.10 *** |
| LPE | 0.16 ± 0.05 | 0.15 ± 0.03 | 0.06 ± 0.03 *** | 0.07 ± 0.04 *** |
| CE | 2.45 ± 0.34 | 2.84 ± 0.45 | 2.73 ± 0.60 | 2.86 ± 0.48 |
| SM | 3.91 ± 0.81 | 3.28 ± 0.81 | 1.59 ± 0.67 *** | 1.37 ± 0.60 *** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franko, A.; Merkel, D.; Kovarova, M.; Hoene, M.; Jaghutriz, B.A.; Heni, M.; Königsrainer, A.; Papan, C.; Lehr, S.; Häring, H.-U.; et al. Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation. Nutrients 2018, 10, 1314. https://doi.org/10.3390/nu10091314
Franko A, Merkel D, Kovarova M, Hoene M, Jaghutriz BA, Heni M, Königsrainer A, Papan C, Lehr S, Häring H-U, et al. Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation. Nutrients. 2018; 10(9):1314. https://doi.org/10.3390/nu10091314
Chicago/Turabian StyleFranko, Andras, Dietrich Merkel, Marketa Kovarova, Miriam Hoene, Benjamin A. Jaghutriz, Martin Heni, Alfred Königsrainer, Cyrus Papan, Stefan Lehr, Hans-Ulrich Häring, and et al. 2018. "Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation" Nutrients 10, no. 9: 1314. https://doi.org/10.3390/nu10091314
APA StyleFranko, A., Merkel, D., Kovarova, M., Hoene, M., Jaghutriz, B. A., Heni, M., Königsrainer, A., Papan, C., Lehr, S., Häring, H. -U., & Peter, A. (2018). Dissociation of Fatty Liver and Insulin Resistance in I148M PNPLA3 Carriers: Differences in Diacylglycerol (DAG) FA18:1 Lipid Species as a Possible Explanation. Nutrients, 10(9), 1314. https://doi.org/10.3390/nu10091314